沙眼衣原体质粒调控基因的上游序列有广泛的ChxR结合位点
2015-05-04李鹏于永慧王涛何君朱虹端青宋立华
李鹏,于永慧,王涛,何君,朱虹,端青,宋立华
1.病原微生物生物安全国家重点实验室,军事医学科学院 微生物流行病研究所,北京 100071;
2.解放军第264医院,山西 太原 030001
沙眼衣原体(Chlamydia trachomatis,Ct)是沙眼和最常见的细菌性性传播疾病的病原体。在我国,致盲性沙眼已多年未见报道,Ct导致的泌尿生殖道感染是常见病,特别是女性感染Ct后易发生盆腔炎、流产、不孕不育等。近年来有报道称Ct感染还可能是宫颈癌的又一病因。由于Ct为专性细胞内寄生,培养和纯化困难,Ct疫苗研究一直进展缓慢。近年来Ct减毒活疫苗研究有突破,在短尾猴沙眼模型上质粒缺失的Ct A2497株是良好的沙眼减毒活疫苗[1]。在小鼠模型上,Ct L2型、F型和鼠衣原体(与Ct亲缘性最高的动物衣原体)的质粒缺失株也已减毒,这说明质粒是Ct的关键毒力因子[2-4]。
质粒缺失Ct的共有表型是包含体中无糖原积累。Ct质粒约7.5 kb,编码8个蛋白(Pgp1~Pgp8),其致病机制主要是通过Pgp4调控Pgp3及至少8个染色体基因(包括糖原合成酶基因glgA、CT084)的转录表达[5]。Pgp4的调控机制还不清楚。有意思的是,Koo等报道了ChxR在大肠杆菌系统中可以激活转录质粒调节基因CT084[6]。ChxR是保守的衣原体蛋白,Koo等认为是衣原体特定发育时期的全局转录调控因子,可能与衣原体的潜伏感染相关,而质粒缺失的A2497的典型感染特征是短时感染。这都说明ChxR可能也参与Pgp4调节基因的转录调控。
ChxR是保守的衣原体蛋白,与大肠杆菌双组分信号转导系统的CpxR同源,但它不含天门冬氨酸磷酸化位点,是非典型的OmpR/PhoB家族的反应调节蛋白。这类调控因子存在于衣原体、螺杆菌、粘球菌、链霉菌和聚球藻中,其转录调节活性不受磷酸化调控,作用机制还不清楚。ChxR是自身激活转录因子[7],其结构已被解析[8-9],但对其调控靶标和机制的了解仍很有限。我们利用凝胶阻滞实验,分析ChxR与4个Pgp4调节基因上游区的相互作用,有助于理解衣原体质粒及ChxR的转录调控机制。
1 材料与方法
1.1 材料
沙眼衣原体 L2(434),大肠杆菌 DH5α、BL21(DE3),载体pCR2.1、pET-21b由本室保存;限制性内切酶、高保真DNA聚合酶为NEB公司产品;DNA提取试剂盒、质粒纯化试剂盒、PCR产物纯化试剂盒为Qiagen公司产品;LightShift化学发光EMSA试剂盒、DNA 3'端生物素标记试剂盒购自Thermo Fish⁃er Scientific公司;TALON金属亲和介质购自Clon⁃tech公司。
1.2 ChxR的重组表达和纯化
