瓯江流域水生植物多样性与生态位研究
2015-03-01王金旺邹颖颖于
王金旺邹颖颖于 丹
(1. 浙江省农业科学院亚热带作物研究所, 温州 325005; 2. 温州市工业科学研究院, 温州 325000; 3. 武汉大学生命科学学院, 武汉 430079)
瓯江流域水生植物多样性与生态位研究
王金旺1邹颖颖2于 丹3
(1. 浙江省农业科学院亚热带作物研究所, 温州 325005; 2. 温州市工业科学研究院, 温州 325000; 3. 武汉大学生命科学学院, 武汉 430079)
根据2010—2013年调查结果, 结合相关文献资料, 研究了瓯江流域的水生植物多样性和物种生态位。结果表明: ①瓯江流域有水生高等植物181种(含亚种、变种), 隶属于41科78属, 属的分布型以世界分布为主, 占总属数 43.59%, 热带性质的属发育较温带性质充分。②30个临时样地划分了河流、沼泽、农田、沟渠、池塘、洪泛湿地等 6类生境类型, 单位面积物种数以洪泛湿地最高 0.09 ind./m2, 其次是沼泽生境0.07 ind./m2; 农田和池塘生境物种多样性随流域高程降低逐渐升高, 其余生境物种多样性以中游最高; 优势种生活型以挺水、漂浮植物为主, 沉水植物、浮叶植物重要值较低; ③挺水植物生态位宽度普遍高于其他生活型, 沉水、浮叶植物生态位宽度普遍较低; 相同生活型植物生态位重叠以漂浮植物间最高, 以沉水植物间最低; 不同生活型植物生态位重叠以挺水—浮叶植物间最高, 以沉水—漂浮植物间最低; 生态型多样的喜旱莲子草(Alternanthera philoxeroides)生态位宽度较大, 与其他物种生态位重叠高, 由此推测喜旱莲子草可能引起瓯江流域水生植物群落结构的变化, 导致乡土水生植物物种多样性的降低。
水生植物; 多样性; 生态位宽度; 生态位重叠; 瓯江流域
河流在人类社会和自然生态系统中发挥着重要作用, 是城市、农业发展的经济和文化载体。随着城市化进程和新农村建设的推进, 对河流利用和开发逐渐加大, 河流空间日渐缩小, 特别是小型自然溪沟被填埋、暗渠化或是原河流自然坡岸硬化, 造成河流面积下降[1]。与此同时, 与河流连通的湿地、湖泊等水体面积也逐渐减少, 如北京湿地资源自1996—2006十年间减少了55.14%[2]。湿地面积的降低影响了河流诸多功能的发挥, 如维持水文的缓冲区功能降低、维系生物多样性生境的丧失等均造成河流生态功能的退化。在河流生态系统研究中, 水生维管束植物具有重要的生态学价值, 水生植物是河流生态系统中重要的初级生产者, 为河流生态系统中其他水生生物提供食物来源和栖息地; 水生植物类群因其受光照[3]、水温[4]、干扰[5]、水体和沉积物理化性质[6]等环境因子, 以及其他生物类群如浮游植物、无脊椎动物、鱼类的影响[7, 8], 常被当作判断河流生态健康状态的指示生物类群[9]。因此, 河流生态系统水生植物多样性研究为掌握河流生态系统健康状况、生态功能提供了简单有效的方法, 也为科学管理河流生态系统提供理论依据。
每一个物种在生态系统中具有自己的生态位,生态位是客观存在的实体, 是分析评价物种之间内在关系及其种群在群落中地位的重要手段[10]。生态位研究在种间关系、群落结构、群落演替、生物多样性, 乃至物种进化中得到了广泛的应用[11—13]。
瓯江是浙江省第二大河流, 发源于浙闽边界百山祖洞宫山, 由西往东流经丽水、温州两市, 在温州湾流入东海, 干流全长 388 km, 流域面积 1.79× 104km2。瓯江干流自源头至丽水大港头称为龙泉溪,属上游; 大港头至青田石溪称为大溪, 属中游; 石溪以下至入海口称为瓯江, 属下游。瓯江支流自上而下有松阴溪、好溪、小溪和楠溪江。整个流域水生生态系统水体类型多样, 在流域上游, 溪流众多,高山湿地发育成熟, 孕育着大量溪源性湿地; 中游流域分布大量的浅水河滩、洪泛湿地、农田、池塘等类型水体; 下游流域河流纵横交错, 如温瑞塘河,入海口分布有滨海湿地。对瓯江流域水生植物的研究曾有一些零星报道[14, 15], 但缺乏全面系统的研究。本研究旨在从生态位角度研究瓯江流域水生植物多样性, 以期为瓯江流域为水生植物资源的有效保护、合理开发利用提供科学依据。
1 研究方法
1.1 研究区概况
瓯江流域位于东经 118°45′—121°00′、北纬27°28—28°48′。东临东海, 南与飞云江流域交界, 西与闽江流域接壤, 北部、西北部与钱塘江流域相邻。龙泉市凤阳山黄毛尖海拔1929 m, 是江浙第一高峰。流域属亚热带季风气候区, 温暖湿润, 四季分明, 雨水充沛, 年均温 18℃, 无霜期 250d以上, 多年平均降雨1746 mm。瓯江多年平均年净流量1.96×1010m3,水资源丰富, 水资源年际变化强烈, 降水季节变化极为明显[16]。
1.2 调查方法
由于在不同季节水生植物优势种类不同, 作者于2010年10月、2011年10月、2012年7月、2012 年10月、2013年5月和2013年10月先后6次对瓯江流域进行水生植物多样性调查和标本采集。调查时, 记录采样点经纬度, 绘制采样点分布图(图1)。调查的生境类型包括天然湿地和人工湿地, 参照《湿地公约》关于湿地类型的分类系统对其进行划分,对天然湿地中“永久性的河流”、“时令河”、“泛滥地”,及各类沼泽进行了适当的调整, 对人工湿地中的“灌溉地”进行了细化, 具体包括以下 6类: ①沼泽(这里指自然形成的分布在山体的积水坑、沼泽化草甸、地下水漏点周围形成的湿地), ②河流(指瓯江流域永久性河流、支流, 及其间歇性的分支溪流、小河), ③洪泛湿地(指季节性、间歇性河水泛滥淹没的河滩), ④池塘(指农用水塘、蓄水池、养殖池塘),⑤农田(指以种植水稻为主的水田), ⑥沟渠(指人工开掘的灌溉渠系)。调查的水生维管束植物指广义水生植物, 包括永久沉没在水中、或者漂浮于水面或光合作用部分漂浮于水面而根系生长于泥中的真性水生植物和生长于含饱和水的土壤湿生植物。所采集的水生植物标本于实验室进行鉴定, 存疑类群请专家鉴定(部分标本保存于浙江省亚热带作物研究所标本室)。此外, 查阅了浙江省内主要标本馆(杭州植物园标本馆、浙江省自然博物馆标本馆、浙江农林大学标本馆、温州大学标本馆)中记载的瓯江流域水生植物。
1.3 生态位研究方法
2013年5月和10月先后两次采用设置临时样地的方法对瓯江流域水生植物进行了调查, 共设置了30个长20 m、宽10 m的临时样地, 其中上游、中游、下游各10个样地(图 1)。调查时记录样地地标, 踏查记录样地内的所有水生植物种类, 采集标本, 记录盖度。
