细菌信号小分子耐药机制的相关研究进展
2015-05-04易胜杰谢靖邱少富宋宏彬
易胜杰,谢靖,邱少富,宋宏彬
1.军事医学科学院 疾病预防控制所,北京 100071;2.中南大学 湘雅医学院基础医学院,湖南 长沙 410013
使用抗生素对抗感染性疾病是医学和人类健康历史上巨大的进步。自从青霉素被发现以来,研究人员基于对细菌关键蛋白的化学抑制研发了多种抗生素,目前已经在抗菌史上取得了十分伟大的成就。然而,这些年多重耐药菌的发生率不断提高,世界卫生组织更是发表声明称,细菌广泛的耐药已经明显成为威胁人类生命与健康的全球性问题[1]。人类的抗菌过程就是细菌与抗生素相互抵抗的过程,由于抗生素滥用等原因导致细菌的耐药性不断提高,但是近几十年来却只有2种新的抗生素被成功运用于临床治疗,这种不平衡将带来严重的安全隐患[2]。新型抗生素的匮乏很大程度上是因为我们关注的药物靶标比较单一,以及对细菌耐药机制的理解还不够深入[3]。
目前被广泛研究的细菌耐药机制主要包括:抗生素灭活酶、改变抗生素靶位结构、降低外膜通透性、加强主动外排系统、形成生物膜等[4-8]。但是最近几年,一种有别于传统理念的耐药机制被发现,并被证明广泛存在于细菌中,涌现了许多优秀的相关研究和评论[9-14]。这些研究聚焦于一些细菌通过代谢产生的信号小分子,这些信号小分子能够轻松地跨膜传递,它们通过诱导抗氧化酶表达等机制使细菌获得广谱的耐药性,甚至极少部分的细菌产生的信号小分子就能够导致整个菌群耐药。这一理论的提出,不仅使得原有的细菌耐药机制得到很好的补充和拓展,也为新型抗生素的研制提供了潜在性的靶标和思路。为了更好地使研究者了解生物小分子导致细菌耐药的机制,探索抗生素新的潜在靶标,我们通过细菌代谢产生的3种信号小分子——一氧化氮(NO)、硫化氢(H2S)和吲哚,探讨信号分子使细菌获得耐药性的相关机制。
1 NO使细菌获得广谱的耐药性
真核生物中的巨噬细胞或其他一些免疫细胞能通过诱导性一氧化氮合酶(inducible nitric oxide synthases,iNOS)催化产生NO,这些NO能够通过硝化应激和氧化应激作用控制胞内细菌的感染[15]。但是,“聪明”的细菌通过进化也能产生一种细菌一氧化氮合酶(bacterial nitric oxide synthases,bNOS)。bNOS没有还原酶区域,生物信息学分析显示许多细菌中都能找到和bNOS同源的蛋白,它们在进化上有密切联系,并且对细菌有着重要的生命学意义[16]。Gusarov等研究发现,细菌内源性的NO能够让细菌获得广谱的耐药性。研究发现,吖啶黄和NO混合处理过后对枯草芽孢杆菌和金黄色葡萄球菌的杀伤能力显著降低,构建的bNOS大肠杆菌表达体系也能显著降低吖啶黄的浓度,其主要原因是NO能直接与吖啶黄反应,使吖啶黄亚硝基化失活。不仅如此,NO还能保护细菌抵御吖啶黄引发的氧化应激损伤。这表明NO能够直接改造某些抗生素,并且抵御抗生素引发的氧化应激损伤,从而帮助细菌获得耐药性。那么,NO如何抵御抗生素引发的氧化应激损伤?