牛磺酸消炎抗氧化机理及其在幼龄动物中的应用
2015-05-09王庆争杜孝贵曹伟平唐志如张小龙
徐 玲 王庆争 杜孝贵 曹伟平 唐志如* 张小龙
(1.西南大学动物科技学院,生物饲料与分子营养实验室,重庆400715;2.湖南省永兴县樟树中学,郴州423311)
牛磺酸消炎抗氧化机理及其在幼龄动物中的应用
徐 玲1王庆争1杜孝贵1曹伟平2唐志如1*张小龙1
(1.西南大学动物科技学院,生物饲料与分子营养实验室,重庆400715;2.湖南省永兴县樟树中学,郴州423311)
牛磺酸广泛存在于动物组织细胞内,具有消炎抗氧化生物学功能。目前研究发现牛磺酸消炎抗氧化机理包括:通过雷帕霉素靶蛋白(mTOR)信号通路促进肠黏膜修复,通过抑制Toll样受体4(TLR4)信号通路缓解肠黏膜炎症反应,通过瞬时受体电位通道M2(TRPM2)离子通道改善细胞线粒体生理功能。本文就牛磺酸消炎抗氧化机理以及在幼龄动物中的应用进行综述,以期为牛磺酸在消炎抗氧化方面研究提供参考意义。
牛磺酸;消炎;抗氧化;幼龄动物
牛磺酸又称2-氨基乙磺酸,分布于中枢神经系统和肌肉组织。包括内源性牛磺酸和外源性牛磺酸,其中内源合成牛磺酸主要在肝脏合成,经由血液循环将其运送到靶组织即可满足大多数动物组织需要[1],但幼龄动物由于自身合成能力有限,必须从外界摄取一定量的牛磺酸来满足自身营养需求。牛磺酸具有广泛的生物学功能,可以作为多种组织渗透剂,也是维持机体内环境稳态的重要因素;此外在幼龄动物消炎抗氧化、增强细胞膜抗氧化能力、削弱减少组织辐射与氧化损伤、调节激素释放和提高机体免疫力等方面也具有重要作用。
1 牛磺酸的合成与代谢
仔猪体内牛磺酸是以蛋氨酸、胱氨酸和半胱氨酸为原料合成而来,其主要合成部位为心脏、肝脏和脑部。在合成过程中,首先由蛋氨酸或胱氨酸去甲基为同型半胱氨酸,同型半胱氨酸与丝氨酸作用合成胱硫醚,胱硫醚在胱硫醚酶作用下失去α-酮酸和氨基生成半胱氨酸。半胱氨酸可经过4条途径合成牛磺酸,其中哺乳动物体内主要合成途径为:半胱氨酸转变为半胱亚磺酸后,在半半胱亚磺酸脱羧酶(cysteinesulfinate decar-boxylase, CSD)作用下脱去二氧化碳(CO2)转化为亚牛磺酸,亚牛磺酸被氧化酶氧化为牛磺酸;半胱亚磺酸也可变为磺基丙氨酸,然后在CSD作用下脱去CO2转化为牛磺酸。因体内牛磺酸的合成受到限速酶CSD的调控,而新生动物CSD活性和合成能力又较低,内源合成不能满足机体所需,所以需要进行外源补充牛磺酸[2]。
肾脏是牛磺酸排泄的主要器官,大多数的牛磺酸未发生反应就排出体外。在饲粮牛磺酸摄入量较低时,机体通过减少肾脏牛磺酸的排泄量来保证其在体内含量[3]。另外,牛磺酸在体内的代谢途径主要有:1)合成牛磺胆酸,促进脂肪和脂类物质的消化吸收;2)生成脒基牛磺酸,产物在低等动物的能量代谢中可作为一种磷酸源;3)分解生成异乙基硫氨酸,具有调节离子生物膜的作用;4)生成氨基甲酸牛磺酸,功能尚待研究[4]。