因某些水生植物如沉水植物株计数比较困难,有些水生植物资源量较少, 在不破坏资源的前提下,采用相对盖度(水生植物群落中某一物种分盖度占所有物种分盖度之和的比值)和相对频度(水生植物临时样方中某一物种出现的频率占所有物种出现频率之和的比值)来确定其在群落中的重要值, 同时计算群落物种多样性指数。

式中, IV表示重要值, RF表示相对频度, RC表示相对盖度,iP为物种i的相对重要值, S为物种数目。
生态位宽度 (1) Levins生态位宽度

式中, Bi是种i的生态位宽度, Pij是种i对第j个资源的利用占它对全部资源利用的频度, 即 nij为种i在资源j上的优势度(本文即样方中物种的重要值), r为资源等级数[17]。



(2) Hurlbert生态位宽度其中, , Ba为生态位宽度, Pij和r的含义同上式, 该方程的值域为[0, 1][18]。
生态位重叠 生态位重叠是指在一定资源序列上, 两个物种利用相同等级资源时相互重叠程度, 其计算公式为[19]:

式中, D表示物种i与物种 h的生态位重叠, 具有域值[0, 1], Pij、Phj分别为两个物种对资源位j的占有率(该文以物种重要值百分率来表示)。
2 结果
2.1 瓯江流域水生植物种属组成和属的分布区特点瓯江流域共有水生高等植物181种(含亚种、变种), 隶属于41科78属(附表1), 其中, 蕨类植物6 科6属6种、分别占总数的14.63%、7.69%和3.31%;单子叶植物15科44属97种, 分别占总数的36.6%、56.41%和53.59%; 双子叶植物20科28属78种, 分别占总数的48.78%、35.90%和43.09%。
根据吴征镒等[20, 21]的划分, 瓯江流域水生植物所有78属的分布区类型有10类(表1), 其中世界分布34属, 占总属数的43.59%, 热带性质属28个, 占总属数 35.90%, 温带性质属 16个, 占总属数的20.51%。瓯江流域水生植物属的分布型与我国相同分布型属数[20]比值表明: 瓯江流域水生植物总属数仅占我国总属数的 2.40%, 其中世界分布型属数占我国同分布型总属数的 34%, 说明瓯江流域水生植物的广域性; 热带性质属数占我国热带性质属数的比值为 1.77%, 高于温带性质属数占我国温带性质总属数的比值(1.21%), 此外, 瓯江流域热带性质属数与温带性质属数比值为 1.75, 表明瓯江流域水生植物热带性质的属发育较温带性质充分。
2.2 物种多样性与优势种
物种多样性 30个临时样地根据生境类型可划分六类, 其中河流6个、沼泽3个、农田8个、沟渠 5个、洪泛湿地 3个和池塘 5个, 各类生境中包含的广义水生植物种类分别为35种、44种、53种、35种、54种和31种, 单位面积物种数以洪泛湿地最高 0.09 ind./m2, 其次是沼泽0.07 ind./m2, 其余生境单位面积物种数相当(0.03—0.04 ind./m2)。

图1 水生植物野外调查采样点分布图(正方形标注表示临时样地调查点)Fig. 1 The distribution of sampling sites (suqares indicating the temporary sample plots)

表1 瓯江流域水生植物属的地理成分分析Tab. 1 The geographical elements of genera of aquatic plants in the Oujiang River
科属种的丰富度及Shannon-Wiener多样性指数在六类生境中差异显著, F科, 5, 24= 6.39, P = 0.01; F属, 5, 24= 3.84, P = 0.01; F种, 5, 24= 4.11, P = 0.01; FH, 5, 24= 10.02, P<0.01。河流和池塘的物种类群丰富度显著低于其他生境, 洪泛湿地和沼泽丰富度高(图2)。
在 30个临时样地中, 沼泽生境(3个)仅分布在瓯江上游, 洪泛湿地生境(3个)仅分布于中游, 其余四类生境在瓯江不同区段多样性指数变化较大(图3)。农田和池塘生境中物种多样性随流域高程的降低呈逐渐升高趋势(农田: H上游=2.34<H中游=2.63、H下游=2.63;池塘: H上游=1.47<H中游=1.59<H下游=2.63), 沟渠生境中物种多样性以中游为高且随流域高程的降低呈降低趋势(H中游=2.56>H上游=2.42>H下游=1.79), 河流生境中物种多样性以中游为高 (H中游=2.20>H上游=2.14>
H下游=1.73)。
物种的重要值及群落的优势种 30个临时样地中记录的水生植物种类有101种(含亚种、变种),占整个流域水生植物种类的 55.8%。河流生境中重要值为 0.01—0.19, 最高的为喜旱莲子草[Alteranthera philoxeroides (Mart.) Griseb.], 排列前9的物种重要值之和为61%, 隶属6科8属, 其中沉水、浮叶、挺水、漂浮生活型物种数比值为1∶1∶4∶3。沼泽生境中重要值为 0.01—0.10, 最高的为水蓼(Polygonum hydropiper Linn.), 排列前 9的物种重要值之和为48%, 隶属6科7属, 其中缺失沉水和浮叶生活型物种, 挺水和漂浮生活型物种比值为 7∶2。农田生境中重要值为 0.01—0.10, 最高的为喜旱莲子草, 排列前9的物种重要值之和为48%, 隶属7科9属, 与沼泽生境类似, 缺失沉水和浮叶生活型物种。沟渠生境中重要值为 0.01—0.18, 最高的为喜旱莲子草,排列前9的物种重要值之和为59%,隶属6科9属,缺失浮叶植物, 沉水、挺水、漂浮生活型物种比值为 3∶4∶2。洪泛湿地生境中重要值为 0.01—0.07,最高为喜旱莲子草, 排列前 9的物种重要值之和为45%, 隶属6科6属, 缺失沉水和浮叶植物, 挺水与漂浮生活型物种比值为8∶1。池塘生境中物种重要值0.01—0.18, 最高的为喜旱莲子草, 排列前9的物种重要值之和为57%, 隶属6科8属, 缺失沉水和浮叶植物, 挺水与漂浮生活型物种比值为6∶3。

