广东康禾自然保护区亚热带常绿阔叶林立木结构及碳密度分布特征
2015-03-02李文斌胡砚秋徐明锋苏志尧吕远辉
李文斌,胡砚秋,徐明锋,苏志尧,陈 卫,吕远辉
(1.华南农业大学林学院,广州510642;2.广东省东源县林业局,河源 517000)
广东康禾自然保护区亚热带常绿阔叶林立木结构及碳密度分布特征
李文斌1,胡砚秋1,徐明锋1,苏志尧1,陈 卫2,吕远辉2
(1.华南农业大学林学院,广州510642;2.广东省东源县林业局,河源 517000)
在广东东源县康禾自然保护区林分起源相同的典型亚热带常绿阔叶林区域,在物种多样性有明显差异的3个群落分别设置1 hm2的样地,研究异质多样性下亚热带常绿阔叶林的立木结构及碳密度分布特征.研究结果表明,随着植物群落丰富度或个体数的增大,二次平均胸径和平均胸径指标都逐渐减小,而样方立木的总胸径和胸高断面积则逐步增加.生态优势度较低但多样性指数较高的群落1和群落2内,植物种类或个体数的增加使群落蓄积量得以增加,群落3则不然.生态优势度高、物种多样性相对较低的群落,林分蓄积和碳密度更高.这项研究结果提示,立木结构指标与林分碳储量没有直接相关关系,而立木多样性高的群落碳储量不一定高;大径级和具有生态优势的种类对亚热带常绿阔叶林蓄积量和碳储量的保持与增加具有主要贡献作用.
立木结构;碳密度;多样性;蓄积量
在全球生物多样性丧失和气候变化的背景下,可持续性的生物多样性保护是森林管理的重要目标之一;而森林碳汇也正成为持续关注的热点[1],面向林业碳汇的森林碳汇管理有助于缓解全球气候变化[2].全球尺度上,处在不同气候带的森林在碳储量上存在较大差别[3],不同的森林类型间碳密度也不尽相同.从森林生产力的角度看,较高的森林生物量和碳储量是森林高生产力的表现.但在不同空间尺度上,自然群落物种多样性与生产力的相关性格局表现各异,一定的空间尺度下,高生产力意味着更多的生态位选择以及更少的物种迁移,使得物种多样性提高,反之,物种多样性的提高也可带来生产力的提高;而在小的空间尺度上,多样性与生产力的关系则变得复杂[4].实际上,生物多样性与生产力关系方面的研究存在许多争议,物种多样性高,生产力不一定就高[5].关于林分物种多样性与林分生物量的关系,有报道指出相同起源但互相间隔的桉树植物群落乔木层植物多样性与乔木层生物量成极显著的正相关[6];相同立地条件下,植物多样性高的阔叶次生林地上部分生物量高于杉木纯林[7];在对不同白桦林进行植物多样性和生物量的比较中,也能得出植物多样性与生物量正相关性的结果[8];另外,植物多样性较高的热带顶极森林群落在植被碳含量上却不一定高于温带地带性顶极森林群落[9].本文通过比较起源相同但植物多样性有明显差异的不同常绿阔叶林立木结构和碳密度的分布情况,研究植物多样性、立木结构与林分碳密度之间的关联性,为亚热带森林保育和碳汇管理工作提供参考.
1 研究地概况
样地设置在河源市东源县东南部的广东康禾省级自然保护区(北纬23°44'37″-23°52'16″,东经115°04'27″-115°09'41″).该区地处中亚热带和南亚热带交界处,年均气温21.1℃,年均降雨量1912 mm,降雨量主要中在4-9月.保护区南北长约14 km,东西宽约6 km,总面积6 484.8 hm2;地貌多样,属半山区半丘陵地带,最高峰白石岗,海拔 893.3 m[10,11].
2 研究方法
2.1 群落调查
在起源相同的亚热带常绿阔叶林内,通过线路调查,选取3个具有共同建群树种和相近环境因子(表1),但在植物多样性上存在差异的植物群落,分别命名为群落1、群落2和群落3,在每个群落内设置面积为1 hm2的标准样地.调查时,在群落内划分100个10 m×10 m的样方,测量胸径≥1 cm立木的胸径、高度、冠幅和枝下高,并鉴定和记录种名.
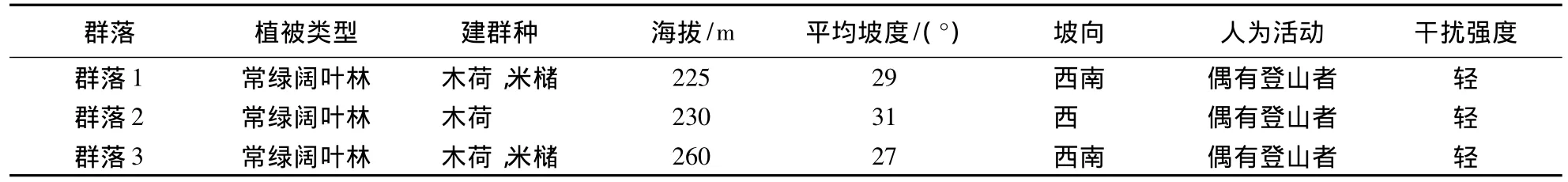
表1 群落基本信息Table1 Community general information
2.2 数据计算与分析
2.2.1 物种多样性指标 采用下面的公式计算物种多样性指数和生态优势度.