以沙眼衣原体L2(434)基因组为模板,采用引物 ChxRf(CCggAATTCggCggggCCTAAACATgTg)、ChxRr(CCgCTCgAgTCTAgAAAgCTTTgTATCTTgTTg Ag)和高保真DNA聚合酶扩增ChxR基因。PCR产物经EcoRⅠ和XhoⅠ酶切后,与经同样酶切处理的载体pET-21b连接,产物转化大肠杆菌BL21(DE3),于37℃在含100 μg/mL青霉素的LB培养基中培养至 D600nm为 0.8,加入 1 mmol/L IPTG 继续培养 2 h后,用TALON介质纯化重组蛋白(洗脱蛋白中加入25%甘油,-80oC冻存)。
1.3 衣原体基因上游序列的探针制备
以L2(434)基因组为模板,用表1中的引物和高保真DNA聚合酶扩增5个基因(glgA、CTL0071、CTL0305、CTL0339、chxR)的上游序列(约150 bp),将PCR产物克隆入载体pCR2.1,经测序验证插入序列后摇菌纯化重组质粒,用BstXⅠ单酶切将质粒分割为载体序列和插入序列2个片段,该DNA混合物直接进行3'端生物素标记,氯仿异戊醇抽提后,加无核酸酶水将插入序列亦即探针的浓度稀释至50 fmol/μL,-20oC保存。
1.4 引物延伸实验
用引物TTACATTTAAggggCATCTTACC和CTC CCAggCCTCCggCTTT(与glgA mRNA互补),PCR扩增glgA的上游序列和部分ORF序列共289 bp,扩增产物纯化后作为测序模板。将与mRNA互补的引物的5'端用[γ-32P]ATP(5000 Ci/mmol)进行放射性标记,退火后用逆转录酶合成cDNA。逆转录产物与mRNA互补引物的测序条带进行8 mol/L尿素-6%聚丙烯酰胺凝胶电泳,放射自显影后,通过引物延伸条带的位置确定转录起始位点。
1.5 寡核苷酸探针制备
为鉴别ChxR与glgA上游序列的结合位点,设计合成5条60 bp的寡核苷酸探针,每条探针有30 bp的重叠序列(转录因子的核酸结合序列通常小于30 bp),共覆盖了ATG上游150 bp长度。探针均为双链,由2条3'端标记生物素的单链DNA等比例混合后缓慢退火制成。本研究所需引物或探针均由Life Technologies公司合成。
1.6 凝胶阻滞实验
所有操作按照LightShift化学发光EMSA试剂盒(含化学发光核酸检测模块)的操作说明书进行。反应体系:1 μL重组ChxR(500 ng,含25%甘油),1×结合缓冲液,2.5%甘油,5 mmol/L MgCl2,50 ng/μL Poly(DI-DC),0.05%NP-40,4 pmol/L未标记探针,20 fmol/L标记探针,加双蒸水至10 μL总体积。
2 结果
2.1 ChxR的重组表达和纯化
