伯氏疏螺旋体膜蛋白OspC研究进展
2015-02-22汪佳婕宝福凯柳爱华
汪佳婕 宝福凯,2,3,4,5 柳爱华
1.昆明医科大学病原生物学与免疫学系 热带医学研究所,云南昆明 650500;2.昆明医科大学云南省高校热带传染病重点实验室,云南昆明 650500;3.云南省热带病示范型国际科技合作基地,云南昆明 650500;4.云南省公共卫生与疾病防控协同创新中心,云南昆明 650500;5.昆明医科大学生物化学与分子生物学系,云南昆明 650500
莱姆病(Lyme disease)作为一种蜱媒传播的传染性疾病, 在一些特殊人群和一些特别地区有很高的感染率。但到目前为止,缺少比较敏感和准确的诊断方法对莱姆病进行早期诊断。因此,要做到及时预防和及时治疗,在世界范围内还是个难题。经研究发现,莱姆病的致病病原体是一种叫做伯氏疏螺旋体(Borrelia Burgdorferi)的螺旋体,其外膜蛋白C(OspC)有非常强的抗原性和较强的免疫原性,机体在感染莱姆病的早期就可以产生特异性的IgM抗体,将这种抗体纯化后,对动物模型进行免疫,结果发现,动物模型在免疫后很短时间内就可检测到高浓度的IgM 抗体,目前,重组OspC 已用于莱姆病的临床诊断中。
1 OspC的细胞定位与化学结构
在进行OspC 的抗原性研究之前,首先对其细胞定位与化学结构做一定的研究,才有可能全面的研究其抗原性机制。
那么,外膜脂蛋白C(OspC)是莱姆病螺旋体的主要外膜蛋白之一,作为一种外膜蛋白,OspC 广泛存在于螺旋体细胞外膜中。从结构上来看,β-桶状结构是其共有的结构特征,不同的β-桶状结构由不同的偶数个β-折叠片组成,β-折叠的数量从8 ~ 22 个不等。在美国中西部、东北部等地区,发现25 种OspC 基因型的表达[1-2]。Elisabeth Baum 在其研究中指出,OspC 的179 氨基酸残基至188 个残基中的第五个C 末端折叠结构是OspC的特异性交互抗体结合的关键位置[3]。OspC 作为一个表面暴露的脂蛋白,可以引起早起的免疫反应[4-7]。在Elisabeth Baum 的研究中,得到了OspC的最大交互反应区域的3D 图形。如图1,在图中,OspC 二聚体的两条链分别被标以亮灰色与暗灰色。同时,在Elisabeth Baum 的研究中将OspC 二聚体中与抗体药物关联度最高的区域指出。其中,绿色部分代表与种OspC 有高关联度,黄色部分表示与两种OspC 蛋白有高关联度,红色部分表示与五种OspC 蛋白有高关联[8]。
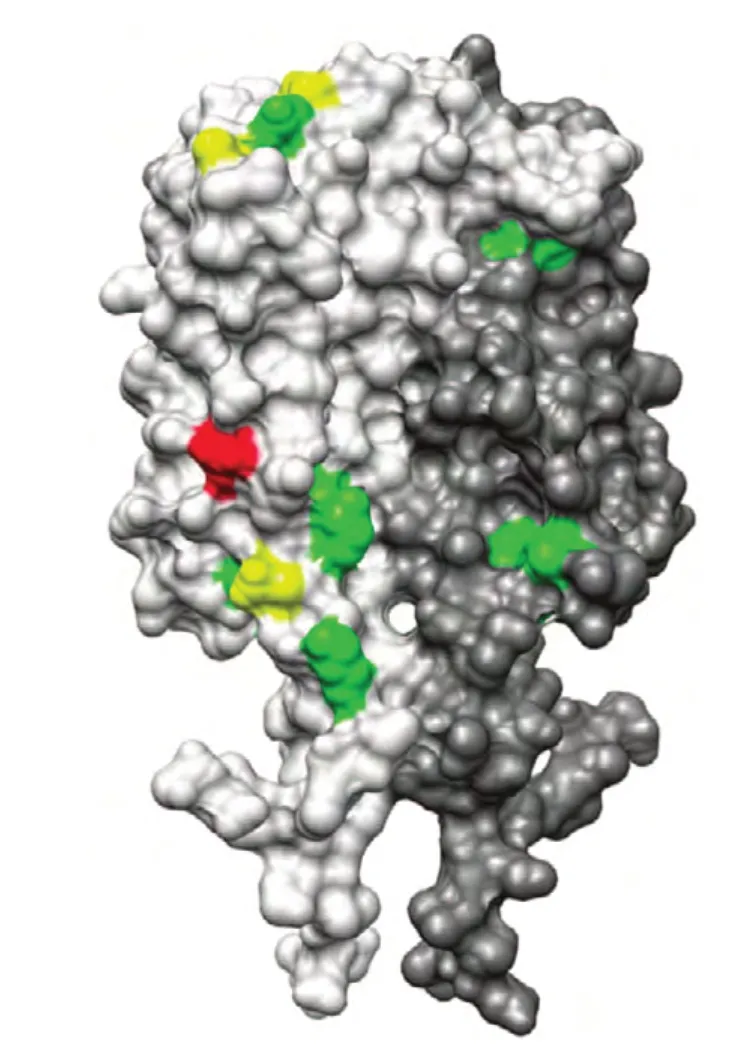
图1 OspC 的最大交互反应区域的3D 图形
2 OspC的表达调控
伯氏疏螺旋体特异性地调节其外膜脂蛋白OspC 的合成,以感染宿主细胞。为了适应哺乳动物体内的免疫环境,螺旋体首先利用OspC 在哺乳动物体内建立起其感染过程。伯氏疏螺旋体大量产生OspA 和OspB,而OspC 在硬蜱体内低表达,从蜱媒介传播到哺乳动物吸血后则出现上调[9],以帮助伯氏疏螺旋体转移到唾液腺[10],进而感染哺乳类动物[11-12]。感染一旦建立起来,OspC 的表达就会被抑制,以避免被宿主的免疫系统清除。同时伯氏疏螺旋体在人体内初期呈现一个高表达状态[13],当抗体产生后杀死了OspC 一直出现一个低表达克隆[14]。在表达基因的启动子中,反向重复序列的上游部分在调节OspC 的表达中起到关键作用。Dan Drecktrah 等[15]通过定点突变技术在分子水平上切割了OspC 表达基因操作子的末端反向重复序列,结果发现,打乱末端反向重复序列但保留邻近的反向重复序列能够阻止OspC 的合成。