2 牛磺酸消炎抗氧化机理
2.1 牛磺酸抑制肠黏膜炎症反应与Toll样受体4(TLR4)信号通路
断奶应激通过神经、内分泌和免疫途径引起肠黏膜屏障功能的下降,主要包括促肾上腺皮质激素释放激素(CRH)的释放,迷走神经的激活,肥大细胞分泌介质引发的免疫细胞、肠道神经细胞和上皮细胞之间的相互作用,引起肠黏膜对小分子(包括细菌来源的化学趋化肽)和大分子(尤其是具有抗原潜能的蛋白质)通透性增强,进而引起抗原通过松弛的细胞连接进入固有层与免疫细胞发生相互作用,加剧应激诱导的炎症反应,最终造成肠黏膜屏障功能的破坏[5]。
Roy等[6]报道,牛磺酸可通过DNA的转录因子切断一氧化氮合成限速酶(NOS)和肿瘤坏死因子α(TNF-α)表达,降低一氧化氮(NO)和TNF-α水平,进而抑制应激诱导的炎症反应,最终保护肺细胞(图1)。Kontny等[7]研究表明,牛磺酸-Cl抑制类风湿性关节炎成纤维细胞样的滑膜细胞炎症细胞因子的产生,其原因在于牛磺酸-Cl抑制了激活蛋白1(AP-1)核转录因子-κB(nuclearfactor-κB,NF-κB)的激活。Barua等[8]研究也发现,牛磺酸能抑制肠黏膜NF-κB的表达及活化,抑制包细胞介素(IL)-1β的生成,减少TNF-α的分泌,阻断炎症的关键步骤及其次级炎症,促进肠黏膜修复及功能恢复。静息时,NF-κB与其抑制蛋白(inhibitory κB,IκB)结合,以无活性形式存在于胞浆中;应激、细菌脂多糖、抗原和自由基等能使IκB快速磷酸化从而激活NF-κB,进入细胞核内与靶基因结合,参与免疫、应激和炎症反应相关基因表达的调控[9],介导炎症反应对肠黏膜的损害[10],造成绒毛萎缩和隐窝增生,引起肠道屏障损伤[11],并且NF-κB活化水平与肠道病理损伤呈显著正相关[12]。
脂多糖(LPS)诱导的信号转导通路中,TLR4是控制着LPS炎症信号进入细胞内启动炎症反应的“门户”蛋白质,是LPS信号转导上游的关键分子,完成介导脂多糖的信号转导通路[13]。TLR4的信号转导主要有2种类型:髓样分化因子88(MyD88)依赖型信号通路和MyD88非依赖型信号通路,MyD88依赖型信号通路的主要接头分子是MyD88和Ma1,MyD88非依赖型信号通路的主要接头分子是β干扰素TIR结构域衔接蛋白(TRIF)和TRIF相关接头分子(TRAM)[14]。在LPS/TLR4信号传导下游通路中,NF-κB的激活能够促进众多与炎症反应相关的基因产物的表达,是LPS诱导细胞产生炎性反应的关键中间环节。在炎症状态下,TLR4表达上调,并激活细胞内信号转导,通过NF-κB促进细胞因子和炎症介质的释放。抑制NF-κB的活化,能明显降低肠坏死范围,改善肠黏膜结构功能[15]。牛磺酸抑制NF-κB的活化是否与信号通路TLR4有关有待进一步研究(图2)。