在大肠杆菌中,重组ChxR若在自身ORF的ATG起始翻译时,重组蛋白几乎全部形成包含体,不利于蛋白纯化;若利用pET-21b载体的ATG起始翻译,则蛋白的溶解性明显提高,可获得高纯度的重组蛋白(图1)。重组ChxR的N端有T7标签,C端有6×His标签,可用TALON试剂较好地得到纯化。纯化的重组蛋白在冻存前要加入终浓度为25%的甘油,否则会形成聚体沉淀,这可能与此蛋白易形成二聚体有关。图1中第1次洗脱的ChxR浓度约为500 ng/μL,其浓度和纯度均满足凝胶阻滞实验的要求。
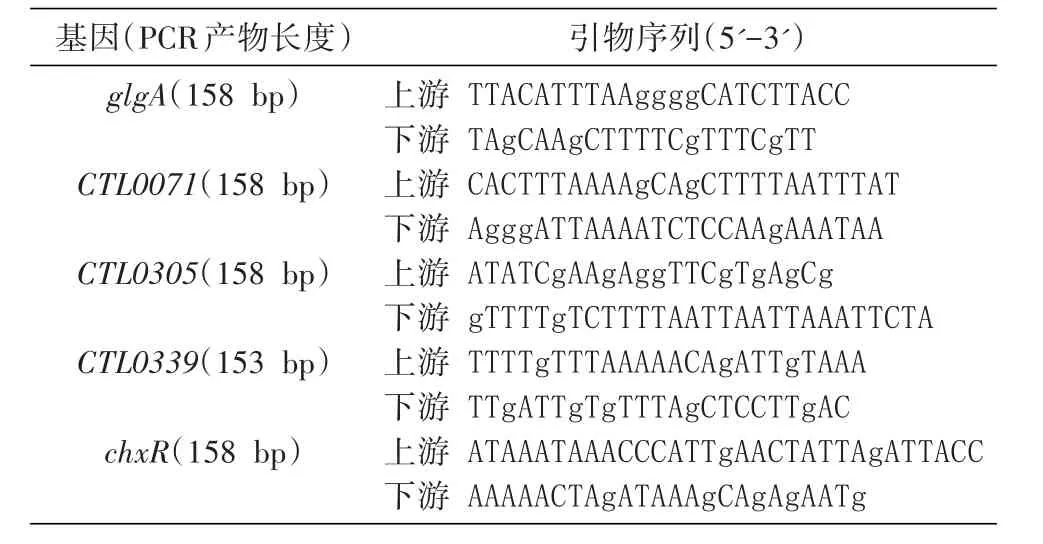
表1 构建glgA、CTL0071、CTL0305、CTL0339、chxR探针的各组引物
2.2 质粒调控基因的上游区含有多个ChxR结合位点
为鉴定ChxR是否与质粒调控基因的上游序列有相互作用,我们利用上述纯化的重组ChxR进行了凝胶阻滞实验(图2)。文献已报道ChxR是自身激活转录因子[7],可识别并结合自身基因的上游区,我们因此以chxR上游区作为阳性对照探针。另外,Koo等报道ChxR可激活CTL0339(CT084)的转录[6],CTL0339探针也因此应与ChxR存在相互作用。图2中chxR探针有4条明显的阻滞条带,这与文献报道一致。让人吃惊的是,质粒调控基因的上游区也有3~6条不等的阻滞带,其中glgA、CTL0071、CTL0305和CTL0339探针的阻滞条带数分别为6、4、3和5条,说明这几个基因的上游区含有多个ChxR结合位点,ChxR可能参与这几个基因的转录调控。
2.3 引物延伸实验结果
糖原合成代谢在衣原体中高度保守,很可能为衣原体的潜伏感染提供关键能量;另外糖原合成酶基因glgA是惟一功能明确的质粒调控基因,我们因此重点探索了ChxR与glgA的转录之间的可能机制。首先利用引物延伸实验鉴定了glgA的转录起始位点(transcriptional start site,TSS),这对下一步理解ChxR与glgA上游区的相互作用很关键。基于生物信息学预测,glgA上游区有一个σ70启动子(TTg ATTATTTTTgTTAAAAgAAATAAT),其TSS因此可能位于-31(glgA ORF中ATG的A设为+1位,以下均同)(图3)。引物延伸实验所需的引物因此设计在此位点的下游75 bp处,实验结果表明glgA的TSS位于-54位,这与推测的TSS不同(图4)。此TSS位于回文序列内部,其转录起始可能受阻遏蛋白调控。
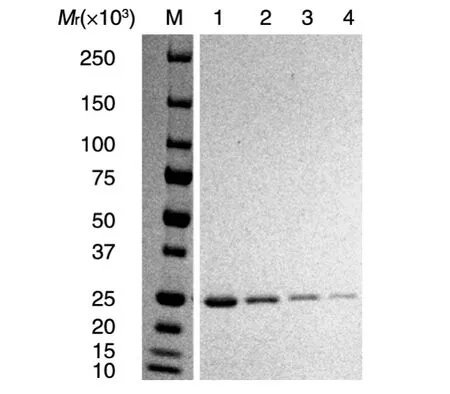
图1 亲和层析纯化的重组ChxR电泳图谱M:Precision plus protein prestained standards;1~4:分别为层析柱的第1~4次洗脱结果
2.4 glgA启动子的上下游区均有ChxR结合位点
