海马MF-CA3突触习得性LTP诱导后PPF效应的变化
2014-12-13严文文刘文晓黄俊柅
严文文,刘文晓,黄俊柅,肖 鹏
(华南师范大学生命科学学院,广州510631)
突触传递的长时程增强(Long-term potentiation,LTP)被认为是学习和记忆的神经基础[1]. 海马内环路有3个兴奋性单突触通路,即穿通纤维通路(Perforant pathway,PP)与DG 颗粒细胞的树突形成的突触(PP-DG),颗粒细胞发出的轴突组成的苔藓纤维(Mossy fibers,MF)与CA3 区锥体细胞的顶树突形成的突触(MF-CA3),锥体细胞的轴突从海马伞传出,其侧支(Schaffer collaterals)反折到CA1 区(Schaffer-CA1),与锥体细胞的顶树突形成的突触[2]. 此外,穿通纤维还存在不经DG 颗粒细胞直接到达CA3 区和CA1 区的投射. 海马离体脑片工作表明在以上通路上均可诱导产生强直性LTP[3].本课题组在直接结合行为学习的研究中表明,动物经行为学习后可在其海马各环路出现习得性LTP[4]. LTP 可分为2 种类型,即NMDA 受体依赖型和非NMDA 受体依赖型[5-6]. 大多数LTP 均为NMDA 受体依赖型,有关NMDA 受体依赖型强直性LTP 的表达机制目前仍有争论,有报导指出其表达过程涉及递质释放量增多[1],也有人认为其表达过程仅与突触后受体有关[7]. MF-CA3 突触的LTP 为非NMDA 受体依赖型,脑片研究表明,该突触通路的强直性LTP 在其诱导和维持过程中双脉冲易化(Paired-pulse facilitation,PPF)效应会下降[8]. PPF效应是指紧接着第1个检测刺激的第2个检测刺激引起突触后反应的增强[9]. 任何改变突触前递质释放率的因素都会改变PPF[10]. 这种PPF 效应的改变提示离体脑片MF-CA3 突触强直性LTP 的诱导及表达涉及突触前机制[11]. 然而,这种强直或离体脑片技术割裂了整体的整合作用,采用整体及结合行为学习的研究方法,对探讨学习记忆与突触传递效应的关系问题显得更为必要. 本文采用双脉冲检测技术,测定在体MF-CA3 突触习得性LTP 诱导前后及表达过程中PPF 效应的变化,对探讨该突触习得性LTP 与突触前机制的关系具有参考意义.
1 材料与方法
1.1 实验动物与分组
实验动物为Sprague Dawley 系雄性成年大鼠,体质量为180~220 g,由中山大学医学院动物实验中心提供,许可证号为SCXK 粤2004-0011. 动物随机分为不同间隔双脉冲检测组(n =8),训练组(n=8)和基线对照组(n =8). 不同间隔双脉冲检测组每实验日分别采用50、75、100、125、150 ms 间隔的双脉冲检测MF-CA3 突触的群体峰电位(Population spike,PS)峰值;训练组每实验日进行明暗辨别学习训练,在每实验日训练作业前及训练作业结束后采用间隔为150 ms 的双脉冲检测MF-CA3 突触PS 峰值;基线对照组不进行训练,每实验日只用间隔为150 ms 的双脉冲检测MF-CA3 突触的PS峰值.
1.2 电极埋植
实验前,所有动物均在无菌条件下进行慢性电极埋植手术.首先在腹腔注射戊巴比妥钠将动物麻醉(剂量为35 mg/kg).电极埋植手术过程在脑立体定位仪上进行. 记录电极为一枚绝缘不锈钢针(Ф 0.2 mm,尖端阻抗0.5~3.0 ΜΩ),尖端定位于海马CA3 区锥体细胞层,根据布瑞希图谱,定位为:AP 3.1~3.5;R(L)3.2~3.4;H 3.4~4.2. 刺激电极由两根绝缘的镍铬合金丝(Ф140 μm)绞合而成,尖端裸露0.2~0.3 mm,两尖端上下相距0.5~0.6 mm,埋植于海马DG 区的颗粒细胞层,定位为AP 3.1~3.5;R(L)1.9~2.1;H 3.5~4.1;参考电极为固定在颅骨上的小螺丝钉. 选择一能记录到突触反应的强度固定不变,然后逐步调节刺激电极和记录电极直至能记录到最大PS 反应的部位,用牙托粉小心封固,术后待动物康复3 天后进行实验.