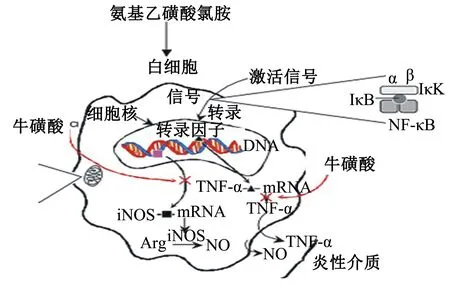
iNOS:诱导型一氧化氮合成酶 inosinducible nitric oxide synthase; Arg:精氨酸 arginine。
图1 牛磺酸抑制致炎介质TNF-α和NO产生的示意图
Fig.1 The inhibition of taurine on inflammatory factor TNF-α and NO[6]
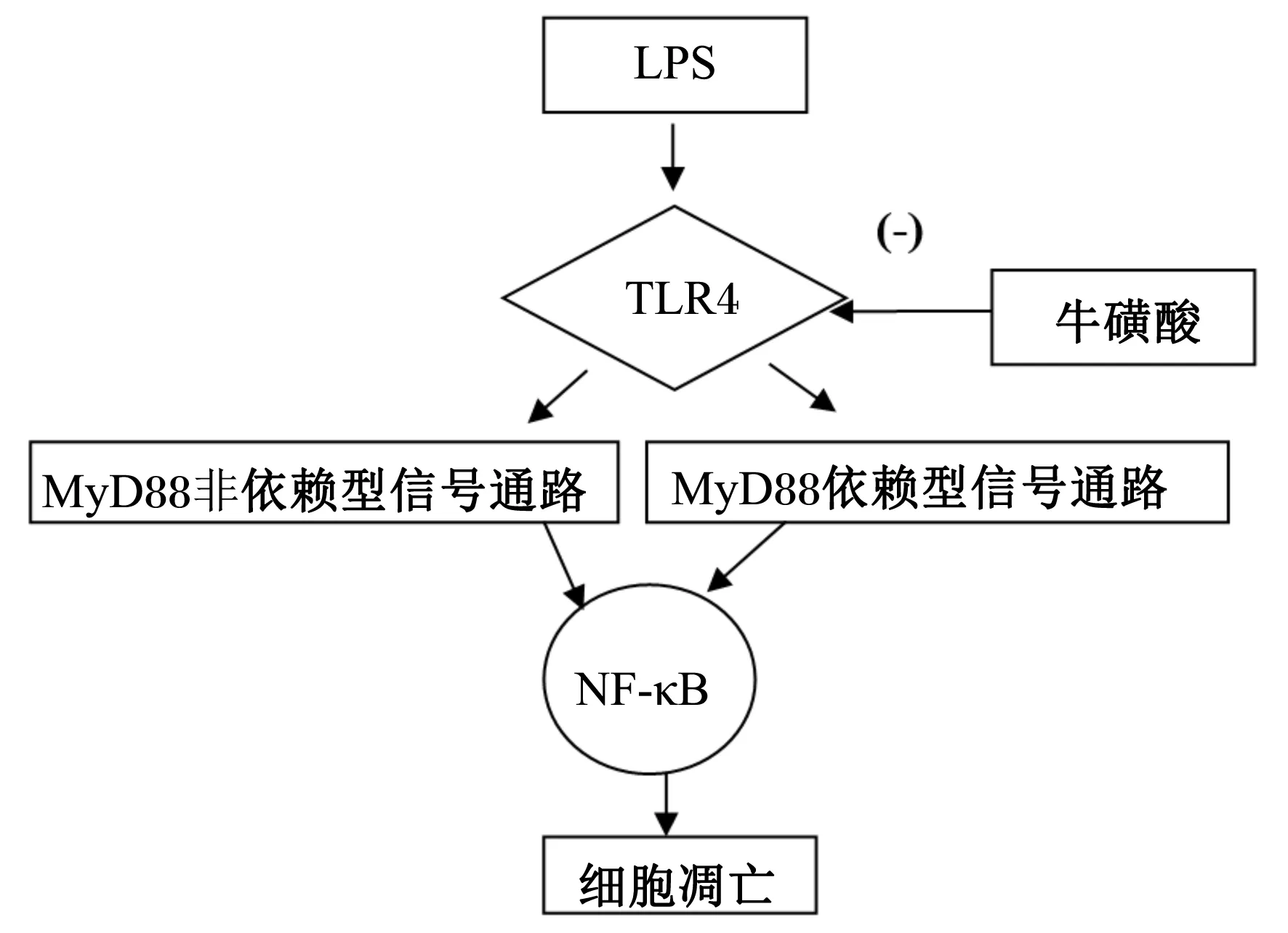
图2 牛磺酸抑制NF-κB活化的路径
2.2 牛磺酸改善线粒体生理功能与瞬时受体电位通道M2(TRPM2)
线粒体是存在于真核生物细胞质中含有核外遗传物质的细器,被称为细胞的“动力站”,其主要作用是进行氧化磷酸化合成三磷酸腺苷(ATP),为机体提供能量,是细胞代谢的主要控制者和细胞死亡程序的执行者[16]。牛磺酸多数生物学功能与线粒体相关。牛磺酸对受损的线粒体膜有保护作用可能与其抑制线粒体丙二醛(MDA)的生成、促进线粒体谷胱甘肽过氧化物酶(GSH-Px)活力的恢复、显著提高体线粒体还原型谷胱甘肽含量、显著提高线粒体超氧化物歧化酶(SOD)的活性、保持线粒体还原型谷胱甘肽含量及游离钙浓度和降低自由基对线粒体的攻击有关[17]。
牛磺酸调控线粒体对钙离子(Ca2+)的摄入与释放。牛磺酸抑制离体大鼠肝脏线粒体对Ca2+的摄入,并且抑制作用随牛磺酸浓度增加而增强,牛磺酸同时促进线粒体对Ca2+的释放,促进作用也呈剂量-效应关系[18]。牛磺酸抑制线粒体Ca2+摄入、促进线粒体Ca2+释放以调节线粒体钙稳态的作用可能是其对细胞进行保护的重要机制之一[2]。TRPM2是瞬时受体电位通道超家族成员之一,TRPM2的激活依赖细胞内Ca2+水平。氧化应激通过产生氧代谢产物如过氧化氢(H2O2),增加血管内皮渗透压,H2O2能促进二磷酸腺苷(ADP)-核糖体的形成,从而开启TRPM2通道(图3)[19]。因此推测牛磺酸可能通过抑制线粒体Ca2+摄入,进而抑制TRPM2的激活,最终达到抑制细胞膜脂质过氧化和保护细胞的目的。
3 牛磺酸在幼龄动物营养中的应用
3.1 家禽
补充牛磺酸可减少猝死综合征(SDS)和肉鸡总死亡率的肉鸡[3],对不同生长发育阶段动物发挥不同调节作用。曾得寿等[20]在肉仔鸡上通过对比发现,试验前期和后期各牛磺酸添加组的平均日增重均高于对照组,料重比低于对照组,且添加0.10%时效果显著。黄春喜等[21]研究发现,肉仔鸡饲粮添加牛磺酸显著降低了肉仔鸡早期(1~7日龄)料重比,显著提高胸腺指数,却对生长性能没有显著影响。但也有研究指出,肉仔鸡饲粮添加牛磺酸可不同程度地提高肉仔鸡生产性能,其中以添加0.10%牛磺酸效果最佳,且前期添加效果优于后期[22]。总体而言,饲粮添加牛磺酸的确可提高肉仔鸡的生产性能。
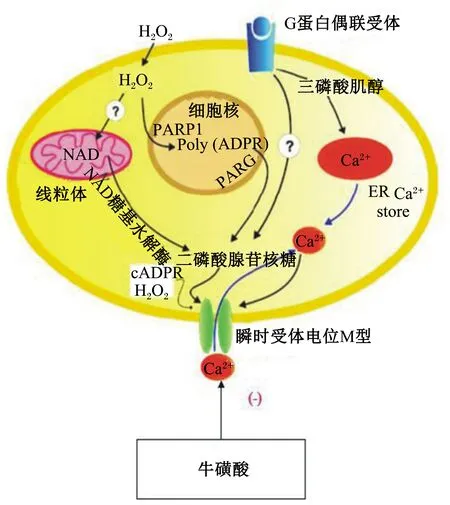