研究人员发现,在绿脓菌素的压力下(NO不能直接与其发生反应),bNOS能够诱导过氧化物歧化酶(cata⁃lase and superoxide dismutase,SOD)的表达,从而抑制胞内氧化应激,并表现出对绿脓菌素的耐药性,但是bNOS缺失株却不能诱导SOD表达,也没有表现出耐药性。不仅如此,加入一定浓度的头孢呋辛后,野生株与bNOS缺失株在生长曲线和生存曲线上都表现出明显差异,而加入活性氧分子(reactive oxy⁃gen species,ROS)抑制剂或外源的NO都能显著提高缺失株的存活率。重复验证实验显示,NO可使细菌对我们常用的许多抗生素耐受[12]。而另一个重要机制在Shatalin等的研究中阐明,炭疽杆菌能通过bNOS产生的NO激活细菌的过氧化氢酶,并抑制对自身有损伤性的芬顿反应,最终达到抵御胞内氧化应激[17]。Gusarov等在枯草芽孢杆菌中也发现了类似机制,细菌依靠bNOS产生的NO来抵御宿主免疫系统带来的氧化应激损伤[18]。bNOS能够抵御来自宿主的氧化应激损伤,这在很大程度上也和抵抗抗生素相似,因为氧化应激很可能是许多抗生素杀菌过程中的关键因素[19]。耐甲氧西林金黄色葡萄球菌去除内源性NO后,就马上变得对氧化应激和万古霉素等抗生素敏感[20]。这些研究都表明,NO能通过多种机制帮助细菌获得广谱的耐药性。
2 H2S是细菌的一种普遍耐药机制
产H2S的机制普遍存在于真核和原核生物中。尽管研究结果有一些差异,但毋庸置疑的是H2S调节多种细胞功能,包括炎症、上皮细胞分泌、伤害感受等[21-23]。在哺乳动物中已发现3种可催化产生H2S的酶,即胱硫醚β-合成酶(cystathionine β-synthase,CBS)、胱硫醚γ-裂解酶(cystathionine γ-lyase,CSE)和3-巯基丙酮酸硫基转移酶(3-mercaptopyruvate sulfurtransferase,3MST)。对细菌的基因组分析发现,绝大部分细菌都至少能表达3种酶中的一种[13]。除此之外,一些特殊的细菌能基于不同的底物和途径产生H2S。如沙门菌就能通过phs和asr操纵子分别以硫代硫酸盐和亚硫酸盐为底物产生H2S[24-25]。在人体内,尤其是肠道环境中,宿主和细菌都能产生大量H2S,并且相互影响、相互作用[26-27]。对于细菌本身,H2S也有着重要的生物学意义。Shatalin等通过对4种不同细菌的研究发现,产H2S基因的缺失株与野生株相比对庆大霉素、卡那霉素、氯霉素更加敏感,在高浓度抗生素压力下缺失株存活率显著降低,但加入外源H2S后,存活率与野生株没有显著差异,这与bNOS对细菌的影响十分相似。对大肠杆菌的进一步研究发现,庆大霉素能够显著抑制3MST缺失株的生长,但是加入ROS抑制剂后生长曲线与野生株没有差异。另外,3MST缺失、过表达和野生株能不同程度地诱导过氧化氢酶和SOD的表达,3MST的活性与过氧化氢酶和SOD表达量是正相关的。这表明H2S能通过激活过氧化氢酶和SOD抵御抗生素引起的氧化应激损伤,从而使细菌获得耐药性。这一机制能使细菌获得长久的耐药性,但是在抗生素发挥作用的初期,H2S主要是通过与游离的Fe2+直接发生反应,从而抑制DNA损伤性的芬顿反应。多次验证实验结果还显示,H2S同样能使细菌对头孢类、喹诺酮类等常见抗生素的耐药性增加[13]。