图 2 不同生境物种分类群数量(小写字母表示科间差异、大写字母表示属间差异, 右上角带’的大写字母表示种间差异)及Shannon-Wiener多样性指数比较(小写字母表示)Fig. 2 Comparison of the numbers of taxonomic groups (Lowercase letters indicates the differences between families; Capital letters indicated the differences between genera; Capital letters at the upper right corner with “’” indicated the differences between species) and Shannon-Wiener index among different habitats (lowercase letters)

图 3 瓯江流域不同区段样点物种多样性指数比较(空心图标代表下游样方、黑色实心图标代表上游样方, 灰色实心图标代表中游样方)Fig. 3 Shannon-Wiener index of different regions in the Oujiang River(Hollow icons indicated the downstream sampling plots, Black solid icons indicated the upper sampling plots, Gray solid icons indicated the middle sampling plots)
由图4可看出, 6类生境中河流、沟渠和池塘生境中重要排列前9的物种重要值之和高于50%, 沼泽、农田、洪泛湿地生境中重要值排列前9的物种重要值之和低于 50%。6类生境中重要值排列前 9的物种共有27种, 其中在各类生境中仅出现1次的物种有15种, 占55.56%(外来种与乡土种比值为0),出现2次的物种有5种, 占18.52%(外来种与乡土种比值为 0), 出现 3次及以上的物种有 7种(凤眼莲Eichhornia crassipes (Mart.) Solms、水芹Oenanthe javanica (Bl.) DC.、丁香蓼 Ludwigia epilobiloides Maxim.、菰Zizania latifolia (Griseb.) Turcz. ex Stapf、秕壳稻Leersia sayanuka Ohwi、水蓼、喜旱莲子草), 占25.93% (外来种与乡土种比值为0.4)。各类生境中重要值较高的物种以挺水和漂浮生活型植物为主, 沉水和浮叶植物重要值低; 相比其他生境, 沟渠、河流生境中沉水和浮叶植物重要值稍高, 即使在沉水和浮叶植物重要值相对较高的沟渠生境中,重要值排列前 9的物种中沉水植物重要值之和仅为13%。

图4 不同生境物种重要值(a. 河流, b. 沼泽, c. 农田, d. 沟渠, e. 洪泛湿地, f. 池塘)Fig. 4 Important value (IV) of species in different habitats (a. river; b. swamp; c. farmland; d. ditch; e. floodpain; f. pond)
2.3 生态位宽度与生态位重叠
6类生境中的优势种(这里以各类生境中重要值最高的前5个物种来计), 以及每类生境中重要值最高的前3个真性水生植物(这里仅指沉水或浮叶类型,不包括漂浮植物)的生态位宽度的计算结果表明(表2): 两种方法计算的生态位宽度基本一致; 生态位宽度为0的物种(在30个样地中仅在一类生境中出现)为6种, 占23%, 如谷精草科(Eriocaulaceae)和茨藻科(Najadaceae)仅在沼泽中发现, 川蔓藻(Ruppia maritima Linn.)和角果藻(Zannichellia palustris Linn.)生长于咸水生境, 仅在河口沟渠生境中发现。挺水类群普遍高于其他类群, 如两种方法计算的生态位宽度最高的前5个物种中挺水植物占80%, 分别为丁香蓼、秕壳稻、水蓼、水芹, Bi和Ba分别为0.75 和0.88、0.75和0.87、0.74和0.80, 其次是生态型多样的喜旱莲子草, Bi和Ba分别为0.72和0.77; 沉水植物生态位宽度普遍较低, 以菹草(Potamogeton crispus Linn.)和轮叶黑藻[Hydrilla verticillata (Linn. f.) Royle]稍高, 为瓯江流域沉水类型的优势植物,主要分布在河流、沼泽、沟渠、农田和洪泛湿地, 在调查的池塘样地中未见。
挺水类群中以禾本科种类最多, 占该类群总数的 36%, 其中最高的是秕壳稻, 其次是菰; 沉水类群以眼子菜科和水鳖科为主, 占该类群种数的83%,其中最高的分别是菹草和轮叶黑藻; 漂浮和浮叶类群种类分散, 各科中仅1种植物。
喜旱莲子草与另外4种生态位宽度大的物种[丁香蓼、秕壳稻、柳叶箬 Isachne globosa (Thunb.) Kuntze、水芹]重叠值分别为0.84、0.80、0.60和0.94; 4种生态位宽度大的挺水植物生态位重叠值亦较大,介于 0.61—0.84, 生态位最宽的沉水植物轮叶黑藻与其他物种的生态位重叠较之其他沉水植物高, 表明生态位宽度大的物种, 生态位相似性大, 生态位重叠程度高(表3)。生态位宽度为0的物种, 如谷精草(Eriocaulon buergerianum Korn.)、南投谷精草(E. nanrtoense Hayata)、川蔓藻、角果藻、纤细茨藻[Najasgrancillima (A. Br.) Magnus]、黄花狸藻(U. aurea Lour.), 这些物种生态位重叠非 0即 1, 起因在于这些物种仅在一类生境中被发现; 这些物种与其他物种生态位重叠亦较低, 如川蔓藻与芦苇[Phragmites australis (Cav.) Trin. ex Steud.]的生态位重叠为0.66,与其他物种生态位重叠低于 0.50; 谷精草与有尾水筛[Blyxa echinosperma (Clarke) Hook. f.]的生态位重叠为 0.82, 与其他物种生态位重叠低于 0.45; 黄花狸藻与大漂(Pistia stratiotes Linn.)的生态位重叠为0.52, 与其他物种生态位重叠低于 0.45, 表明生态位宽度窄的物种生态位重叠程度亦低。

表2 不同生活型优势物种生态位宽度Tab. 2 The niche breadth of the dominant species in different life forms