上述公式中,Pi=ni N,为第i个物种的个体与总个体数的比值,N为所有种的个体总数,s为种数,ni为第i种的个体数,Nmax为样地或样方中最优势种的个体数[12].
2.2.2 重要值 重要值(Importance value,IV)计算采用公式:

式中FR为相对频度(relative frequency,FR),DR为相对密度(relative density,DR),BR为相对优势度即相对胸高断面积(relative basal area,BR).
2.2.3 蓄积量与碳密度 根据《广东省森林资源调查常用数表》[13]查找各类立木的二元立木材积公式:

公式中,V代表立木材积(m3),D为胸径(cm),H为树高(m),所有立木材积之和则为群落的林分蓄积量.
基于林分蓄积量,计算群落生物量,使用公式[14]:

其中,X为蓄积量(m3),Y为生物量(t).由于群落内各种类含碳系数接近,由生物量计算碳储量时,取含碳系数0.5[14];群落碳密度为单位面积的碳储量.
2.2.4 平均胸径 林分算术平均胸径和二次平均胸径是测度立木平均直径的常用指标[15],按以下公式计算:

重要值、多样性指数的计算以及多响应置换过程分析(MRPP,multi-response permutation procedures)在PC-ORD 6.0生态分析软件包[16]中完成,其中,多响应置换过程分析用于揭示不同植物群落划分具有差异性的统计学意义[17].群落结构相关性分析在多元统计分析软件STATISTICA 8.0上运行.
3 结果与分析
3.1 物种组成和多样性
三个群落起源相同,但在植物多样性上存在一定差别,多样性为群落1>群落2>群落3,其中群落3的植物多样性与群落1和2相差较大.在群落优势种个体数大小和优势度上,则表现为群落3大于群落2,远大于群落1,样地立木个体数也是群落3最多(表2).
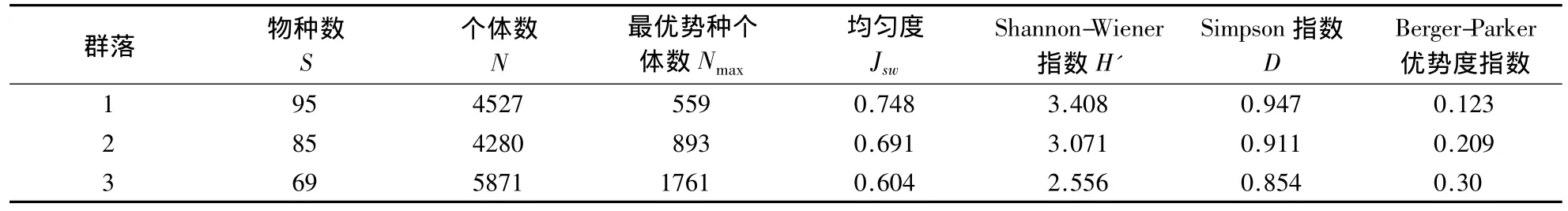
表2 群落组成和多样性Table 2 Community composition and diversity
多响应置换过程分析的方法揭示了不同植物群落划分的统计学意义,其中P值表明了分组的显著性,而T值越负则表示组间分离越明显,A值大小则表示组内一致性的高低.对3个群落组成异质性的多响应置换过程分析说明,各群落分组具有显著差异性和组内一致性(P<0.0001,A=0.0831).群落间的配对比较显示,群落两两之间比较具有显著的差异性(P<0.0001),群落1和群落3之间的T值最小(T=-64.5730),A最大(A=0.0776),表现出的组间分离最明显,组内一致性也最高;其次是群落2和群落3(T= -45.6813,A=0.0627),群落1 和群落2(T= -45.4847,A=0.0513).
在每个群落选取10个相对优势种(表3)比较重要值大小,群落1中重要值最大的前5个种类是木荷Schima superba、米槠Castanopsis carlesii、罗伞树Ardisia quinquegona、栲Castanopsis fargesii、黄樟Cinnamomum porrectum;群落2为米槠、罗伞树、多毛茜草树Aidia pycnantha、木荷、栲;群落3为米槠、木荷、罗伞树、鼠刺Itea chinensis、栲.三个植物群落在优势种类上表现出相似性,乔木层共同优势种类为米槠、木荷、栲,林下优势种类为罗伞树.重要值是各植物种类在群落中的影响程度和地位的综合反映,从单个指标上看,则有不同的排序,比如群落1中米槠、栲、黄樟、黄杞Engelhardtia roxburghiana、红锥Castanopsis hystrix、山乌桕Sapium discolor对该群落的胸高断面积贡献较大,实际上,红锥、山乌桕在样方中的频度与多度相比其余8个种类要小,而罗伞树、杨桐Adinandramillettii、毛冬青Ilex pubescens等林下种类出现频度和多度很高,但是胸高断面积很小,群落2和群落3也是如此,表明胸径普遍较大的种类在增加群落蓄积上的潜力更大.