1.3 行为训练模型
采用的学习模型为明暗辨别学习,在MD-IA 三等分辐射式迷宫(长45 cm ×宽14 cm ×高16 cm)中进行. 训练方法同前文[12-13],即每实验日同一时间进行20 次训练测试,每2 次测试间的时间间隔随机为20~40 s,随机给予不同灯臂电击,电击强度为0.45 mA,电击持续时间为1 s,电击延时5 s,以20次测试中90%的正确反应率为达到学会标准. 动物行为达学会标准后再巩固训练3 d.
1.4 数据采集与处理
检测以PS 峰值为评价指标. 检测刺激强度固定在训练前能引起1/3 至1/2 最大PS 效应的刺激强度,波宽为0.1 ms,双脉冲(即第一个脉冲与第二个脉冲)的间隔为从50、75、100、125、150 ms 中选取记录到稳定PPF 的脉冲间隔参数.检测刺激由刺激电极经SEN-7203 生理刺激器和MEZ-7101 隔离器输出,诱发电位由记录电极经SS-102J 微电极放大器,DSJ-F 生理放大器及A/D 卡三级放大输出,原始数据由“NEPAS 型生理信号分析处理系统”[14]在微机上进行采集和分析. PS 峰值增加30%以上表明出现习得性LTP[15]. PPF 易化率的计算为第二个脉冲反应的PS 峰值与第一个脉冲反应的PS 峰值之比(PSA2/PSA1,%).数据用平均值±标准差表示,实验结果采用非配对、双尾t 检验. 实验结束后使用普鲁士蓝法对刺激电极和记录电极定位进行检查,将定位不准确者剔除.
2 结果与分析
2.1 在MF-CA3 突触上检测到PPF 效应
当双脉冲间隔为50 ms 或75 ms 时(图1),在MF-CA3 突触上可检测到双脉冲抑制效应(Pairedpulse depression,PPD),双脉冲抑制率分别为43.87% ±14.74%和58.50% ±8.11%.当双脉冲间隔为100、125、150 ms 时,在MF-CA3 突触上检测到PPF 效应,PPF 易化率分别为109.74% ±36.31%、125.08% ± 37.12% 和139.37% ± 11.86%. 由于150 ms 双脉冲间隔检测到的PPF 效应最稳定,故选取该间隔的双脉冲检测训练组及基线对照组的PS峰值.连续7 天,在每天同一时间对动物进行双脉冲检测,结果表明,基线对照组在长达7 天的检测中,其MF-CA3 突触上第一个检测刺激引起的PS 峰值是稳定的(图2),表明本研究采用的双脉冲技术对MF-CA3 突触PS 峰值的检测未见累加效应. 即与以往的单脉冲检测的方法一样,可用第一个检测刺激引起的PS 峰值作为评价动物学习行为习得过程中突触效应的变化.