相同或不同生活型物种之间的生态位重叠数据表明, 相同生活型植物中以漂浮植物生态位重叠最高0.24—0.78, 以沉水植物最低0—0.82, 生态位重叠均值排序为漂浮生活型(0.50)>浮叶生活型(0.46)> 挺水生活型(0.39)>沉水生活型(0.26); 不同生活型植物生态位重叠以“挺水-浮叶”最高, 以“沉水-漂浮”最低, 生态位重叠均值排序为“挺水-浮叶” (0.44)>“漂浮-浮叶”(0.40)>“挺水-漂浮” (0.34)>“沉水-浮叶”(0.32)>“挺水-沉水”(0.31)>“沉水-漂浮”(0.16), 可见沉水植物与其他生活型植物生态位重叠较低。
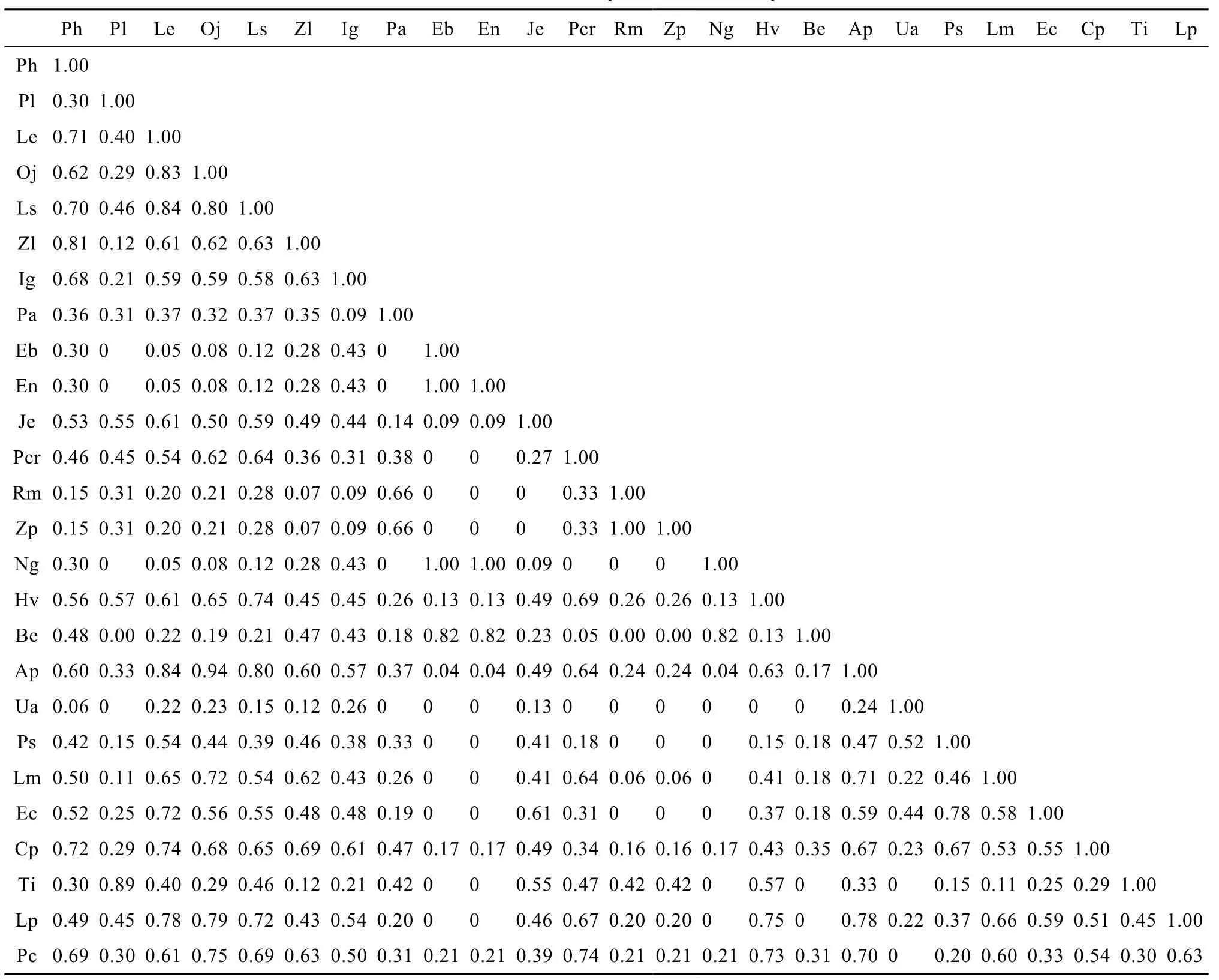
表3 优势种生态位重叠统一用英文简称Tab. 3 The niche overlap of the dominant species
3 讨论
3.1 气候地带性对瓯江流域水生植物多样性的影响
水生生境相对稳定, 通常水生植物多以世界分布为主, 但是气候地带性仍是一些对温度敏感类群分布的主要限制因子[22]。瓯江流域地处中亚热带南端与南亚热带北缘的交汇处, 光热充分, 一些热带性质的属种可从亚洲热带经华南延伸至瓯江流域:如水蕹(Aponogeton lakhonensis A. Camus)见于龙泉,水筛属植物(Blyxa spp.)多见于庆元、龙泉、松阳、遂昌、瓯海、云和等地, 稗荩[Sphaerocaryum malaccense (Trin.) Pilger]见于瓯海、龙泉等地, 鳞籽莎(Lepidosperma chinense Nees et Meyen)见于瓯海等。值得注意的是, 国内最早曾于2001年在海南省发现凤眼莲实生苗[23], 2012年笔者在庆元采集到凤眼莲实生苗。另外, 一些温带性质的属种从我国东北、华北分布向南延伸分布于瓯江流域: 如曲轴黑三棱(Sparganium fallax Graebn.)分布于永嘉、缙云, 慈姑属植物(Sagittaria spp.)在山区有较多分布等。相比温带性质的属种, 热带性质属种在瓯江流域内分布更广。因此,瓯江流域水生植物表现出较高的热带性质成分。
3.2 瓯江流域不同生境中水生植物多样性
物种多样性格局是群落演替动态过程的体现,反映了植物与环境的动态关系, 是生境因子对植物生长影响的反应结果[24]。在调查的6类生境样地中,洪泛湿地单位面积物种数最多, 多样性最高, 可能原因在于大量堰坝的修建形成的河流梯级开发改变了自然演替过程。据《瓯江志》记载, 自1949至1990 年, 流域内建成大中型水库16座、小型水库49座, 各类堰坝 1.7万余条[16]。水坝下游因上游来水量受限,形成大量洪泛湿地, 在洪泛区形成诸多与下游河道保持连通的回水湾, 这些回水湾静水生境为水生植物提供了避难所, 加上季节性洪水消涨, 大量陆源营养的输入为水生植物的生长提供了良好生境[25]。但由于缺乏可靠的水生植物历史调查资料, 无从比较瓯江梯级开发前后因流域环境的改变而造成的水生植物多样性的变化。沼泽生境因其分布于海拔600 m以上山区, 一些温带性质属种向南延伸也主要分布于高海拔沼泽生境[22], 丰富了沼泽生境物种多样性。
不同生境斑块之间的连通性可在一定程度上调控生境斑块中物种多样性[26], 相互连通的生境斑块间物种传播方向性导致一些生境物种丰富度较低[27],通过比较发现瓯江流域水生植物多样性随流域高程的升高呈降低趋势, 然而不同生境中的水生植物多样性随流域高程的变化的差异较大。在上游, 农田和池塘常见于山坳或小型盆地, 生境间彼此隔离;在中游, 瓯江流域主要盆地(碧湖-丽水盆地、壶镇盆地、西屏-古市盆地)集聚于此, 农田和池塘分布广,生境间彼此连通, 河流和沟渠成为水生植物传播的一种主要廊道; 在下游, 水网密度大, 但是河水污染严重[15], 河流中水生植物少, 农田和池塘则成为水生植物的避难所, 由此造成农田和池塘水生植物多样性随流域高程降低而升高, 河流和沟渠水生植物多样性则在中游为高, 上游高于下游的格局。
3.3 瓯江流域不同生境中水生植物优势种与生态位
6类生境中重要值排列前 9的物种共有 27种,在各类生境出现3次及以上的仅有7种, 其中喜旱莲子草和凤眼莲为外来植物, 其中喜旱莲子草生态位宽度不是最高, 但其重要值在除沼泽以外的生境中最高, 尤其是在河流、沟渠、池塘生境中其重要值高于 0.15, 为群落优势种; 凤眼莲生态位宽度<0.60, 但凤眼莲常形成单优漂浮植物群落。耐受-优势权衡(Tolerance-dominance trade-off)为喜旱莲子草和凤眼莲常成为群落优势种提供了解释[28], 因这两者常见于污染较重的生境中, 对污染的强耐受性促使其成为这类生境的优势种。与凤眼莲不同, 喜旱莲子草为瓯江流域不同生境的优势种还与其表型可塑性强有关, 表型可塑性常被认为是物种生境拓展、增加生态位宽度的有效方法[29, 30], 如枯水期喜旱莲子草以陆生型状态生长, 降雨后以湿生型或挺水型生长, 丰水期则以漂浮型状态生长, 不同生态型的转换使其有效拓展生境。一些物种通过器官特化(Specialization)拓展生境, 在调查中发现漂浮植物浮萍通过根系的螺旋状伸长可附生在岩壁上而不被水流冲走, 这种器官功能特化为其生态位扩展提供了途径。此外, 压力耐受性物种(Stress-tolerant species)生态位宽度较窄, 仅分布在特殊生境[28], 如圆叶挖耳草(U. striatula)、短梗挖耳草(U. caerulea)分布于山间阴湿的石壁上。
生态位重叠可以用于分析驱动物种分布的因子[19]。瓯江流域相同生活型中漂浮植物间生态位重叠最高,沉水植物间生态位重叠最低, 不同生活型物种间以“挺水-浮叶”植物间生态位重叠最高, “沉水-漂浮”植物间最低, 起因在于挺水和浮叶植物主要分布在沿岸带, 水深是其分布的主要影响因子, 有效光照则为沉水植物生长的主要限制因子[31, 32]。值得注意的是, 喜旱莲子草与其他物种生态位重叠普遍较高,入侵种对乡土植物群落的影响取决于其与乡土物种的生态位重叠程度, 生态位重叠高, 存在入侵种取代乡土植物的风险[33]。因此, 喜旱莲子草可能引起瓯江流域水生植物群落结构的变化, 导致乡土水生植物物种多样性的降低。
致谢:
本研究部分标本识别和鉴定得到了丁炳扬、熊先华的帮助, 在此表示感谢!