同时发现,温度,pH 以及DNA 超螺旋对于OspC 的合成也起到重要影响。Schwan 等[16]发现,在温度为32 ~ 37℃的条件下, OspC 的表达量大幅度地提高。由此说明,适宜的温度是OspC 大量表达的重要条件, 在不适宜的温度下,OspC 不表达或表达量很低。因此,近年来很多国外学者认为,pH 值、温度以及其他因素或许会对OspC 的免疫效果起到关键的影响[17-20]。更重要的是,比起核苷酸的序列,末端反向重复序列的碱基互补配对的存在对于控制OspC 的表达更为重要。Dan Drecktrah 的结果显示,顺式作用元件在OspC 毒力因子的表达上有关键的作用。
3 OspC致病性的研究
在对伯氏疏螺旋体外表面膜蛋白(OspC)致病性进行研究时发现,用OspC 与Salp15 结合,一起感染哺乳类动物,可以帮助伯氏疏螺旋体在局部定居,并抵抗免疫杀伤。2OspC 作为一个有效的免疫靶,表达下调时帮助伯氏疏螺旋体免疫逃逸,对OspC 在早期的感染中起重要作用[13]。虽然OspC是伯氏疏螺旋体关键的毒力因子之一,但是它在伯氏疏螺旋体的感染中的具体作用目前还不清楚。为了确定在接种了各种不同类型的OspC 菌株后,OspC 是否会引起宿主反应, Antonara 等分别接种了野生型OspC 菌株,突变型菌株以及完全突变型菌株后,对巨噬细胞,中性粒细胞以及细胞因子的产生进行了测定。在21 种细胞因子的测定中发现巨噬细胞趋化蛋白(MCP-1),角质细胞起源趋化因子(KC, CXCL1)以及血管内皮生长因子(VEGF)在伯氏疏螺旋体的接种位点有表达量上升的趋势。并且在接种一周内的多个时间点OspC 的表达量有不同的变化[21]。
4 OspC的诊断价值
到目前为止,缺少比较敏感和准确的诊断方法对莱姆病进行早期的诊断。因此,要做到及时诊断和治疗,在世界范围内还是个难题。经研究发现,莱姆病的致病病原体伯氏疏螺旋体外膜蛋白C(OspC)有较强的免疫原性,机体在感染莱姆病的早期就可以产生特异性的OspC IgM 抗体,将OspC 纯化后,对动物模型进行免疫,结果发现,动物模型在免疫后很短时间内就可以检测到高浓度的IgM 抗体。目前,重组OspC 已用于莱姆病的临床诊断。
近年的研究表明,OspC 作为伯氏疏螺旋体的主要外膜蛋白之一,具有很强的抗原性。Gilmore等[17]利用OspC 免疫12 只初生小鼠, 然后利用伯氏疏螺旋体菌株感染这12 只获得免疫的初生小鼠, 由此发现了OspC 能够使动物产生高浓度的抗体, 使其具有抵抗伯氏疏螺旋体感染的能力。
目前有不少研究报道指出,OspC 在宿主体内可以引起早期免疫反应,因此有研究者致力于利用OspC 作为抗原用来诊断早期的莱姆病[22-24]。早年,Hauser 等[25]培养了大量不同莱姆病的菌株,并从中提取了大量的OspC 作为抗原,利用ELASA技术检测了222 例莱姆病患者的血清和133 例对照供血者的血清。其结果显示,有几株菌株间的IgG 的检测灵敏度非常相似。利用Western blot 技术验证后发现,OspC 确实可以作为一种灵敏的莱姆病早期诊断抗原。在此之后,Rousselle 等[6]通过对比772 株狭义氏疏螺旋体的OspC 与14kD 鞭毛蛋白片段等其他细胞抗原平行应用于检测莱姆病病人和无症状的对照组的IgG 与IgM, 结果发现IgM 抗体的差异性最大。另外, Hauser 等[26]又做了进一步的研究,利用WB 技术检测了菌株PKo、PBi 与Pka2 的 灵 敏 度, 结 果 发 现OspC 对早期莱姆病感染的检出灵敏度要远远高于其他抗原,并最终推荐了OspC 作为PKo 诊断的最敏感的抗原。