3 少数细菌产生的吲哚就能导致菌群耐药
吲哚是一种细胞间的信号分子,多种革兰阴性和阳性细菌都能产生[28]。产生吲哚最重要的一种酶是色氨酸酶(tryptophanase,TnaA),它能可逆地将色氨酸转化为吲哚、丙酮酸和氨[29-30]。许多研究认为吲哚在大肠杆菌中是一种压力信号分子,在抗生素等压力状态下,吲哚能通过激活药物外排泵相关基因从而对抗生素耐受[28,31-32]。Lee等基于此进行了进一步的研究,他们不断提高大肠杆菌培养基中诺氟沙星的浓度,发现整个菌群都能耐药,但绝大多数大肠杆菌个体的耐药性都没有整体的耐药性高。通过全基因组测序分析,他们发现某些大肠杆菌在关键耐药基因上发生了突变,如gyrB22,而这些突变让这些大肠杆菌获得对诺氟沙星的耐药性。这些耐药菌通过产生吲哚可以保护其他对诺氟沙星敏感的菌株,但那些处于抗生素压力下的非耐药菌却不能产生吲哚。为了进一步探究吲哚导致大肠杆菌耐药的机制,研究者通过对野生株和TnaA缺失株在抗生素压力下的转录组对比分析,发现吲哚能够上调一些药物外排泵相关基因,如mdtE。更有意思的是,分析结果显示一个小RNA oxyS的表达量被下调了,这个小RNA是细菌内氧化应激的感受器。这些结果表明,吲哚至少可能通过激活药物外排基因以及降低氧化应激损伤使大肠杆菌对抗生素耐受[14]。吲哚诱导持留菌的形成或许也是导致细菌耐药的一种机制。持留菌的显著特点是,在抗生素等逆境条件下处于非分裂增长的休眠状态,降低代谢活动以维持自身的生存;而当抗生素或逆境条件消失时,持留菌又可重新恢复生长,并进行增殖再生。Vega等发现,在氧氟沙星的压力下,大肠杆菌野生株的存活率明显高于TnaA缺失株,但在无色氨酸的培养基中,两者之间没有差异。同时研究人员还用微流控技术监测到在卡那霉素处理后,吲哚能明显促进持留菌形成,这表明吲哚信号在诱导大肠杆菌形成持留菌对抗生素耐受的过程中发挥着关键性作用。吲哚信号能通过激活大肠杆菌的氧化应激和噬菌体侵袭压力反应从而对抗生素产生耐受,最终导致了持留菌的形成[33]。吲哚的产生受多种因素如碳源、氮源、pH值、环境压力等影响[34],可能是多种因素使细菌“感受到危机”从而产生吲哚,产生的吲哚再逐步引导细菌转变成持留菌。
4 信号分子之间的协作
3种信号分子有一个共同点,就是它们能使细菌对抗多种抗生素和氧化应激损伤。胞内氧化应激的特征是代谢或呼吸过程中产生的ROS与抗氧化系统的平衡被打破。抗生素发挥抗菌作用时,氧化应激一直被认为是一个重要的因素[19]。尽管最近一些研究对这一理论提出了质疑[35-36],但本文所涉及的3种信号分子在抵抗氧化应激和抗生素这一点上表现出很强的一致性[12-13,33]。这也暗示着它们在代谢通路和作用机制上一定有着密切的联系。NO、H2S和吲哚产生的底物是3种不同的氨基酸,分别为精氨酸、半胱氨酸和色氨酸。胞内高浓度的半胱氨酸会促进有DNA损伤性的芬顿反应[37],而NO能通过影响半胱氨酸来抑制芬顿反应[18]。