[1] Dudgeon D. Prospects for sustaining freshwater biodiversity in the 21stcentury: linking ecosystem structure and function [J]. Current Opinion in Environmental Sustainability, 2010, 2(5): 422—430
[2] Li L L, Gong H L, Zhao W J. Change detection and driving factor analysis of Beijing wetlands from 1996 to 2006 [J]. Journal of Capital Normal University (Natural Science Edition), 2008, 29(3): 95—102 [李玲玲, 宫辉力, 赵文吉. 1996—2006年北京实地面积变化信息提取与驱动因子分析.首都师范大学学报(自然科学版), 2008, 29(3): 95—102]
[3] Ray A M, Rebertus A J, Ray H L. Macrophyte succession in Minnesota beaver ponds [J]. Canadian Journal of Botany, 2001, 79(4): 487—499
[4] Unmuth J M L, Lillie R A, Dreikosen D S, et al. Influence of dense growth of Eurasian watermilfoil on lake water temperature and dissolved oxygen [J]. Journal of Freshwater Ecology, 2000, 15(4): 497—503
[5] Petticrew E L, Kalff J. Water flow and clay retention in submerged macrophytes beds [J]. Canadian Journal of Fisheries and Aquatic Science, 1992, 49(12): 2483—2489
[6] Lambert, S J, Davy A J. Water quality as a threat to aquatic plants: discriminating between the effects of nitrate, phosphate, boron and heavy metals on macrophytes [J]. New Phytologist, 2011, 189(4): 1051—1059
[7] Chick J H, McIvor C C. Patterns in the abundance and composition of fishes among beds of different macrophytes: viewing a littoral zone as a landscape [J]. Canadian Journal of Fisheries and Aquatic Science, 1994, 51(12): 2873—2882
[8] Mjelde M, Faafeng B A. Ceratophyllum demersum hampers phytoplankton development in some small Norwegian lakes over a wide range of phosphorus concentrations and geographical latitude [J]. Freshwater Biology, 1997, 37(2): 355—365
[9] Dawson H. Guidance for the field assessment of macrophytes of rivers within the STAR Project. 2002. http:// www.eu-star.at/frameset.htm
[10] Chen Y R, Yin L K. Community composition and niche change characteristics of dominant species in the windbreaking and sand-fixing forest, Xinjiang, China [J]. Journal of Plant Ecology (Chinese Version), 2008, 32(5): 1126—1133 [陈艳瑞, 尹林克.人工防风固沙林演替中群落组成和优势种群生态位变化特征.植物生态学报, 2008, 32(5): 1126—1133]
[11] Zhang J Y, Zhao H L, Zhang T H, et al. Niche dynamics of main populations of plants communities in the restoring succession process in Horqin Sandy Land [J]. Acta Ecologica Sinica, 2003, 23(12): 2741—2746 [张继义, 赵哈林, 张铜会, 等. 科尔沁沙地植物群落恢复演替系列种群生态位动态特征。生态学报, 2003, 23(12): 2741—2746]
[12] Ding B Y, Jin X F, Yu M J, et al. Impact to native species by invaded subaqueous plant Cabomba caroliniana [J]. Oceanologia et Limnologia Sinica, 2007, 38(4): 336—342 [丁炳扬, 金孝锋, 于明坚, 等.水盾草入侵对沉水植物群落物种多样性组成的影响.海洋与湖沼, 2007, 38(4): 336—342]
[13] Shen Z H, Fang J Y. Niche comparison of two Fagus species based on the topographic patterns of their populations [J]. Acta Phytoecologica Sinica, 2001, 25(4): 392—398 [沈泽昊,方精云.基于种群分布地形格局的两种水青冈生态位比较研究.植物生态学报, 2001, 25(4): 392—398]
[14] Zhu S C, Xu Y Y, Ye X G, et al. Preliminary analysis of the resources and floras of aquatic plants in Lishui City, Zhejiang Province [J]. Journal of Taizhou University, 2003, 25(6): 45—50 [朱圣潮, 徐燕云, 叶小郭, 等.丽水市水生维管束植物资源及区系的初步分析.台州学院学报, 2003, 25(6): 45—50]
[15] Hu R Y, Dai L P, Chen L. Studies on aquatic vascular plant resources of Wenzhou suberb [J]. Jianxi Science, 2002, 20(1): 47—51 [胡仁勇, 戴灵鹏, 陈靓. 温州市近郊水生维管束植物资源.江西科学, 2002, 20(1): 47—51]
[16] Oujiang Zhi Compilation Committee. Oujiang Zhi [M]. Beijing: China Water & Power Press. 1995, 45—61 [瓯江志编纂委员会.瓯江志.北京: 水利电力出版社. 1995, 45—61]
[17] Levins R. Evolution in Changing Environmens: Some Theoretical Explorations [M]. Princeton: Princeton University Press. 1968, 120
[18] Hurlbert S H. The measurement of niche overlap and some relatives [J]. Ecology, 1978, 59(1): 67—77
[19] Broennimann O, Fitzpatrick M C, Pearman P B, et al. Measuring eological niche overlap from occurrence and spatial environmental data [J]. Global Ecology and Biogeography, 2012, 21(4): 481—497
[20] Wu Z Y, Sun H, Zhou Z K, et al. Floristics of Seed Plants from China [M]. Beijing: Science Press. 2011, 120—319 [吴征镒, 孙航, 周浙昆, 等.中国种子植物区系地理.北京:科学出版社. 2011, 120—319]
[21] Wu Z Y. The areal-types of Chinese genera of seed plants [J]. Acta Botanica Yunnanica, 1991, Supplement IV: 1—139 [吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 增刊IV: 1—139]
[22] Ding B Y, Zhang Q M, Fang Y Y. The floristic features and the geographical distribution of aquatic vascular plants in Zhejiang Province [J]. Acta Hydrobiologica Sinca, 2001, 25(2): 128—136 [丁炳扬, 张庆勉, 方云亿. 浙江淡水维管束植物的区系特点与地理分布. 水生生物学报, 2001, 25(2): 128—136]
[23] Xie Y H, Yu D. Reproductive strategies, resource competition, ecological effects and control countermeasures of Water Hyacinth [A]. In: Xu R M, Ye W H (Eds.), Biological invasion-Theory and practice [C]. Beijing: Science Press. 2003, 186—188 [谢永宏, 于丹.凤眼莲的繁殖对策、资源竞争、生态影响和防治对策.见: 徐汝梅, 叶万辉, 生物入侵: 理论与实践.北京: 科学出版社. 2003, 186—188]
[24] Capers R S, Selsky R, Bugbee G J. The relative importance of local conditions and regional processes in structuring aquatic plant communities [J]. Freshwater Biology, 2010, 55(5): 952—966
[25] Yu D, Kang H, Chen Y Y. Lake bend effect and its influence on the specific diversity of aquatic plants in the lakes of middle basins of Changjiang River [J]. Acta Ecologica Sinica, 1996, 16(5): 476—483 [于丹, 康辉, 陈宜瑜.湖湾效应对长江中游湖泊水生植物多样性的影响.生态学报, 1996, 16(5): 476—483]
[26] Akasaka M, Takamura N. Hydrologic connection between ponds positively affects macrophyte α and γ diversity but negatively affects β diversity [J]. Ecology, 2012, 93(5): 967—973
[27] Altermatt F, Schreiber S, Holyoak M. Interactive effects of disturbance and dispersal directionality on species richness and composition in metacommunities [J]. Ecology, 2011, 92(4): 859—860
[28] Boulangeat I, Lavergne S, Es J V, et al. Niche breadth, rarity and ecological characteristics within a regional flora spanning large environmental gradients [J]. Journal of Biogeography, 2012, 39(1): 204—214
[29] Richards C L, Bossdorf O, Muth N Z, et al. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions [J]. Ecology Letters, 2006, 9(8): 981—993
[30] Nicotra A B, Atkin O K, Bonser S P, et al. Plant phenotypic plasticity in a changing climate [J]. Trends in Plant Science, 2010, 15(12): 684—692
[31] Chapin D M, Paige D K. Response of delta vegetation to water level changes in a regulated mountain lake, Washington State, USA [J]. Wetlands, 2013, 33(3): 431—444
[32] Wang J W, Yu D, Xiong W, et al. Above- and belowground competition between two submersed macrophytes [J]. Hydrobiologia, 2008, 607(1): 113—122
[33] Thum R, Lennon J T. Comparative ecological niche models predict the invasive spread of variable-leaf milfoil (Myriophyllum heterophyllum) and its potential impact on closely related native species [J]. Biological Invasions, 2010, 12(1): 133—143
附录: 瓯江流域水生植物名录
1. 水韭科 Isoetaceae
水韭属 Isoetes Linn.