Jobe 等[27]的研究中发现,在人类莱姆病的早期阶段中,ELISA 可以作为一种有效的方式来检测OspC 抗体,同时也能为成功治疗莱姆病作为一个重要的判断依据。
目前看来,OspC 作为伯氏疏螺旋体的主要抗原之一, OspC 虽然具有比较大的差异, 但其具有很强的特异性。因此,在诊断学临床应用中,将OspC作为诊断抗原,具有很大的临床优势。同时,如果将OspC 与其他的抗原相结合,进行配伍诊断,那么这种诊断方法不仅能够提高诊断的灵敏度, 还能减少与其他病原体的交叉反应。这将在将来莱姆病的诊断中显示出很强的优势与潜力。预计在今后的莱姆病防治中也会发挥一定的作用。至于在伯氏疏螺旋体的致病性上的研究, 目前的研究主要涉及于T 细胞的免疫反应、病原体本身的作用以及不同基因型螺旋体的作用。预计在以后的研究中,将会把OspC 与其他蛋白的研究相结合,从而获取更多的研究结果,在莱姆病的各方面获取进一步的进展。
付钰广等[28]利用来自SZ 菌株的重组蛋白OspC,进行WB 实验检测显示,重组的OspC 与阳性血清有较强的反应,同时SZ 菌株作为我国的流行菌株,若利用SZ 菌株的OspC 建立相对应的ELISA方法来检测伯氏疏螺旋体,这对我国莱姆病的预防和控制有很大作用,并有助于消除世界范围内莱姆病病患。同时,其研究的重组蛋白OspC(SZ 菌株)为建立羊体内的伯氏疏螺旋体抗体的ELISA 检测方法奠定了重要的基础。
目前在世界范围内对于OspC 的研究已经有了初步结果,对OspC 的研究有助于对莱姆病的发病原因,发病机理和发病规律进行了解。并且已经初步建立了利用OspC 作为检测抗原用来检测早期的莱姆病。但对于OspC 的深入研究还远远不够,还有待更深一步的实验与临床研究。
[1] Kramer VL,Beesley C.Temporal and spatial distribution of Ixodes pacificus and Dermacentor occidentalis(Acari:Ixodidae) and prevalence of Borrelia burgdorferi in Contra Costa County,California[J].J Med Entomol,1993,30(3):549-554.
[2] Barbour AG,Travinsky B.Evolution and distribution of the ospC Gene,a transferable serotype of Borrelia burgdorferi[J].MBio,2010,1(4)::e00153-10.
[3] Elisabeth Baum1,Arlo Z,Randall M,et al.Barbo.Inferring Epitopes of a Polymorphic Antigen Amidst Broadly Cross-Reactive Antibodies Using Protein Microarrays:A Study of OspC Proteins of Borrelia burgdorferi[J].PLoS ONE,2013,8(6):1-11.