表3 群落优势种的结构特征Table 3 Structural characters of community dominant species
3.2 群落结构
将每个群落内的物种数分别与平均胸径、总胸径、二次平均胸径和胸高断面积作相关性分析(图1),从中可见物种数和平均胸径、二次平均胸径都呈显著的负相关关系;物种数分别与总胸径和胸高断面积成显著正相关关系和线性相关性较低的正相关关系.分析群落样方内物种个体数与平均胸径、总胸径、二次平均胸径、胸高断面积的相关性,得到个体数与平均胸径、二次平均胸径都成显著的负相关关系,个体数与总胸径成显著的正相关关系,与胸高断面积成线性相关性较低的正相关关系(图2).以上相关性趋势说明随着物种数和个体数的增加,样地内物种个体胸径大小愈加参差不齐,导致了两个平均胸径指标的下降;如果样地内具有更多大径级的立木种类或个体,那么总胸径以及断面积就会更大.进一步说明大径级立木对亚热带常绿阔叶林蓄积量的保持与增进具有主要的作用.

图1 群落样方丰富度与立木胸径指标的关系Fig.1 The relationship between stand richness and DBH index in the community samples

图2 群落样方总多度与立木胸径指标的关系Fig.2 The relationship between total stand abundance and DBH index in the community samples
3.3 立木碳蓄积及其分布
蓄积量大小与立木的胸径大小成正比,作各群落物种数和个体数与其蓄积量的相关性分析时发现,群落1和群落2物种数和个体数分别与其蓄积量成显著的正相关关系,群落3个体数与该群落的蓄积量呈现显著负相关关系,而物种数与群落蓄积量成不显著的负相关(图3).结合表2和表4,总体上群落3优势种类的优势度比群落1和2高,优势种类胸高断面积所占的比重远远大于其它种类,而且各样方的立木结构情况差异较大,作物种或个体数与蓄积量的相关性分析时,所表现的相关性不强;而图3中群落1和群落2的情况说明内部立木结构异质性不强的群落,物种数和个体数的增加有利于群落蓄积量的增长.事实上,3个群落蓄积量、生物量和碳密度大小比对结果为,群落1<群落2<群落3(图4),而物种数,多样性指数的大小比较为群落1>群落2>群落3,物种多度比较为群落2<群落1<群落3,物种优势度大小排序为群落1<群落2<群落3(表2),表明尽管群落3物种丰富度小于群落1和群落2,但优势种类明显,个体数量多且胸径较大,这都使得群落3的蓄积量更高,分别对群落内的各物种计算生物量和碳密度后,结果也是群落3的生物量和碳密度最大.
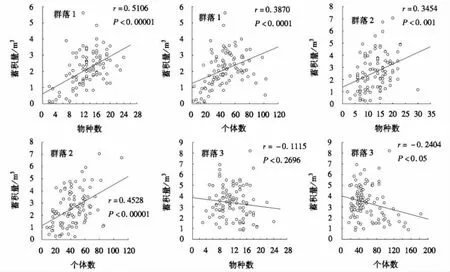
图3 不同群落立木蓄积量与样方多度及丰富度的关系Fig.3 The relationship between stand volume and abundance in different community samples