东方水韭 Isoetes sinensis H. Liu et Q. F. Wang
2. 木贼科 Equisetaceae
木贼属 Equisetum Linn.
节节草 Equisetum ramosissima Desf.
3. 水蕨科 Ceratopteridaceae
水蕨属 Ceratopteris Brongn.
水蕨 Ceratopteris thalictroides (Linn.) Brongn.
4. 蘋科 Marsileaceae
蘋属 Marsilea Linn.
*蘋 Marsilea quadrifolia Linn.
5. 槐叶蘋科 Salviniaceae
槐叶 蘋属 Salvinia Adans.
槐叶 蘋 Salvinia natans (Linn.) All.
6. 满江红科 Azollaceae
满江红属 Azolla Lam.
*满江红 Azolla imbricata (Roxb.) Nakai
7. 三白草科 Saururaceae
(1) 蕺菜属 Houttuynia Thunb.
鱼腥草 Houttuynia cordata Thunb.
(2) 三白草属 Saururus Linn.
三白草 Saururus chinensis (Lour.) Baill.
8. 蓼科 Polygonaceae
(1) 蓼属 Polygonum Linn.
萹蓄 Polygonum aviculare Linn.
*火炭母 Polygonum chinense Linn.
*戟叶箭蓼 Polygonum hastato-sagittatum Mak.
*水蓼 Polygonum hydropiper Linn.
*蚕茧蓼 Polygonum japonicum Meisn.
*愉悦蓼 Polygonum jucundum Meisn.
*酸模叶蓼 Polygonum lapathifolium Linn.
绵毛酸模叶蓼 Polygonum lapathifolium var. salioifolium Sibth.
*长鬃蓼 Polygonum longisetum De Br.
圆基长鬃蓼 Polygonum longisetum De Br. var. rotundum A. J. Li
小花蓼 Polygonum muricatum Meisn.
红蓼 Polygonum orientale Linn.
春蓼 Polygonum persicaria Linn.
*暗果春蓼 Polygonum persicaria Linn. var. opacum (Sam.) A. J. Li丛枝蓼 Polygonum posumbu Buch.-Ham. ex D. Don
*疏花蓼 Polygonum praetermissum Hook. f.
伏毛蓼 Polygonum pubescens Blum.
*箭叶蓼 Polygonum sieboldii Meisn.
细叶蓼 Polygonum taquetii Levl.
*戟叶蓼 Polygonum thunbergii Sieb. et Zucc.
*粘毛蓼 Polygonum viscosum Hamilt.ex D. Don
(2) 酸模属 Rumex Linn.
*酸模 Rumex acetosa Linn.
齿果酸模 Rumex dentatus Linn.
*羊蹄 Rumex japonicus Houtt.
9. 苋科 Amaranthaceae
莲子草属 Alternanthera Forsk.
*喜旱莲子草 Alternanthera philoxeroides (Mart.) Griseb.*莲子草 Alternanrhera sessilis (Linn. ) DC.
10. 睡莲科 Nymphaeaceae
(1) 莼属 Brasenia Schreb.
莼菜 Brasenia schreberi J. F. Gmel.
(2) 芡属 Euryale Salisb. ex DC.
芡实 Euryale ferox Salisb. ex Koenig et Sims
(3) 萍蓬草属 Nuphar J. E. Smith
萍蓬草 Nuphar pumilum (Timm.) DC.
(4) 睡莲属 Nymphaea Linn.
睡莲 Nymphaea tetragona Georgi
11. 金鱼藻科 Ceratophyllaceae
金鱼藻属 Ceratophyllum Linn.
*金鱼藻 Ceratophyllum demersum Linn.
12. 毛茛科 Ranunculaceae
毛茛属 Ranunculus Linn.
茴茴蒜 Ranunculus chinensis Bunge
*石龙芮 Ranunculus sceleratus Linn.
扬子毛茛 Ranunculus sieboldii Miq.
13. 十字花科 Cruciferae
碎米荠属 Cardamine Linn.
弯曲碎米荠 Cardamine flexuosa With.
*碎米荠 Cardamine hirsuta Linn.
*水田碎米荠 Cardamine lyrata Bunge
14. 茅膏菜科 Droseraceae
茅膏菜属 Drosera Linn.
茅膏菜 Drosera peltata Smith ex Willdenow
圆叶茅膏菜 Drosera rotundifolia Linn.
匙叶茅膏菜 Drosera spathulata Labill.
15. 虎耳草科 Saxifragaceae
扯根菜属 Penthorum Linn.
扯根菜 Penthorum chinense Pursh
16. 水马齿科 Callitrichaceae
水马齿属 Callitriche Linn.
*沼生水马齿 Callitriche palustris Linn.
17. 千屈菜科 Lythraceae
(1) 水苋菜属 Ammannia Linn.
*水苋菜 Ammannia baccifera Linn.
多花水苋 Ammannia multiflora Roxb.
(2) 节节菜属 Rotala Linn.
*节节菜 Rotala indica (Willd.) Koehne
轮叶节节菜 Rotala mexicalla Cham. et Schlecht.
*圆叶节节菜 Rotala rotundifolia (Roxb.) Koehne
18. 菱科 Trapaceae
菱属 Trapa Linn.
*野菱 Trapa incisa Sieb. et Zucc.
欧菱 Trapa natans Linn.
19. 柳叶菜科 Onagraceae
丁香蓼属 Ludwigia Linn.
水龙 Ludwigia adscendens (Linn.) Hara
*丁香蓼 Ludwigia epilobiloides Maxim.
卵叶丁香蓼 Ludwigia ovalis Miq.
*黄花水龙 Ludwigia peloides (Kunth) P. H. Raven subsp. stipulacea (Ohwi) P. H. Raven
20. 小二仙草科 Haloragaceae
狐尾藻属 Myriophyllum Linn.