[4] Kraiczy P,Hunfeld KP,Peters S,et al.Borreliacidal activity of early Lyme disease sera against complementresistant Borrelia afzelii FEM1 wild-type and an OspClacking FEM1 variant[J].J Med Microbiol,2000,49(10):917-928.
[5] Montgomery RR,Malawista SE,Feen KJ,et al.Direct demonstration of antigenic substitution of Borrelia burgdorferi ex vivo: exploration of the paradox of the early immune response to outer surface proteins A and C in Lyme disease[J].J Exp Med,1996,183(1):261-269.
[6] Rousselle JC,Callister SM,Schell RF,et al.Borreliacidal antibody production against outer surface protein C of Borrelia burgdorferi[J].J Infect Dis,1998,178(3):733-741.
[7] Jobe DA,Lovrich SD,Schell RF,et al.C-terminal region of outer surface protein C binds borreliacidal antibodies in sera from patients with Lyme disease[J].Clin Diagn Lab Immunol, 2003,10(4):573-578.
[8] Baum E,Randall AZ,Zeller M.,et al.Inferring epitopes of a polymorphic antigen amidst broadly cross-reactive antibodies using protein microarrays: a study of OspC proteins of Borrelia burgdorferi[J].PLoS One,2013,8(6):e67445.
[9] Eicken C,Sharma V,Klabunde T,et al.Crystal structure of Lyme disease antigen outer surface protein C from Borrelia burgdorferi[J].J Biol Chem,2001,276(13):10010-10015.
[10] Pal U,Yang XF,Chen M,et al.OspC facilitates Borrelia burgdorferi invasion of Ixodes scapularis salivary glands[J].J Clin Invest,2004,113(2):220-230.
[11] Ramamoorthi N,Narasimhan S,Pal U,et al.The Lyme disease agent exploits a tick protein to infect the mammalian host[J].Nature,2005,436(7050):573-577.