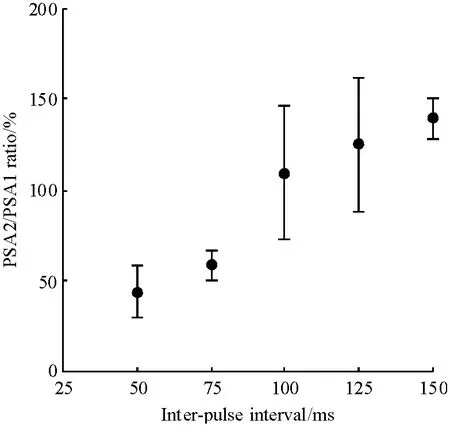
图1 不同间隔的双脉冲刺激检测到的双脉冲效应(n=8)Figure 1 Paired-pulse efficacy induced by paired-pulse stimulation of different inter-pulse interval (n=8)
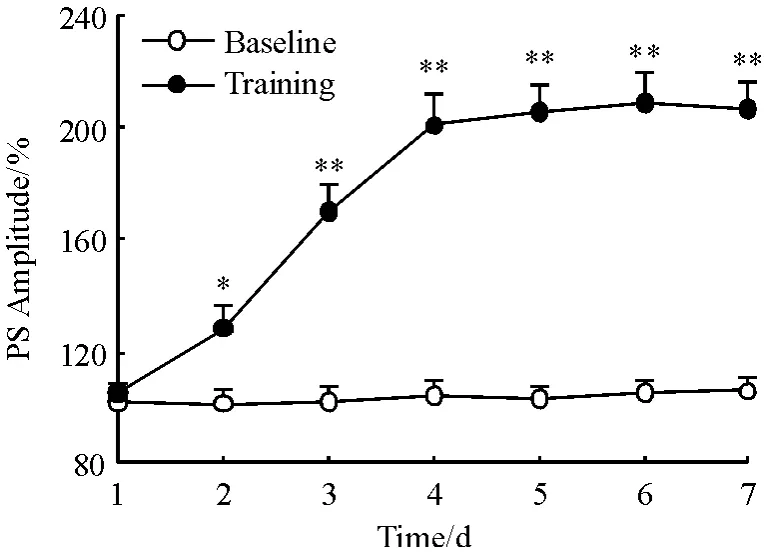
图2 双脉冲检测对PS 峰值的影响(n=8)Figure 2 Effect of paired-pulse test on PS amplitude(n=8)
2.2 明暗辨别学习后MF-CA3 突触出现习得性LTP
随着行为训练,训练组动物的正确反应率逐日提高(图3),在第4 训练日达到学会标准,其正确反应率为90.21% ±5.35%,经过3 天巩固训练,第7训练日的正确反应率为95.00% ±3.54%. MF-CA3突触的PS 峰值随着行为训练逐日增大,在达学会标准(第4 训练日)后PS 峰值为训练前基线水平的200.92% ±10.43%,经过3 天的巩固训练,PS 峰值也持续保持在最高水平,第7 训练日的PS 峰值为206.55% ±9.46%(图2). 而基线对照组在连续7天的检测中,PS 峰值则稳定在基线水平(100%)左右. 这表明动物经过明暗辨别学习后,随着行为的习得而产生了习得性LTP(图2).
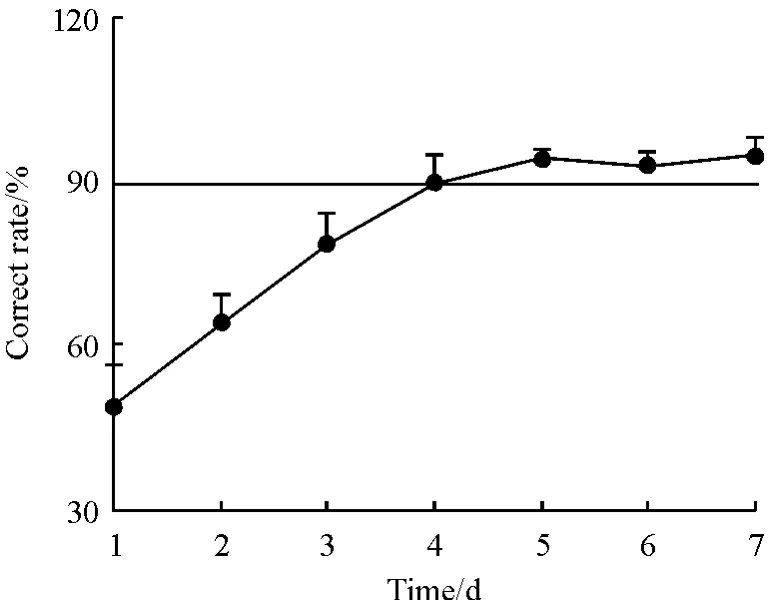
图3 明暗辨别学习后正确反应率的变化(n=8)Figure 3 Change of correct rate after behavioral learning(n=8)
2.3 MF-CA3 突触习得性LTP 形成及稳定后PPF易化率下降
结果表明,随着行为学习的进行,在习得性LTP形成后,PPF 由训练前平均的138.36% ±9.25%,降低到习得性LTP 形成后(第4 训练日)的114.75% ±8.42%,差异显著(P <0.01);在习得性LTP 稳定过程中PPF 持续维持在这个降低的水平.对照组的PPF 易化率在连续7 天的检测中均稳定在140%左右未见显著变化(图4 A、4B).