图4 不同群落立木蓄积量与林分碳储量Fig.4 Stand volume and carbon stock in different communities
4 讨论
群落组成异质性的多响应置换过程分析表明各群落分组具有显著差异性和组内一致性,群落两两配对比较中,群落3与群落1和群落2之间的差异较大,群落3的立木组成中,米槠的优势度明显,胸高断面积远大于其它种类,另外,群落3总体上立木多度最高,物种多样性最低.用每个群落样方内的物种数和个体数分别与平均胸径、二次平均胸径、总胸径和胸高断面积作相关性分析得到的相关性结果表明,当群落内大胸径种类数量一定的情况下,小胸径立木种类和个体的增加使得平均胸径指标降低,而总胸径和断面积指标逐渐增加.作各群落物种数和个体数与其蓄积量的相关性分析时发现,群落1和群落2样方内的物种数和个体数分别与其蓄积量成显著的正相关关系,群落3则不然,该群落物种数和个体数较多的样方,反而蓄积量低,这与大胸径立木多数分布在物种和个体数较少的样方有关.三个群落蓄积量、生物量和碳密度大小比对中,群落1<群落2<群落3,多样性指数的大小比较却是群落1>群落2>群落3,群落3多样性指数最低,但生物量和碳密度最大.以上说明立木结构指标与林分碳储量没有直接相关关系,而立木多样性高的群落碳储量不一定高,说明了该亚热带常绿阔叶林在小尺度空间上多样性、立木结构与群落碳储量的关系较为复杂,同时符合物种多样性不一定与生产力成正相关关系的说法.
我国天然林中以中龄林、成熟林和过熟林的碳密度较高[18],而森林植被碳储量又主要分布在乔木层和灌木层[9,19],亚热带常绿阔叶林碳储量主要集中在乔木层大胸径立木中[20],而不一定在优势种类中.三个群落共同优势种类为米槠、木荷、栲、罗伞树,而蓄积量和碳储量在群落立木中所占比重最大种类有米槠、木荷和栲,特别是群落3,米槠的重要值,多度,胸高断面积都远大于其它种类,对群落蓄积量和碳储量的贡献最大.另外一些种类,比如红锥、南酸枣、华润楠Machilus chinensis等,也具有很大的增加林分蓄积量和碳储量的潜力,相比而言,罗伞树、杨桐、毛冬青等林下种类出现频度和多度很高,但是胸高断面积很小,碳密度低,不过仍是林下碳储量的重要组成部分,其它一些生态优势度不高的种类,特别是稀有种,则促进了群落植物多样性的提高.具有不同物种的植物群落生产力往往是由少数高产种类决定的[21],在碳汇林、近自然林的营造中,一方面注重增加和保护林内植物种类的多样性,另一方面,选择当地的具有碳汇潜力的种类进行种植;对于现有的天然碳汇林,应减少人为砍伐,对大径级乔木进行保留.老龄林立木密度低却主要由大乔木组成,具有显著的碳汇效果[22].
[1]李怒云,宋维明.气候变化与中国林业碳汇政策研究综述[J].林业经济,2006(5):60-64.
[2]CANADELL JG,RAUPACH M R.Managing forests for climate change mitigation[J].Science,2008,320(5882):1456 -1457.
[3]BONAN G B.Forests and climate change:Forcings,feedbacks,and the climate benefits of forests[J].Science,2008,320(5882):1444-1449.
[4]KASSEN R,BUCKLING A,BELL G,et al.Diversity peaks at intermediate productivity in a laboratory microcosm[J].Nature,2000,406(6795):508 -512.
[5]岳天祥.生物多样性研究及其问题[J].生态学报,2001,21(3):462 -467.
[6]温远光,陈放,刘世荣,等.广西桉树人工林物种多样性与生物量关系[J].林业科学,2008,44(4):14-19.
[7]孙德宙,詹有生,盛炜彤,等.江西大茅山常绿阔叶次生林与人工林生物多样性分析[J].林业科学研究,1998,11(4):63-67.
[8]郭其强,张文辉,何景峰,等.黄龙山不同白桦林群落结构特征研究[J].西北植物学报,2007,27(1):132-138.
[9]王斌,杨校生.4种典型地带性森林生态系统碳含量与碳密度比较[J].湖南农业大学学报:自然科学版,2010,36(4):464-469.
[10]梁佰华,潘秋荣,韦春梅.广东东源康禾省级自然保护区野生水果植物资源调查[J].亚热带植物科学,2012,41(3):56-60.
[11]林中大.广东康禾自然保护区植物区系研究[J].广西林业科学,2007,36(2):75-77+81.
[12]马克平,刘玉明.生物群落多样性的测度方法——Ⅰα多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[13]广东省林业局和广东省林业调查规划院.广东省森林资源调查常用数表[EB].广东省林业局,2009.
[14]徐新良,曹明奎,李克让.中国森林生态系统植被碳储量时空动态变化研究[J].地理科学进展,2007,26(6):1-10.
[15]CURTISR O,MARSHALLD D.Technical note:Why quadraticmean diameter?[J].Western Journal of Applied Forestry,2000,15(3):137 -139.
[16]MCCUNE B,MEFFORDM J.PC-ORD.Multivariate analysis of ecological data,Version 6.08[DB/CD].Gleneden Beach,Oregon,USA:MjM Software Design,2011.
[17]MCCUNE B,GRACE JB.Analysis of Ecological Communities[M].Gleneden Beach,Oregon,USA:MjM Software Design,2002:1-300.
[18]王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16.
[19]漆良华,彭镇华,张旭东,等.退化土地植被恢复群落物种多样性与生物量分配格局[J].生态学杂志,2007,26(11):1697-1702.
[20]张鹏超,张一平,杨国平,等.哀牢山亚热带常绿阔叶林乔木碳储量及固碳增量[J].生态学杂志,2010,29(6):1047-1053.