图3 牛磺酸调节线粒体TRPM2离子通道开启的路径
在消化吸收方面,家禽胰腺脂肪酶活性在出生时很低,伴随着成长而提升,脂肪酶活性的不足是限制脂肪消化和利用的重要因素[23]。Zeng等[24]试验表明,添加0.15%牛磺酸能显著高胰腺和小肠脂肪酶活性。牛磺酸与胆汁酸共轭能促进脂肪乳化提高脂肪酶的活动,然后增加了中性脂肪、总胆固醇(TC)、脂溶性维生素和其他脂溶性物质的消化率,从而提高饲料转化率促进生产[25]。Shim等[26]研究牛磺酸对家禽蛋白质合成和脂质代谢影响中发现牛磺酸显著降低肝脏TC和甘油三酯的浓度,刺激57、40和23 ku蛋白质在肉鸡肝脏的体外合成,但抑制54、37和24 ku蛋白质体外合成。总体表明牛磺酸在治疗降低胆固醇和甘油三酯的浓度时,也会影响肝脏中蛋白质合成。
饲粮添加牛磺酸倾向于增加6周肉鸡空肠的绒毛高度和隐窝深度,显著提高养分吸收和生产性能[26]。Satsu等[27]报道指出,老鼠饲粮补充牛磺酸可增加肌肉、肝脏、小肠、肾脏和其他组织牛磺酸含量,因此随着牛磺酸丰富型鸡肉逐渐作为营养食品和功能性食品,Huang等[28]研究发现,肉用鸡饲粮持续补充6周牛磺酸,增加肝、乳房和大腿肉牛磺酸含量且存在剂量依赖方式。其中添加8 g/kg牛磺酸与对照组相比增加了大腿和胸部肌肉牛磺酸含量的1.89和4.03倍。
牛磺酸及其衍生物能通过清除机体活性氧及其他氧化性有害物或通过增强机体抗氧化能力来保护动物组织,使其免受氧化基与自由基的损伤。李丽娟等[29]通过饲粮中添加牛磺酸研究爱拔益加肉鸡抗氧化酶活性发现饲粮中添加0.10%和0.15%的牛磺酸,血清和肝脏中谷GSH-Px、SOD活性及总抗氧化能力(T-AOC)最高,MDA浓度最低,而MDA是经自由基引发脂质过氧化的最终分解产物,其浓度高低可以反映体内脂质过氧化的程度。田莉莉等[30]研究表明,在肉仔鸡饲粮添加牛磺酸可以提高其抗氧化功能且3周龄肉仔鸡影响较显著。
在幼龄鸡上,相关研究报道指出牛磺酸可显著提高3周龄肉鸡胸下指数、脾脏指数和6周龄法氏囊指数[31]。而这3个指数是衡量机体免疫器官发育程度的重要指标。Wang等[32]饲粮补充0.05%牛磺酸增大了日本鹌鹑法氏囊和胸腺的相对重量。同样对于笼养蛋雏鸭饲粮添加0.10%牛磺酸显著提高了蛋雏鸭的平均日增重、SOD活性、GSH-Px活性、T-AOC以及法氏囊和脾脏指数[33]。在分子水平上,牛磺酸影响鸡端粒反转录酶(chicken telomerase reverse transcriptase,chTERT)转录水平及马立克氏病脾脏T淋巴细胞系(MDCC-MS)细胞端粒酶活性的下调,可能抑制肿瘤细胞的增殖,其中1%、2%效果最显著[34]。
3.2 仔猪