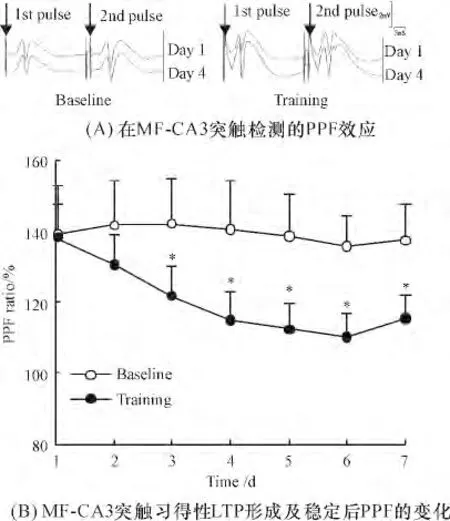
图4 MF-CA3 突触习得性LTP 形成及稳定后PPF 的变化(n=8)Figure 4 Change of PPF after learning-dependent LTP at MFCA3 synapses(n=8)
3 讨论
强直性LTP 的工作表明,海马内不仅存在经典的NMDA 受体依赖型LTP,还存在非NMDA 受体依赖型LTP[5-6]. 有关脑片NMDA 受体依赖型LTP 的诱导机制目前已基本阐明,认为高频刺激引起突触后膜产生持续的去极化,去极化一定程度导致常态下阻断NMDA 受体通道的Mg2+移开,NMDA 受体与突触前膜释放的谷氨酸递质结合而被激活[16],导致大量Ca2+内流,继而触发了一系列与Ca2+有关的生理生化反应,激活第二信使系统,改变膜的性质,触发了LTP 的产生[17]. 在PP-DG 突触上的习得性LTP 研究表明,NMDA 受体对习得性LTP 具有关键的启动作用,且与L 型电压依赖性钙通道存在相互关系或依赖性[4].有关NMDA 受体依赖型LTP 形成后的保持或稳定机制目前还存在着争论,主要表现为突触前学派和突触后学派之间的争论. 突触前学派认为其表达过程是谷氨酸递质释放量增多的结果[1];而突触后学派则认为其表达过程仅与突触后受体有关[7]. 在体PP-DG 突触上习得性LTP 的研究表明,DG 区习得性LTP 的表达与维持需突触前递质与突触后AMPA 受体的参与[4]. 在PP-CA1 突触上,PPF 易化率在习得性LTP 形成后较形成前有显著下降,提示在PP-CA1 通路上习得性LTP 的表达与突触前机制有关[18].