产生吲哚的色氨酸酶TnaA与半胱氨酸在调控和代谢上也有密切联系。这表明3种信号分子在代谢的上游就表现出了密切的联系。Shatalin等还探究了NO和H2S对抗氧化应激和抗生素过程中的协同作用,他们发现无论是NO还是H2S缺失后,另一种信号分子的表达量在抗生素的压力下会比野生株更高。这表明,NO和H2S对抗氧化应激和抗生素时具有协同作用,并在一定程度上能够相互补偿[13]。而在吲哚的作用下,大肠杆菌的NO反应系统也同样被激活,从而协同抵抗氧化应激和抗生素[14]。尽管有些细菌并不能产生3种信号分子,但它们有可能进化出类似的补偿机制。如沙门菌不能产生NO,但它能利用宿主产生的NO降低自身呼吸强度,从而减少氨基糖苷类抗生素的摄入量[38]。更有意思的是,沙门菌甚至能够拦截大肠杆菌的吲哚信号,通过诱发氧化应激反应或激活药物外排基因从而达到对抗生素耐受[39-40]。以上研究表明,3种信号分子很可能在对抗氧化应激和抗生素时具有某种共同的作用机制,在不同的情况下发挥作用,却又能相互协作相互影响(图1)。
5 信号小分子导致细菌耐药的特征
一般研究耐药菌都会集中在许多与某种特定抗生素相关的耐药基因,这种通过基因突变或基因水平转移对某种药物产生耐药性的情况会造成整个群体都变成耐药株。因为抗生素的筛选会淘汰不耐药的菌株,含有耐药基因的菌株就被选择出来。但是信号分子导致的群体耐药则是完全不同的情况,极少量菌株会产生一些信号分子并且分泌到胞外,它们将通过激活药物外排泵或抗氧化酶等多种机制使得整个菌群产生耐药。因此,菌群的多样性被保存下来,并且绝大多数菌株并不具有太强的耐药性。这种情况已在上述研究中被证实,耐药菌株产生的吲哚能够保护菌群中不耐药的菌株[14],而大肠杆菌野生株产生的H2S也能保护3MST缺失的突变株[13]。NO、H2S和吲哚这些信号小分子能够很方便地跨膜运输,在菌群中发挥功能。少量个体的利他行为就能以最小的代价使群体对外界抗生素产生耐受。如果外界抗生素压力只是暂时的,那么这样的策略就能最大限度地保留种群的多样性。这种利他行为是一种类似亲缘选择的现象,既保证了菌群的生存,又保留了探索更有利突变的空间,符合微生物的社会进化理论[41]。
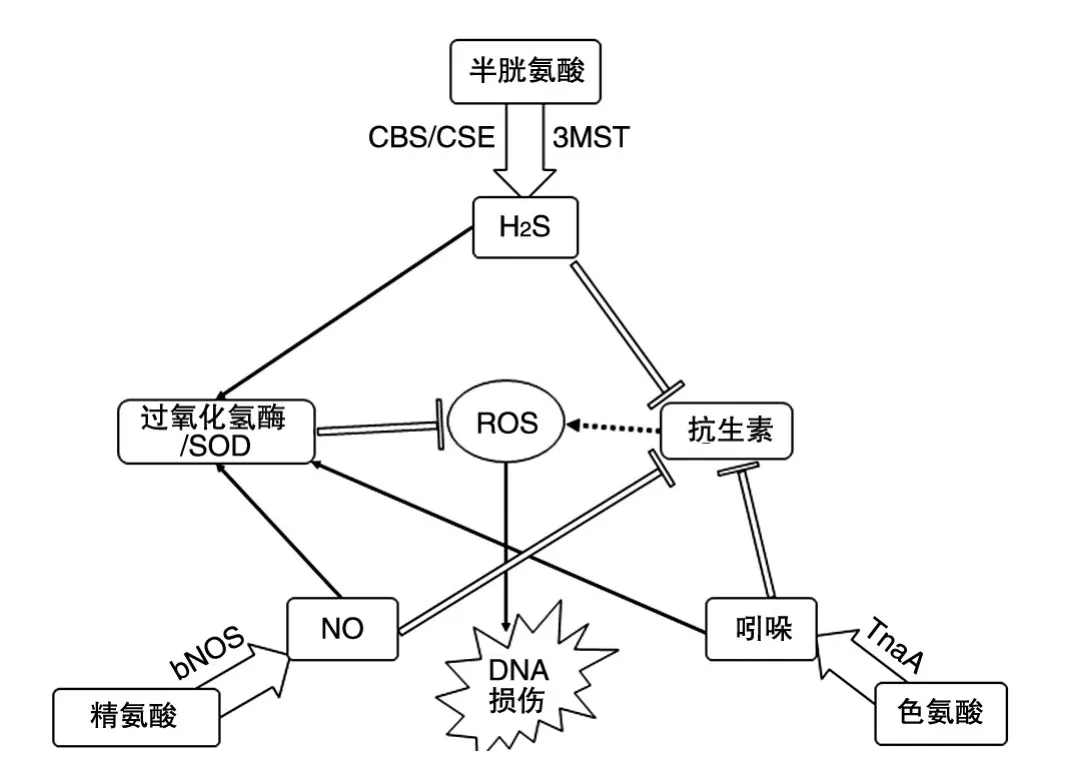
图1 3种信号分子的耐药机制和协同作用
6 结语
细菌信号小分子帮助细菌抵御抗生素,是对目前细菌耐药理论和模型的一个非常重要的补充,有助于我们更全面地理解细菌耐药的机制。更重要的是,这些新发现能帮助我们扩大抗生素药物筛选的范围,通过抑制这些生物小分子发挥作用研制新型抗生素或抗生素的佐剂。目前已发现添加bNOS的抑制物能使细菌变得对抗生素更加敏感[42-43]。但是,目前对这些信号小分子产生耐药的具体分子机制还不是非常清楚,尤其是多种小分子是否有非常明确的共同通路或耐药机制。还有许多问题亟待广大学者去攻破,但这项研究对抗生素的研发、对人类的生命健康安全具有重大意义。
[1] The dangers of hubris on human health[R].World Economic Forum:Global Risks 2013.