[12] Grimm D,Tilly K,Byram R,et al.Outer-surface protein C of the Lyme disease spirochete: a protein induced in ticks for infection of mammals[J].Proc Natl Acad Sci USA,2004,101(9):3142-3147.
[13] Seemanapalli SV,Xu Q,McShan K,et al.Outer Surface Protein C Is a Dissemination-Facilitating Factor of Borrelia burgdorferi during Mammalian Infection[J].PLoS ONE,2010,5(12):e15830.
[14] Xu Q,Seemanapalli SV,McShan K,et al.Constitutive expression of outer surface protein C diminishes the ability of Borrelia burgdorferi to evade specific humoral immunity[J].Infect Immun,2006,74(9): 5177-5184.
[15] Drecktrah D,Hall LS,Hoon-Hanks LL,et al.An inverted repeat in the ospC operator is required for induction in Borrelia burgdorferi[J].PLoS One,2013,8(7):e68799.
[16] Schwan TG,Piesman J,Golde WT,et al.Induction of an outer surface protein on Borrelia burgdorferi during tick feeding[J].Proc Natl Acad Sci USA,1995,92(7):2909-2913.
[17] Gilmore RD Jr,Mbow ML.Conformational nature of the Borrelia burgdorferi B31 outer surface protein C protective epitope[J].Infect Immun,1999,67(10):5463-5469.
[18] Wang G,van Dam AP,Dankert J.Evidence for frequent OspC gene transfer between Borrelia valaisiana sp.nov.and other Lyme disease spirochetes[J].FEMS Microbiol Lett,1999,177(2):289-296.
[19] Zhong W,Gern L,Stehle T,et al.Resolution of experimental and tick-borne Borrelia burgdorferi infection in mice by passive,but not active immunization using recombinant OspC[J].Eur J Immunol,1999,29(3):946-957.
[20] Wang IN,Dykhuizen DE,Qiu W,et al.Genetic diversity of ospC in a local population of Borrelia burgdorferi sensu stricto[J].Genetics,1999,151(1):15-30.
[21] Antonara S,Ristow L,McCarthy J,et al.Effect of Borrelia burgdorferi OspC at the site of inoculation in mouse skin[J].Infect Immun,2010,78(11):4723-4733.
[22] Wilske B,Habermann C,Fingerle V,et al.An improved recombinant IgG immunoblot for serodiagnosis of Lyme borreliosis[J].Med Microbiol Immunol,1999,188(3):139-144.
[23] Mathiesen MJ,Christiansen M,Hansen K,et al.Peptidebased OspC enzyme-linked immunosorbent assay for serodiagnosis of Lyme borreliosis[J].J Clin Microbiol,1998,36(12):3474-3479.
[24] Magnarelli LA,Flavell RA,Padula SJ,et al.Serologic diagnosis of canine and equine borreliosis:use of recombinant antigens in enzyme-linked immunosorbent assays[J].J Clin Microbiol,1997,35(1):169-173.
[25] Hauser U,Lehnert G,Wilske B.Validity of interpretation criteria for standardized Western blots (immunoblots) for serodiagnosis of Lyme borreliosis based on sera collected throughout Europe[J].J Clin Microbiol,1999,37(7):22411.
[26] Hauser U,Lehnert G,Wilsk E.B1Diagnost ic value of proteins of three Borrelia species(Borrelia burgdorferi sensu lato)and implications for development an d use of recombinant antigens for serodiagnosis of Lyme borreliosis in Europe[J].Clin Diagn Lab Immunol,1998,5(4):456-462.
[27] Jobe DA,Kowalski TJ,Bloemke M,et al.Rapid decline of OspC borreliacidal antibodies following treatment of patients with early Lyme disease[J].Clin Vaccine Immunol,2011,18(6):1034-1037.
[28] 付钰广,关贵全,牛庆丽,等.伯氏疏螺旋体外膜蛋白C 的表达及抗原性分析[J].中国兽医科学,2011,41(7):661-665.