目前有关非NMDA 受体依赖型LTP 的研究比较少,该类型LTP 最早在海马MF-CA3 突触上发现[19]. 离体脑片研究表明,该突触通路的强直性LTP 的诱导和维持与突触前Ca2+的内流和谷氨酸递质释放的增加有关[11,20-22],且该过程中伴随着PPF 效应下降[8,23]. PPF 是由于第一个检测刺激后突触末端有部分残留的Ca2+,和第二次检测刺激引起的内流的Ca2+加合在一起,增强递质的释放,因而表现出第二次检测反应大的结果. 如果第一个脉冲所引起的递质释放率高,那么残留的Ca2+少,这样第二个脉冲所引起的递质释放就少了,即PPF 易化率小;相反,如果第一个脉冲刺激所引起的递质释放率低,那么残留的Ca2+增加,这样第二个脉冲刺激就会引起更多的递质释放,PPF 易化率就大[9].因而,任何改变递质释放率的因素都会改变PPF 易化率[10]. 由此可见,如果LTP 的表达与突触前递质释放无关,那么由突触前递质释放增多所引起的PPF 效应理应不受影响;相反,如果LTP 形成及表达涉及突触前递质释放的增加,那么由于递质的释放量在一定时间内是有限的,因而势必会影响PPF 效应[24]. 该分析已在实验工作中得到证明[7]. 因此,通过观察LTP 形成过程中PPF 的变化即可为研究该突触效应与突触前机制的关系提供参考. 本文采用双脉冲检测技术和慢性电极埋植技术,结合动物行为训练的方法,观察动物行为习得过程中在MFCA3 突触上习得性LTP 形成及表达过程中PPF 效应的变化. 当双脉冲间隔为50 或75 ms 时,MF-CA3突触出现双脉冲抑制效应;间隔为100 或125 ms时,双脉冲易化效应不稳定;而当双脉冲间隔为150 ms 时,可出现稳定的双脉冲易化效应,故选择150 ms 的双脉冲间隔检测MF-CA3 突触上习得性LTP形成及表达过程中PPF 效应的变化. 实验结果表明,随着行为训练的进行,训练组动物明暗辨别学习的正确反应率逐日提高,并在第4 训练日达到学会标准;MF-CA3 突触的PS 峰值逐日增大,在达到学会标准日达到最高水平,并随着行为习得的巩固保持在最高水平,提示行为习得后MF-CA3 突触形成习得性LTP. PPF 易化率在习得性LTP 形成过程中逐渐下降,在习得性LTP 形成后(第4 训练日)显著下降至最低水平,并在习得性LTP 的维持过程中保持在该水平,提示MF-CA3 突触习得性LTP 的形成及表达可能与突触前递质释放的改变有关,这对探讨该突触习得性LTP 的形成及表达与突触前机制的关系具有一定的参考意义. 然而,鉴于整体自然状态下的情况更为复杂,MF-CA3 突触习得性LTP的形成及表达机制仍有待进一步研究与探讨.
[1]Bliss T V,Collingridge G L. A synaptic model of memory:Long-term potentiation in the hippocampus[J]. Nature,1993,361(6407):31-39.
[2]Amaral D B,Witter M P. The three-dimensional organization of the hippocampal formation:A review of anatomical data[J]. Neuroscience,1989,31(3):571-591.
[3]Witter M P. Organization of the entorhinal hippocampal system:A review of current anatomical data[J]. Hippocampus,1993,3(1):33-44.
[4]肖鹏,许世彤,区英琦. 与学习过程相关的海马突触传递效能变化及其机制探讨[C]∥复旦大学生命科学学院.复旦神经生物学讲座.上海:复旦大学,2000:149-157.
[5]Reskel D J,Nicoll R A. Evidence for all-or-none regulation of neurotransmitter release:Implication for long-term potentiation[J]. The Journal of Physiology,1993,471(3):481-500.
[6]Aniksztejn L,Ben-Ari Y. Novel form of long-term potentiation produced by a K+channel blocker in the hippocampus[J]. Nature,1991,349(6304):67-69.
[7]Xiao P,Staubli U,Kessler M,et al. Selective effects of Aniracetam across receptor types and forms of synaptic facilitation in hippocampus[J]. Hippocampus,1991,1(4):373-380.
[8]Jaffe D,Johnston D. Induction of long-term potentiation at hippocampal mossy-fiber synapses follows a Hebbian rule[J]. Journal of Neurophysiology,1990,64(3):948-960.