早期断奶仔猪饲粮中分别或同时添加0.25%甘氨酰谷氨酰胺和0.10%牛磺酸能有效地提高其肠道吸收功能,显著提高粗蛋白质(CP)、干物质(DM)、有机物(OM)消化率[35]。但是Liu等[36]研究发现,添加1.5%或3.0%牛磺酸对断奶猪生长性能、肠道健康均产生不利影响,其中炎性细胞因子(IL-6和TNF-α)和半胱天冬酶3(caspase-3)释放增强,caspase-3基因表达上调和胰高血糖素样肽2(GLP-2)分泌减弱,这些过程可能导致对断奶仔猪肠道功能有毒影响。因此,添加时应注意过量牛磺酸引起的毒性作用。在仔猪热应激方面,牛磺酸还可起到增加猪的采食量、降低体内皮质醇浓度的功能。
4 小 结
目前关于牛磺酸各方面的研究大多都是建立在以小鼠为试验动物的基础上,切实运用在畜禽(尤其是猪)上的临床研究较少。牛磺酸在幼龄动物上的营养作用还有很多问题有待解决,不仅要从牛磺酸的合成工艺上着手,降低成本改善工艺,缩短与欧美国家的差距;还应对畜禽进行定量分阶段深入研究牛磺酸生物学功能,量化不同种类动物不同生长发育阶段最适量,为饲料生产提供准确依据。总之,牛磺酸作为幼龄动物重要营养成分不论研究还是工业生产都具有广阔发展前景。
[1] QIAN X J,VINNAKOTA S,EDWARDS C,et al.Molecular characterization of taurine transport in bovine aortic endothelial cells[J].Biochimica et Biophysica Acta:Biomembranes,2000,1509(1/2):324-334.
[2] SCHULLER-LEVIS G B,PARK E.Taurine:new implications for an old amino acid[J].FEMS Microbiology Letters,2003,226(2):195-202.
[3] BLAIR R,JACOB J P,GARDINER E E.Lack of an effect of taurine supplementation on the incidence of sudden death syndrome in male broiler chicks[J].Poultry Science,1991,70(3):554-560.
[4] 徐晓峰,张力莉.牛磺酸在动物营养中的应用研究进展[J].饲料与畜牧,2007(4):31-33.
[5] SMITH F,CLARK J E,OVERMAN B L,et al.Early weaning stress impairs development of mucosal barrier function in the porcine intestine[J].American Journal of Physiology-Gastrointestinal and Liver Physiology,2010,298(3):G352-G363.