*穗花狐尾藻 Myriophyllum spicatum Linn.
*轮叶狐尾藻 Myriophyllum verticillatum Linn.
21. 伞形科 Apiaceae
水芹属 Oenanthe Linn.
短辐水芹 Oenanthe benghalensis Benth. et Hook. f.
*西南水芹 Oenanthe dielsii H. Boiss.
*水芹 Oenanthe javanica (Bl.) DC.
线叶水芹 Oenanthe linearis Wall. ex Cand.
22. 旋花科 Convolvulaceae
番薯属 Ipomoea Linn.
*空心菜 Ipomoea aquatica Forsk.
23. 玄参科 Scrophulariaceae
(1) 石龙尾属 Limnophila R. Br.
*石龙尾 Limnophila sessiliflora (Vahl) Bl.
(2) 母草属 Lindernia All.
泥花草 Lindernia antipoda (Linn.) Alston
*母草 Lindernia crustacea (Linn.) F. Muell.
狭叶母草 Lindernia micrantha D. Don
*陌上菜 Lindernia procumbens (Krock.) Philcox.
(3) 通泉草属 Mazus Lour.
通泉草 Mazus pumilus (N. L. Burman) Steenis
(4) 婆婆纳属 Veronica Linn.
水苦荬 Veronica undulata Wall.
24. 胡麻科 Pedaliaceae
茶菱属 Trapella Oliv.
*茶菱 Trapella sinensis Oliv.
25. 狸藻科 Lentibulariaceae
狸藻属 Utricularia Linn.
*黄花狸藻 Utricularia aurea Lour.
南方狸藻 Utricularia australis R. Br.
挖耳草 Utricularia bifida Linn.
短梗挖耳草 Utricularia caerulea Linn.
圆叶挖耳草 Utricularia striatula J. Smith
26. 爵床科 Acanthaceae
水蓑衣属 Hygrophila R. Br..
*水蓑衣 Hygrophila ringens (Linn.) R. Brown ex Sprengel
27. 香蒲科 Tyhaceae
香蒲属 Typha Linn.
*狭叶香蒲 Typha angustifolia Linn.
28. 黑三棱科 Sparganiaceae
黑三棱属 Sparganium Linn.
曲轴黑三棱 Sparganium fallax Graebn.
29. 眼子菜科 Potamogetonaceae
(1) 眼子菜属 Potamogeton Linn.
*菹草 Potamogeton crispus Linn.
*鸡冠眼子菜 Potamogeton cristatus Regel et Maack
微齿眼子菜 Potamogeton maackianus A.Benn Benn.
竹叶眼子菜 Potamogeton malaianus Miq.
尖叶眼子菜 potamogeton oxyphyllus Miq.
*小眼子菜 Potamogeton pusillus Linn.
(2) 川蔓藻属 Ruppia Linn.
*川蔓藻 Ruppia maritima Linn.
(3) 角果藻属 Zannichellia Linn.
*角果藻 Zannichellia palustris Linn.
30. 茨藻科 Najadaceae
茨藻属 Najas Linn.
草茨藻 Najas graminea Del.
*纤细茨藻 Najas grancillima (A. Br.) Magnus
31. 水蕹科 Aponogetonacaea
水蕹属 Aponogeton Linn. f.
*水蕹 Aponogeton lakhonensis A. Camus
32. 泽泻科 Alismataceae
(1) 泽泻属 Alisma Linn.
窄叶泽泻 Alisma canaliculatum A. Br. et Bouche
东方泽泻 Alisma orientale (Sam.) Juzep.
(2) 刺果泽泻属 Echinodorus Rich. et Engelm. ex A. Gray刺果泽泻 Echinodorus cordifolius (Linn.) Griseb
(3) 毛茛泽泻属 Ranalisma Stapf.
长喙毛茛泽泻 Ranalisma rostratum Stapf.
(4) 慈菇属 Sagittaria Linn.
小叶慈姑 Sagittaria potamogetifolia Merr.
*矮慈菇 Sagittaria pygmaea Miq.
*野慈菇 Sagittaria trifolia Linn.
33. 水鳖科 Hydrocharitaceae
(1) 水筛属 Blyxa Noronha ex Thouars
无尾水筛 Blyxa aubertii Rich.
*有尾水筛 Blyxa echinosperma (clarke) Hook. f.
*水筛 Blyxa japonica (Miq.) Maxim. ex Aschers et Gurke (2) 黑藻属 Hydrilla Rich.
*黑藻 Hydrilla verticillata (Linn. f.) Royle
(3) 水鳖属 Hydrocharis Linn.
*水鳖 Hydrocharis dubia (Bl.) Backer
(4) 水车前属 Ottelia Pers.
*水车前 Ottelia alismoildes (Linn.) Pers.
(5) 苦草属 Vallisneris Linn.
*苦草 Vallisneria natans (Lour.) Hara
刺苦草 Vallisneris spinulosa Yan.
34. 禾本科 Gramineae (Poaceae)
(1) 稗属 Echinochloa Beauv.
长芒稗 Echinochloa caudata Roshev.
*光头稗 Echinochloa colona (Linn.) Link
(2) 甜茅属 Glyceria R. Br.
甜茅 Glyceria acutiflora Torrey ssp. japonica (Steud.) T. Koyama et Kawano
(3) 柳叶箬属 Isachne R. Br.
*柳叶箬 Isachne globosa (Thunb.) Kuntze
矮小柳叶箬 Isachne pulchella Roth
平颖柳叶箬 Isachne truncata A. Camus
(4) 鸭嘴草属 Ischaemum Linn.
*毛鸭嘴草 Ischaemum anthephoroides (Steud.) Miq.
有芒鸭嘴草 Ischaemum aristatum Linn.
(5) 假稻属 Leersia Swartz
*秕壳稻Leersia sayanuka Ohwi
(6) 雀稗属 Paspalum Linn.
双穗雀稗 Paspalum distichum Linn.
(7) 芦苇属 Phragmites Adans.
*芦苇 Phragmites australis (Cav.) Trin. ex Steud.
(8) 米草属 Spartina Schreb.
互花米草 Spartina alterniflora Loisel.
(9) 稗荩属 Sphaerocaryum Nees ex Hook. f.
稗荩 Sphaerocaryum malaccense (Trin.) Pilger
(10) 菰属 Zizania Linn.
*菰 Zizania latifolia (Griseb.) Turcz. ex Stapf
35. 莎草科 Cyperaceae
(1) 球柱草属 Bulbostylis Kunth
球柱草 Bulbostylis barbata (Rottb.) Kunth
(2) 苔草属 Carex Linn.
*短尖苔草 Carex breviculpis C. B. Clark.
*十字苔草 Carex cruciata Wahlenb.
*垂穗苔草 Carex dimorpholepis stend.
弯囊苔草 Carex dispalata Boott
*穹隆苔草 Carex gibba Wahlenb.
长囊苔草 Carex harlandii Boott
*镜子苔草 Carex phacota Spreng.
粉被苔草 Carex pruinosa Boott
相仿苔草 Carex simulans C. B. Clark.
(3) 莎草属 Cyperus Linn.
异型莎草 Cyperus difformis Linn.
球形莎草 Cyperus glomeratus Linn.
畦畔莎草 Cyperus haspan Linn.
*碎米莎草 Cyperus iria Linn.
直穗莎草 Cyperus orthostachys Franch. et Sav.
*毛轴莎草 Cyperus pilosus Vahl
*香附子 Cyperus rotundus Linn.