[21]HUSTONM A.Hidden treatments in ecologicalexperiments:re-evaluating the ecosystem function ofbiodiversity[J].Oecologia,1997,110(4):449 -460.
[22]LUYSSAERT S,SCHULZE E D,BORNER A,et al.Old-growth forests as global carbon sinks[J].Nature,2008,455(7210):213-215.
(责任编辑:吴显达)
Stand structure and biomass carbon density in a subtropical evergreen broadleaved forest of heterogeneous diversity
LIWen-bin1,HU Yan-qiu1,XU Ming-feng1,SU Zhi-yao1,CHENWei2,LYU Yuan-hui2
(1.College of Forestry,South China Agricultural University,Guangzhou,Guangdong 510642,China;
2.Forestry Administration of Dongyuan County,Heyuan,Guangdong 517000,China)
In the typical subtropical evergreen broadleaved forest area that originated from the same time in the Kanghe Provincial Nature Reserve of Dongyuan,Guangdong Province,three forest communitieswith heterogeneous diversity were identified by a route survey and an 1 hm2sample plot was set in each plant community separately to investigate the standing tree structure and carbon density distribution across the diversity gradient.The correlations of community species richness and the number of individualswith stand structure attributes in the sample plots exhibited various patterns.Species richness,the number of individuals increased as quadraticmean diameter and average DBH decreased,while the total DBH and basal area gradually increased.In Community 1 and Community 2,whose ecological dominance was lower and diversity index was higher as compared with Community 3,plant richness or number of individuals increased,which led to a higher stand volume,while such correlations in Community 3 were not the case.By comparing the standing tree structure and carbon density of these communities,itwas proved that community with lower diversity index had higher stand volume,biomass and carbon density in contrastwith the community with fewer dominant trees and higher diversity index.Trees with large DBH and ecologically dominant speciesmainly contributed to maintaining and increasing the stand volume and carbon storage in subtropical evergreen broadleaved forest.Our findings suggested that therewas an indirect relationship between standing tree structure index and stand carbon sequestration and high standing tree diversity did not lead to high community carbon sequestration.The critical role of treeswith large DBH and ecologically dominant species in the construction and conservation of forests targeted at carbon sequestration had to be emphasized.
standing tree structure;carbon density;diversity;stand volume
S718.54
A
1671-5470(2015)03-0256-08
10.13323/j.cnki.j.fafu(nat.sci.).2015.03.007
2014-07-26
2014-11-12
广东省林业科技创新专项(2011KJCX031-02)和2012年度国家级大学生科技创新项目资助.
李文斌(1990-),男,硕士研究生.研究方向:森林生态学.Email:liwenbeing@hotmail.com.通讯作者苏志尧(1963-),男,教授,博士生导师,研究方向:森林生态学.Email:zysu@scau.edu.cn.