[6] ROY A,MANNA P,SIL P C.Prophylactic role of taurine on arsenic mediated oxidative renal dysfunction via MAPKs/NF-κB and mitochondria dependent pathways[J].Free Radical Research,2009,43(10):995-1007.
[7] KONTNY E,SZCZEPANSKA K,KOWALCZEWSKI J,et al.The mechanism of taurine chloramine inhibition of cytokine (interleukin-6,interleukin-8) production by rheumatoid arthritis fibroblast-like synoviocytes[J].Arthritis & Rheumatism,2000,43(10):2169-2177.
[8] BARUA M,LIU Y,QUINN M R.Taurine chloramine inhibits inducible nitric oxide synthase and TNF-α gene expression in activated alveolar macrophages:decreased NF-κB activation and IκB kinase activity[J].The Journal of Immunology,2002,167(4):2275-2281.
[9] ROGLER G,BRAND K,VOGL D,et al.Nuclear factor κB is activated in macrophages and epithelial cells of inflamed intestinal mucosa[J].Gastroenterology,1998,115(2):357-369.
[10] GITTER A H,BENDFELDT K,SCHULZKE J D,et al.Leaks in the epithelial barrier caused by spontaneous and TNF-α-induced single-cell apoptosis[J].The Journal of the Federation of American Societies for Experimental Biology,2000,14(12):1749-1753.
[11] SCHREIBER S,NIKOLAUS S,HAMPE J.Activation of nuclear factor κB in inflammatory bowel disease[J].Gut,1998,42(4):477-484.
[12] THIELE K,BIERHAUS A,AUTSCHBACH F,et al.Cell specific effects of glucocorticoid treatment on the NF-κBp65/IκBα system in patients with Crohn’s disease[J].Gut,1999,45(5):693-704.
[13] XU Y,JAGANNATH C,LIU X D,et al.Toll-like receptor 4 is a sensor for autophagy associated with innate immunity[J].Immunity,2007,27(1):135-144.
[14] AKIRA S.Mammalian Toll-like receptors[J].Current Opinion in Immunology,2003,15(1):5-11.
[15] JOBIN C,HOLT L,BRADHAM C A,et al.TNF receptor-associated factor-2 is involved in both IL-1β and TNF-α signaling cascades leading to NF-κB activation and IL-8 expression in human intestinal epithelial cells[J].The Journal of Immunology,1999,162(8):4447-4454.
[16] HENZE K,MARTIN W.Evolutionary biology:essence of mitochondria[J].Nature,2003,426(6963):127-128.
[17] 汪朝晖,张贤康,缪明永,等.牛磺酸在大鼠缺血再灌注过程中对肝线粒体的保护作用[J].中西医结合肝病杂志,2002,12(6):351-354,357.
[18] 马思聪,李磊,李海燕,等.瞬时受体电位2通道介导动物氧化应激的机理及其营养调控[J]动物营养学报,2013,25(10):2231-2237.
[19] HECQUET C M,AHMMED G U,VOGEL S M,et al.Role of TRPM2 channel in mediating H2O2-induced Ca2+entry and endothelial hyperpermeability[J].Circulation Research,2008,102(3):347-355.
[20] 曾得寿,高振华,赵京辉,等.牛磺酸对肉仔鸡生产性能、免疫器官发育及抗氧化能力的影响[J].中国兽医学报,2009,29(6):774-778.
[21] 黄春喜,袁建敏,周淑亮,等.牛磺酸对肉仔鸡生长性能、消化器官和免疫器官发育的影响[J].动物营养学报,2011,23(5):854-861.
[22] 刘玉芝,刘艳琴.牛磺酸对肉仔鸡生产性能和免疫功能的影响[J].扬州大学学报:农业与生命科学版,2008,29(4):45-48.
[23] NIR I,NITSAN Z,MAHAGNA M.Comparative growth and development of the digestive organs and of some enzymes in broiler and egg type chicks after hatching[J].British Poultry Science,1998,34(3):523-532.
[24] ZENG D S,GAO Z H,HUANG X L,et al.Effect of taurine on lipid metabolism of broilers[J].Journal of Applied Animal Research,2012,40(2):86-89.
[25] YUAN J M,WANG Z H.Effect of taurine on intestinal morphology and utilisation of soy oil in chickens[J].British Poultry Science,2010,51(4):540-545.