(4) 荸荠属 Eleocharis R. Br.
*荸荠 Eleocharis dulcis (Burm.f.) Trin. ex Henschel
龙师草 Eleocharis tetraquetra Nees
*牛毛毡 Eleocharis yokoscensis (Franch. et Sav.) Tang et Wang
(5) 飘拂草属 Fimbristylis Vahl
*两歧飘拂草 Fimbristylis dichotoma (Linn.) Vahl
拟二叶飘拂草 Fimbristylis diphylliodes Makino
*日照飘拂草 Fimbristylis miliaceae (Linn.) Vahl
五棱杆飘拂草 Fimbristylis quinguangularis (vahl) Kunth
(6) 水蜈蚣属 Kyllinga Rottb.
*水蜈蚣 Kyllinga brevifolia Rottb.
(7) 鳞籽莎属 Lepidosperma Labill.
鳞籽莎 Nees et Meyen
(8) 藨草属 Scirpus Linn.
*萤蔺 Scirpus juncoides Roxb.
华东 藨草 Scirpus karuizawensis Makino
*海三棱 藨草 Scirpus × mariqueter Tang et F. T. Wang
类头状花序 藨草 Scirpus subcapitatus Thw.
*水毛花 Scirpus triangulatus Roxb.
*荆三棱 Scirpus yagara Ohwi
36. 天南星科 Araceae
(1) 菖蒲属 Acorus Linn.
*菖蒲 Acorus calamus Linn.
金钱蒲 Acorus gramineus Soland. ex Aiton
(2) 大 薸属 Pistia Linn.
* 大 薸 Pistia stratiotes Linn.
37. 浮萍科 Lemnaceae
(1) 兰氏萍属 Landoltia Les et D. J. Crawford
兰氏萍 Landoltia punctata (G. Meyer) Les et D. J. Crawford
(2) 浮萍属 Lemna Linn.
*浮萍 Lemna minor Linn.
稀脉萍 Lemna perpusilla Torr.
(3) 紫萍属 Spirodela Schleid.
*紫萍 Spirodela polyrhiza (Linn.) Schleid.
(4) 无根萍属 Wolffia Horkel ex Schleid.
无根萍 Wolffia arrhiza (Linn.) Wimmer
38. 谷精草科 Eriocaulaceae
谷精草属 Eriocaulon Linn.
*谷精草 Eriocaulon buergerianum Korn.
*白药谷精草 Eriocaulon cinereum R. Br.
江南谷精草 Eriocaulon faberi Ruhl.
*南投谷精草 Eriocaulon nanrtoense Hayata
*四国谷精草 Eriocaulon sikokianum Maxim.
39. 鸭跖草科 Commelinaceae
(1)鸭跖草属 Commelina Linn.
饭包草 Commelina benghalensis Linn.
*鸭跖草 Commelina communis Linn.
(2) 水竹叶属 Murdannia Royle
*疣草 Murdannia keisak (Hassk.) Hand.-Mazz.
*牛轭草 Murdannia loriforinis (Hassk. ) R. S. Rao et Kammathy*裸花水竹叶 Murdannia nudiflora (Linn.) Brenan
*水竹叶 Murdannia triquetra (Wall.) Bruckn.
40. 雨久花科 Pontederiaceae
(1) 凤眼莲属 Eichhornia Kunth.
*凤眼莲 Eichhornia crassipes (Mart.) Solms
(2) 鸭舌草属 Monochoria Presl.
*鸭舌草 Monochoria vaginalis (Burm. f.) Presl. ex Kunth
41. 灯心草科 Juncaceae
灯心草属 Juncus Linn.
*翅茎灯心草 Juncus alatus Franch. et Sav.
*星花灯心草 Juncus diastrophanthus Buch.
*灯心草 Juncus effusus Linn.
*江南灯心草 Juncus prismatocarpus R. Br.
野灯心草 Juncus setchuensis Buch.
标注*为临时样地出现的水生植物。
THE DIVERSITY, NICHE BREADTH, AND NICHE OVERLAP OF AQUATIC PLANTS IN THE OUJIANG RIVER
WANG Jin-Wang1, ZOU Ying-Ying2and YU Dan3
(1. Institute of Subtropical Crops, Zhejiang Academy of Agricultural Sciences, Wenzhou 325005, China; 2. Wenzhou Institute of Industrial Sciences, Wenzhou 325000, China; 3. College of Life Sciences, Wuhan University, Wuhan 430079, China)
The diversity and niche perspectives of aquatic plants are considered as key indicators of the ecological functions in a river ecosystem, and hence provide essential information for the management of rivers. Here we measured the diversity and niche of aquatic plants in different habitats in the Oujiang River, based on the field investigation from 2010 to 2013 and the previous reports. The results were shown below: (1) there were 181 species (including subspecies and varieties) that belonged to 78 genera and 41 families in the Oujiang River. Ten areal types were classified from these genera, in which the cosmopolitan accounted for the majority (43.59%), followed by the tropical and the temperate. The ratio of the tropical to the temperature was 1.75. Moreover, the relative ratio of the tropical genera to the same areal type nationwide (1.77%) was higher than that of the temperature (1.21%) in the Oujiang River. These suggested that tropical genera were further developed than the temperate type in the Oujiang River. (2) The 30 temporary plots were divided into six types of habitats, including river, swamp, farmland, ditch, pond, and floodpain. The species density was the highest (0.09 ind/m2) in the floodplains, followed by the swamps (0.07 ind/m2). The species diversity (Shannon-Wiener Index, H) gradually increased as the reaches became lower in the farmland and the pond (farmland: Hup=2.335 < Hmid=2.634, Hlow=2.63; pond: Hup=1.468 < Hmid=1.585 < Hlow=2.632), but in other habitats the species diversity was the highest in the middle reaches. The emergent and freely floating plants were usually the dominant aquatic species, whereas the values of importance of the submerged and floating-leaf plants were lower. (3) The niche breadth of the emergent plants was generally higher than that of other life forms of plants, and submerged and floating-leaf plants usually had low niche breadth. The niche overlap of the same life-form plants was the highest (0.5) between the freely floating plants, and was lowest (0.26) between the submerged plants. The niche overlap of the different life-form plants was the highest (0.44) between the emergent and the floating-leaf plants, and was the lowest (0.16) between the submerged and the freely floating plants. The value of importance of Alternanthera philoxeroides was the highest in all the surveyed plots except for the swamps. Moreover, the niche overlap between A. philoxeroides and other species was generally higher than that between other species in the same plots. Therefore the alien species A. philoxeroides may alter the community structure of aquatic plants, resulting in a lower species diversity of native aquatic plants.
Aquatic plants; Diversity; Niche breadth; Niche overlap; Oujiang River
Q145+.2
A
1000-3207(2015)06-1184-14
10.7541/2015.155
2014-09-15;
2015-03-25
国家自然科学青年基金“浙西南三大外流区水生植物多样性格局驱动力和压力研究(31300297)”; 温州市科技计划项目“瓯江流域水生植物资源现状及其潜在应用价值(S20090013)”; 温州水专项“温州河道水体水生植物与微生物生态修复模式技术研究与示范(S20140021)”资助
王金旺(1980—), 男, 湖北枣阳人; 博士; 主要从事水生植物学研究。E-mail: kingwwang@163.com
于丹(1957—), 男, 教授; 主要从事水生植物学研究。E-mail: yudan01@public.wh.hb.cn