[2] Hamad B.The antibiotics market[J].Nat Rev Drug Discovery,2010,9(9):675-676.
[3] Clatworthy A E,Pierson E,Hung D T.Targeting virulence:a new paradigm forantimicrobialtherapy[J].NatChem Biol,2007,3(9):541-548.
[4] Jacoby G A.β-lactamase nomenclature[J].Antimicrob Agents Chemother,2006,50(4):1123-1129.
[5] Jones L A,McIver C J,Kim M J,et al.The aadB gene cas⁃sette is associated with blaSHV genes in Klebsiella species producing extended-spectrum beta-lactamases[J].Antimicrob Agents Chemother,2005,49(2):794-797.
[6] Hernandez-Alles S,Benedi V J,Martinez-Martinez L,et al.Development of resistance during antimicrobial therapy caused by insertion sequence interruption of porin genes[J].Antimi⁃crob Agents Chemother,1999,43(4):937-939.
[7] Anderl J N,Zahller J,Roe F,et al.Role of nutrient limita⁃tion and stationary-phase existence in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin[J].Antimi⁃crob Agents Chemother,2003,47(4):1251-1256.
[8] Piddock L J.Multidrug-resistance efflux pumps-not just for resistance[J].Nat Rev Microbiol,2006,4(8):629-636.
[9] Luhachack L,Nudler E.Bacterial gasotransmitters:an innate defense against antibiotics[J].Curr Opin Microbiol,2014,21C:13-17.
[10]Lee H H,Collins J J.Microbial environments confound antibi⁃otic efficacy[J].Nat Chem Biol,2012,8(1):6-9.
[11]Bernier S P,Letoffe S,Delepierre M,et al.Biogenic ammo⁃nia modifies antibiotic resistance at a distance in physically separated bacteria[J].Mol Microbiol,2011,81(3):705-716.
[12]Gusarov I,Shatalin K,Starodubtseva M,et al.Endogenous ni⁃tric oxide protects bacteria against a wide spectrum of antibi⁃otics[J].Science,2009,325(5946):1380-1384.
[13]Shatalin K,Shatalina E,Mironov A,et al.H2S:a universal defense againstantibioticsin bacteria[J].Science,2011,334(6058):986-990.
[14]Lee H H,Molla M N,Cantor C R,et al.Bacterial charity work leads to population-wide resistance[J].Nature,2010,467(7311):82-85.
[15]Schairer D O,Chouake J S,Nosanchuk J D,et al.The poten⁃tial of nitric oxide releasing therapies as antimicrobial agents[J].Virulence,2012,3(3):271-279.
[16]Filippovich S Y.Bacterial NO synthases[J].Biochemistry(Mos⁃cow),2010,75(10):1217-1224.
[17]Shatalin K,Gusarov I,Avetissova E,et al.Bacillus anthracisderived nitric oxide is essential for pathogen virulence and survival in macrophages[J].Proc Natl Acad Sci USA,2008,105(3):1009-1013.
[18]Gusarov I,Nudler E.NO-mediated cytoprotection:instant ad⁃aptation to oxidative stress in bacteria[J].Proc Natl Acad Sci USA,2005,102(39):13855-13860.
[19]Kohanski M A,Dwyer D J,Hayete B,et al.A common mech⁃anism of cellular death induced by bactericidal antibiotics[J].Cell,2007,130(5):797-810.
[20]van Sorge N M,Beasley F C,Gusarov I,et al.Methicillin-re⁃sistant Staphylococcus aureus bacterial nitric-oxide synthase affects antibiotic sensitivity and skin abscess development[J].J Biol Chem,2013,288(9):6417-6426.
[21]Medani M,Collins D,Docherty N G,et al.Emerging role of hydrogen sulfide in colonic physiology and pathophysiology[J].Inflammatory Bowel Dis,2011,17(7):1620-1625.