为定位ChxR与glgA上游区的结合位点,我们设计了5条长60 bp、重叠30 bp的寡核酸探针,根据TSS推测的启动子位于第2和第3条探针(图5A)。图5B结果表明探针1~5分别有2、2、1、0和1条阻滞条带,很明显多数ChxR结合位点位于启动子和启动子下游区。虽然文献报道ChxR是全局转录增强子,但本实验强烈提示ChxR可能是glgA的转录抑制子。
3 讨论
衣原体感染特别是沙眼衣原体感染是临床常见病,常导致沙眼、不孕不育、动脉粥样硬化、肺炎等。衣原体的致病机制仍有待探讨,最新研究表明衣原体质粒蛋白Pgp4调控的染色体基因与致病性紧密相关。即使在质粒缺失的衣原体毒株如肺炎衣原体和鹦鹉热衣原体中,这些染色体基因同样保守存在。这些染色体基因的致病机理是目前衣原体领域的研究热点。本研究探索了这些染色体基因的转录调控与转录调控因子ChxR间的关联。
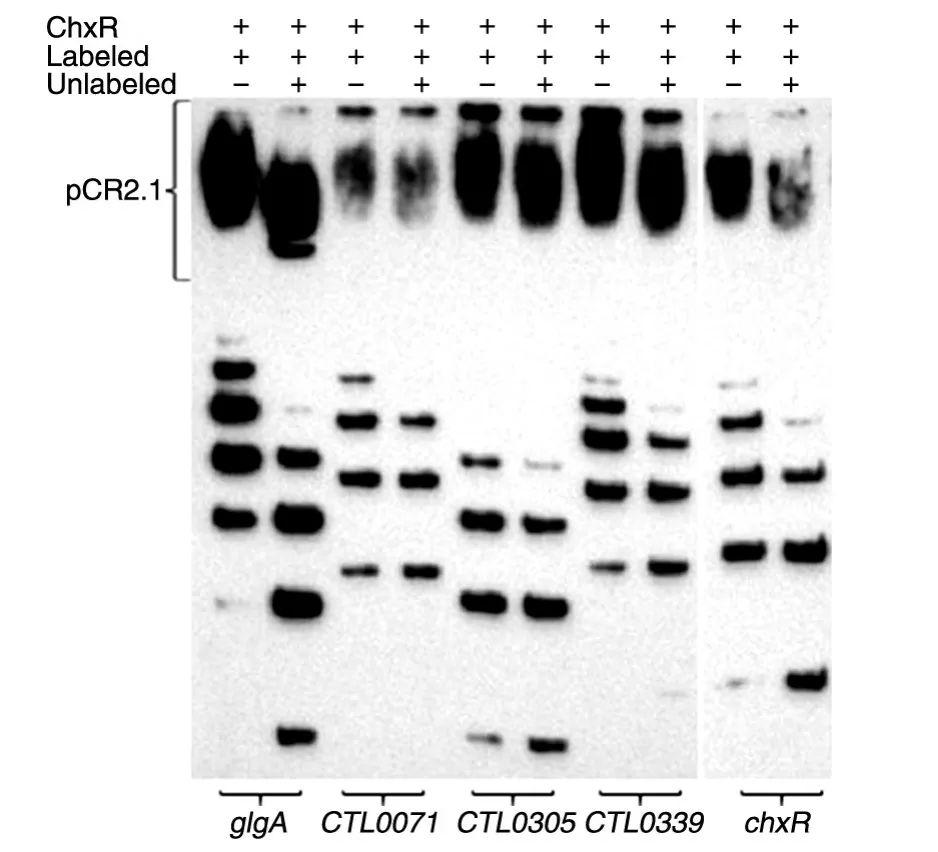
图2 凝胶阻滞实验

图3 生物信息学预测的glgA启动子和转录起始位点pgsA、glgA、tRNA示衣原体基因的位置和方向;下面的核酸序列标注了预测的glgA启动子、TSS、RBS(核糖体结合位点)和TL(翻译起始位点)
Pgp4调控染色体基因表达的机制和ChxR的功能都还不清楚,我们通过体外凝胶阻滞实验发现Pgp4调控的4个染色体基因的上游区有多个ChxR结合位点,提示ChxR参与这些染色体基因的转录调控;通过鉴定glgA mRNA的转录起始位点和设计寡核苷酸探针进行凝胶阻滞实验,我们进一步发现ChxR在glgA转录中可能起到阻遏蛋白的作用。这些研究提示glgA的转录表达可能是多个转录调控因子协同作用的结果。
图4 引物延伸实验
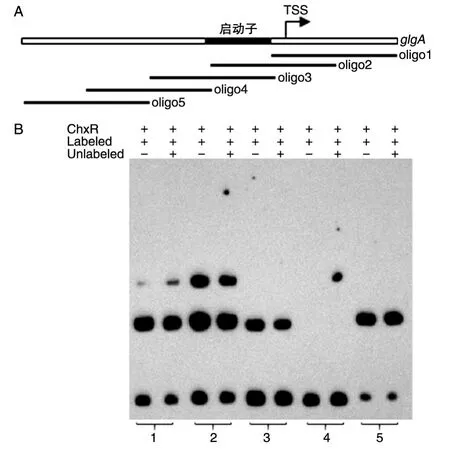
图5 寡核苷酸探针oligo1~5与ChxR的凝胶阻滞实验A:oligo1~5为5条寡核苷酸探针,其中1号探针紧邻glgA ORF;B:oligo1~5分别有2、2、1、0和1条阻滞条带
本研究除了提示ChxR参与质粒调控基因表达外,重要的是发现ChxR有广泛的DNA结合活性,但我们没能找到其识别结合的DNA序列特征,很可能有更多的基因受其调控。我们的实验结果支持ChxR可能是全局调控因子,该蛋白可能同时为转录增强子和抑制子。新出现的衣原体遗传操作技术为下一步深入研究ChxR和Pgp4协同调控基因转录的机制提供了重要工具[5,10-13]。
[1] Kari L,Whitmire W M,Olivares-Zavaleta N,et al.A live-at⁃tenuated chlamydial vaccine protects against trachoma in non⁃human primates[J].J Exp Med,2011,208(11):2217-2223.