[9]Salin P A,Scanziani M,Malenka R C,et al. Distinct short-term plasticity at two excitatory synapses in the hippocampus[J]. Proceedings of the National Academy of Sciences of the United States of America,1996,93(23):13304-13309.
[10]Chu H Y,Wu Q,Zhou S,et al. SKF83959 suppresses excitatory synaptic transmission in rat hippocampus via a dopamine receptor-independent mechanism[J]. Journal of Neuroscience Research,2011,89(8):1259-1266.
[11]Zalutsky R A,Nicoll R A. Comparison of two forms of long-term potentiation in single hippocampal neurons[J].Science,1990,248(4963):1619-1624.
[12]Li C H,Wang S Z,Cai Z L,et al. Effects of intrahippocampal L-NAME treatment on the behavioral long-term potentiation in dentate gyrus[J]. Neuroscience Letters,2012,528(2):201-204.
[13]Yan W W,Wang C Y,Zeng J,et al. Low-frequency stimulation of dorsal norephinephrine bundle reverses behavioral long-term potentiation and learning performance in rats[J]. Neuroscience,2014,265:238-244.
[14]刘文晓,许世彤,区英琦. NEPAS-Ⅱ型诱发电位微机自动分析处理系统[J]. 生理学通报,1992,9(1/2):21-22.Liu W X,Xu S T,Ou Y Q. A microcomputer auto-analytic and processing system of NEPAS-Ⅱevoked potentials[J]. Sheng Li Xue Tong Bao,1992,9(1/2):21-22.
[15]Jones J P,Meck W H,Williams C L,et al. Choline availability to the developing rat fetus alters adult hippocampal long-term potentiation[J]. Developmental Brain Research,1999,118:159-167.
[16]Bashir Z I,Alford S,Davies S N,et al. Long-term potentiation of NMDA receptor-mediated synaptic transmission in the hippocampus[J]. Nature,1991,349(6305):156-158.
[17]Kauer J A,Malenka R C,Nicoll R A. NMDA application potentiates synaptic transmission in the hippocampus[J]. Nature,1988,334(6179):250-252.
[18]刘文晓,孙平,肖鹏. PP-CA1 突触通路的PPF 效应在习得性LTP 形成后的降低[J]. 生理通讯,2000,19(supply1):27-30.Liu W X,Sun P,Xiao P. The decrease of paired-pulse facilitation after learning-dependent LTP at PP-CA1 synapse in hippocampus[J]. Caps News Communication,2000,19(supply1):27-30.
[19]Alger B E,Teyler T J. Long-term potentiation and shortterm plasticity in the CA1,CA3 and dentate regions of the rat hippocampal slices[J]. Brain Research,1976,110(3):463-480.
[20]Castillo P E,Weisskopf M G,Nicoll R A. The role of Ca2+channels in hippocampal mossy fiber synaptic transmission and long-term potentiation[J]. Neuron,1994,12(2):261-269.
[21]Barnes S J,Opitz T,Merkens M,et al. Stable mossy fiber long-term potentiation requires calcium influx at the granule cell soma,protein synthesis,and microtubule-dependent axonal transport[J]. The Journal of Neuroscience,2010,30(39):12996-13004.
[22]Evstratova A,Tóth K. Information processing and synaptic plasticity at hippocampal mossy fiber terminals[J].Frontiers in Cellular Neuroscience,2014,doi:10.3389/fncel.2014.00028.
[23]Son H,Carpenter D O. Interactions among paired-pulse facilitation and post-tetanic and long-term potentiation in the mossy fiber-CA3 pathway in rat hippocampus[J].Synapse,1996,23(4):302-331.
[24]Staubli U,Kessler M,Lynch G. Aniracetam has proportionately smaller effects on synapses expressing long-term potentiation:Evidence that receptor changes subserve LTP[J]. Psyco-biology,1990,18:377-381.