[26] SHIM K S,JUNG H J,NA C S,et al.Effects of taurine on lipid metabolism and protein synthesis in poultry and mice[J].Asian-Australasian Journal of Animal Sciences,2009,22(6):865-870.
[27] SATSU H,KOBAYASHI Y,YOKOYAMA T,et al.Effect of dietary sulfur amino acids on the taurine content of rat tissues[J].Amino Acids,2002,23(4):447-452.
[28] HUANG C X,WANG B,MIN Z,et al.Dietary inclusion level and time effects of taurine on broiler performance,meat quality,oxidative status and muscle taurine content[J].British Poultry Science,2014,55(5):598-604.
[29] 李丽娟,王安,王鹏.牛磺酸对爱拔益加肉雏鸡生长性能及抗氧化功能的影响[J].动物营养学报,2010,22(3):696-701.
[30] 田莉莉,李林.牛磺酸对肉仔鸡抗氧化能力的影响[J].饲料研究,2012(2):75-76.
[31] 李万军.牛磺酸对肉鸡饲粮养分利用率、免疫器官发育及屠宰性能的影响研究[J].中国农学通报,2012,28(23):6-10.
[32] WANG F R,DONG X F,TONG J M,et al.Effects of dietary taurine supplementation on growth performance and immune status in growing Japanese quail (Coturnixcoturnixjaponica)[J].Poultry Science,2009,88(7):1394-1398.
[33] 杨小然,王安,郭志杰.牛磺酸对笼养蛋雏鸭生长性能、抗氧化功能及免疫器官发育的影响[J].动物营养学报,2011,23(5):807-812.
[34] 于洋洋,陈涛,邱淑敏,等.牛磺酸对MDCC-MSB1细胞端粒酶表达水平及其活性的影响[J].饲料工业,2014,35(5):17-19.
[35] 张建斌,车向荣,陈娟娟.甘氨酰谷氨酰胺和牛磺酸对断奶仔猪肠道吸收功能的影响[J].中国畜牧兽医,2010,37(8):20-22.
[36] LIU Y,MAO X B,YU B,et al.Excessive dietary taurine supplementation reduces growth performance,liver and intestinal health of weaned pigs[J].Livestock Science,2014,168:109-119.
*Corresponding author, professor, E-mail: tangzhiru2326@sina.com.cn
(责任编辑 武海龙)
Anti-Inflammatory and Antioxidant Mechanism of Taurine and Its Application in Young Animals
XU Ling1WANG Qingzheng1DU Xiaogui1CAO Weiping2TANG Zhiru1*ZHANG Xiaolong1
(1.KeyLaboratoryforBio-FeedandAnimalNutrition,CollegeofAnimalScienceandTechnology,SouthwestUniversity,Chongqing400715,China; 2.YongxingCountyZhangxuMiddleSchool,Chenzhou423311,China)
Taurine is widely found in the cells of animal tissues and has anti-inflammatory and antioxidant biological function. Recent studies have found that anti-inflammatory and anti- oxidative mechanism of taurine is as follows: taurine promotes the repair of intestinal mucosa through mammalian target of rapamyein (mTOR) signaling pathway; taurine alleviates the inflammatory reaction of intestinal mucosa by inhibiting Toll-like-receptors (TLR4) signaling pathway; taurine improves mitochondrial function in cell physiology through the transient receptor potential melastatin type 2 (TRPM2) ion channels. In order to provide reference significance for taurine in anti-inflammatory and anti-oxidative research, the anti-inflammatory and anti-oxidative mechanism of taurine and the application of taurine in the young animals were reviewed in this paper.[ChineseJournalofAnimalNutrition, 2015, 27(12):3683-3688]
taurine; anti-inflammatory; antioxidant; young animals
10.3969/j.issn.1006-267x.2015.12.006
2015-06-08
雅琪创新基金项目(YQ 201401);国家973重点基础研究发展计划项目(2013CB127303);农业部948项目(2015Z74);中央高校基本业务费专项资金(XDJK2014B028,XDJK2015D005)
徐 玲(1994—),女,四川德阳人,本科生,研究兴趣为动物营养与饲料科学。E-mail: 1154885613@qq.com
*通信作者:唐志如,教授,硕士生导师,E-mail: tangzhiru2326@sina.com.cn
S811.2
A
1006-267X(2015)12-3683-06