[2] Sigar I M,Schripsema J H,Wang Y,et al.Plasmid deficien⁃cy in urogenital isolates of Chlamydia trachomatis reduces in⁃fectivity and virulence in a mouse model[J].Pathog Dis,2014,70(1):61-69.
[3] O'Connell C M,Ingalls R R,Andrews C W Jr,et al.Plas⁃mid-deficient Chlamydia muridarum fail to induce immune pa⁃thology and protectagainstoviductdisease[J].JImmunol,2007,179(6):4027-4034.
[4]Carlson J H,Whitmire W M,Crane D D,et al.The Chlamyd⁃ia trachomatis plasmid is a transcriptional regulator of chromo⁃somal genes and a virulence factor[J].Infect Immun,2008,76(6):2273-2283.
[5] Song L,Carlson J H,Whitmire W M,et al.Chlamydia tracho⁃matis plasmid-encoded Pgp4 is a transcriptional regulator of virulence-associated genes[J].Infect Immun,2013,81(3):636-644.
[6] Koo I C,Walthers D,Hefty P S,et al.ChxR is a transcrip⁃tionalactivatorin Chlamydia[J].ProcNatlAcad SciUSA,2006,103(3):750-755.
[7] Hickey J M,Weldon L,Hefty P S.The atypical OmpR/PhoB response regulator ChxR from Chlamydia trachomatis forms ho⁃modimers in vivo and binds a direct repeat of nucleotide se⁃quences[J].J Bacteriol,2011,193(2):389-398.
[8] Hickey J M,Lovell S,Battaile K P,et al.The atypical re⁃sponse regulatorprotein ChxR hasstructuralcharacteristics and dimer interface interactions that are unique within the OmpR/PhoB subfamily[J].J Biol Chem,2011,286(37):32606-32616.
[9] Barta M L,Hickey J M,Anbanandam A,et al.Atypical re⁃sponse regulator ChxR from Chlamydia trachomatis is structur⁃ally poised for DNA binding[J].PLoS One,2014,9(3):e91760.
[10]Wang Y,Kahane S,Cutcliffe L T,et al.Development of a transformation system for Chlamydia trachomatis:restoration of glycogen biosynthesis by acquisition of a plasmid shuttle vec⁃tor[J].PLoS Pathog,2011,7(9):e1002258.
[11]Kari L,Goheen M M,Randall L B,et al.Generation of tar⁃geted Chlamydia trachomatis null mutants[J].Proc Natl Acad Sci USA,2011,108(17):7189-7193.
[12]Nguyen B D,Valdivia R H.Virulence determinants in the ob⁃ligate intracellularpathogen Chlamydia trachomatis revealed by forward genetic approaches[J].Proc Natl Acad Sci USA,2012,109(4):1263-1268.
[13]Song L,Carlson J H,Zhou B,et al.Plasmid-mediated trans⁃formation tropism of chlamydial biovars[J].Pathog Dis,2014,70(2):189-193.