[22]Wang R. Physiological implications of hydrogen sulfide: a whiff exploration that blossomed[J].Physiol Rev,2012,92(2):791-896.
[23]Schemann M,Grundy D.Role of hydrogen sulfide in visceral nociception[J].Gut,2009,58(6):744-747.
[24]Heinzinger N K,Fujimoto S Y,Clark M A,et al.Sequence analysis of the phs operon in Salmonella typhimurium and the contribution of thiosulfate reduction to anaerobic energy metabolism[J].J Bacteriol,1995,177(10):2813-2820.
[25]Huang C J,Barrett E L.Sequence analysis and expression of the Salmonella typhimurium asr operon encoding production of hydrogen sulfide from sulfite[J].J Bacteriol,1991,173(4):1544-1553.
[26]Flannigan K L,McCoy K D,Wallace J L.Eukaryotic and prokaryotic contributions to colonic hydrogen sulfide synthesis[J].Am J PhysiolGastrointestLiverPhysiol,2011,301(1):G188-193.
[27]Linden D R.Hydrogen sulfide signaling in the gastrointestinal tract[J].Antioxidants Redox Signaling,2014,20(5):818-830.
[28]Lee J H,Lee J.Indole as an intercellular signal in microbial communities[J].FEMS Microbiol Rev,2010,34(4):426-444.
[29]Lee J,Jayaraman A,Wood T K.Indole is an inter-species biofilm signal mediated by SdiA[J].BMC Microbiol,2007,7:42.
[30]Newton W A,Snell E E.Formation and interrelationships of tryptophanase and tryptophan synthetases in Escherichia coli[J].J Bacteriol,1965,89:355-364.
[31]Hirakawa H,Inazumi Y,Masaki T,et al.Indole induces the expression of multidrug exporter genes in Escherichia coli[J].Mol Microbiol,2005,55(4):1113-1126.
[32]Kobayashi A,Hirakawa H,Hirata T,et al.Growth phase-de⁃pendent expression of drug exporters in Escherichia coli and its contribution to drug tolerance[J].J Bacteriol,2006,188(16):5693-5703.
[33]Vega N M,Allison K R,Khalil A S,et al.Signaling-mediat⁃ed bacterial persister formation[J].Nat Chem Biol,2012,8(5):431-433.
[34]Han T H,Lee J H,Cho M H,et al.Environmental factors af⁃fecting indole production in Escherichia coli[J].Res Microbi⁃ol,2011,162(2):108-116.
[35]Keren I,Wu Y,Inocencio J,et al.Killing by bactericidal an⁃tibiotics does not depend on reactive oxygen species[J].Sci⁃ence,2013,339(6124):1213-1216.
[36]Liu Y,Imlay J A.Cell death from antibiotics without the in⁃volvementofreactive oxygen species[J].Science,2013,339(6124):1210-1213.
[37]Park S,Imlay J A.High levels of intracellular cysteine pro⁃mote oxidative DNA damage by driving the fenton reaction[J].J Bacteriol,2003,185(6):1942-1950.
[38]McCollister B D,Hoffman M,Husain M,et al.Nitric oxide protects bacteria from aminoglycosides by blocking the energydependent phases of drug uptake[J].Antimicrob Agents Che⁃mother,2011,55(5):2189-2196.
[39]Vega N M,Allison K R,Samuels A N,et al.Salmonella ty⁃phimurium intercepts Escherichia coli signaling to enhance an⁃tibiotic tolerance[J].Proc Natl Acad Sci USA,2013,110(35):14420-14425.
[40]Blair J M,Cloeckaert A,Nishino K,et al.Alternative explana⁃tion for indole-induced antibiotic tolerance in Salmonella[J].Proc Natl Acad Sci USA,2013,110(48):E4569.
[41]West S A,Griffin A S,Gardner A,et al.Social evolution the⁃ory for microorganisms[J].Nat Rev Microbiol,2006,4(8):597-607.
[42]Holden J K,Li H,Jing Q,et al.Structural and biological studies on bacterial nitric oxide synthase inhibitors[J].Proc Natl Acad Sci USA,2013,110(45):18127-18131.
[43]Holden J K,Kang S,Hollingsworth S A,et al.Structurebased design of bacterial nitric oxide synthase inhibitors[J].J Med Chem,2015,58(2):994-1004.
