断鸣管神经支对白腰文鸟鸣声的影响
2014-12-13林桂霞郑志强王松华王怡淳李东风
林桂霞,郑志强,吕 源,王松华,王怡淳,吴 敏,李东风
(华南师范大学生命科学学院,广州510631)
鸟类是自然界中具有复杂发声行为的动物. 对鸟类发声行为的研究己经成为探讨语言学习记忆神经机制的热点[1].
鸣管是鸟类特有的发声器官,位于气管和支气管的交界处.支配鸣肌收缩的神经支为气管鸣管支(NXIIts),它是舌下神经(NXII)自延髓发出后的一条分支.鸟类的发声是在发声中枢控制下由NXIIts支配鸣肌收缩而产生的[2].
白腰文鸟(White-rumped Munia,Lonchura striata),又称十姐妹,为燕雀目金腹科鸣禽.其声音信号具有高度的重复性和规律性,是研究鸣声学习理想的模型动物.目前对其发声行为的神经机制、发声行为的性别差异等已有相关的报道[3-7]. 但目前尚无断鸣管神经对白腰文鸟叫声性别差异的报道.
本实验采用声谱分析的方法,对断鸣管神经前后的白腰文鸟鸣声进行声学特征的比较和分析,进一步探讨鸟类发声器官神经支配的侧别优势及雄雌差异,为进一步了解鸟类发声机制提供新的认识.
1 材料与方法
实验用有效成年白腰文鸟12 只,体质量10~13 g,购于广州花地湾市场,置于华南师范大学生命科学学院动物房饲养.自然采光,室温25℃,供给饲料和水.实验动物使用符合国家和学校动物伦理委员会规定并遵守国际惯例.
实验动物饲养一周后便可进行正常叫声的采集,采集叫声后分组进行实验. 雄鸟左右组各3 只(断神经先左后右3 只,先右后左3 只),雌鸟亦然.
用10%的水合氯醛轻度麻醉动物后,切开颈部皮肤,在气管两侧暴露出鸣管神经.剪断一侧鸣管神经后,缝合皮肤.3 d 后待叫声稳定,便可进行鸣叫声音的采集.然后断另一侧鸣管神经,同法再采集鸣叫声音.
录制的音频文件用Cool Edit Pro 软件进行整理和筛选,并用Wavesurfer 软件进行波形、语图等的采集和分析.制作波形图(waveform)、语图(sonogram)和功率频谱图(power spectrum). 用Photoshop 软件作图.再利用SAP 软件分析音节时长(Syllable duration,SD)、振幅(Amplititude,Amp)、基频(Fundamental frequency,FF)、主 频(Principal frequency,PF)、相对幅值(Relative amplititude,RA)、调频(Frequency modulation,FM)、调幅(Amplititude modulation,AM)和熵(Entropy). 数据进行组内单因素方差分析,用均值±标准误(mean ±SE)表示. 对手术前后的值进行t 检验分析.
实验结束,解剖检查性腺,确认性别.
2 结果与分析
2.1 白腰文鸟的正常鸣叫
2.1.1 雄鸟鸣叫与鸣唱 图1A 是单次鸣叫的波形图,可见单个脉冲即为单音节. 正常情况下,鸣叫时SD 为(724.8 ±12.50)ms. 图1B 是语图,横轴表示时间,纵轴表示频率,颜色的深浅表示能量的大小.能量主要集中在2~5 kHz 的频率范围,具有谐波.图1C 是功率频谱图,PF 为3 843 Hz.
成年雄鸟能够鸣唱.每首鸣曲由若干短语组成.图2 显示,鸣曲序幕音i 之后,由a、b、c 短语交替出现,每个短语又由几个不同音节组成.
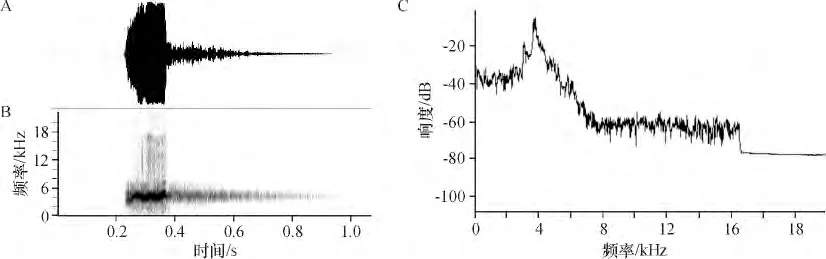
图1 成年雄性白腰文鸟正常鸣叫的波形图(A)、语图(B)、和功率频谱图(C)Figure 1 Waveform (A),sonogram (B)and power spectrum (C)of normal calls in adult male of White-rumped Munia

图2 成年雄性白腰文鸟的鸣曲波形图Figure 2 Normal song waveform in adult male White-rumped Munia
2.1.2 雌鸟鸣叫 成年雌鸟只会鸣叫.单次鸣叫声为单音节.能量主要集中在2~6 kHz,SD 为(378.59±43.38)ms.频率范围0~7 kHz 之间. PF 为3 907 Hz(图3).
2.1.3 两性白腰文鸟正常鸣叫的声学参数比较对各项声学参数进行比较后,发现叫声存在明显的性别差异.雄鸟的SD、Amp 和Entropy 都明显高于雌鸟,而RA、AM、FM 却明显低于雌鸟,PF 和FF 差异不大(图4).
2.2 断雄性白腰文鸟鸣管神经支对其鸣叫的影响
雄性白腰文鸟断左侧NXIIts 后,音量变小,音调变低. 2 kHz 以上的高频声音减少,尤其是7 kHz以上的高频声音几乎消失,而2 kHz 以下的低频声音则增多. 主频率部分对应的音量明显变小. SD、PF,Entropy 均下降,RA 增加.
断右侧NXIIts 后,多数鸣叫声学参数几乎没有发生变化. 只有Entropy 下降.
断双侧NXIIts 后,音量、音调和音色都严重受到影响.SD、PF、Entropy 下降,RA 增加(图5).
2.3 断雄性白腰文鸟鸣管神经支对其鸣唱的影响
与正常鸣唱相比(图6A),断左侧NXIIts 后,雄鸟鸣唱音量降低,部分音节消失,音色大减. 音节序列发生明显变化(图6B).断右侧NXIIts 后,除了脉冲形状稍有变化外,鸣曲的多数音节和短语都未受到较大影响(图6C).断双侧NXIIts 后,导致鸣曲音量变小、音调变低、音色变差(图6D).
2.4 断雌性白腰文鸟鸣管神经支对其鸣叫的影响
雌性白腰文鸟断左侧NXIIts 后,叫声音调变低.1~3 kH 之间的频率成分明显增多,而音节整体的频率范围基本不变. PF 下降,Entropy 增加. 断右侧NXIIts 后,音量、音调和音色几乎没有变化.PF 有所下降. 断双侧NXIIts 后,音量下降、音调降低. 4 kHz 以下的频率成分增多. SD、PF、Entropy 下降,RA 增加(图7).
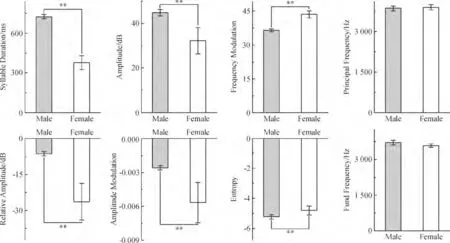
图4 雄雌白腰文鸟鸣叫声学参数对比Figure 4 Call comparison of acoustic analysis in male and female White-rumped Munia
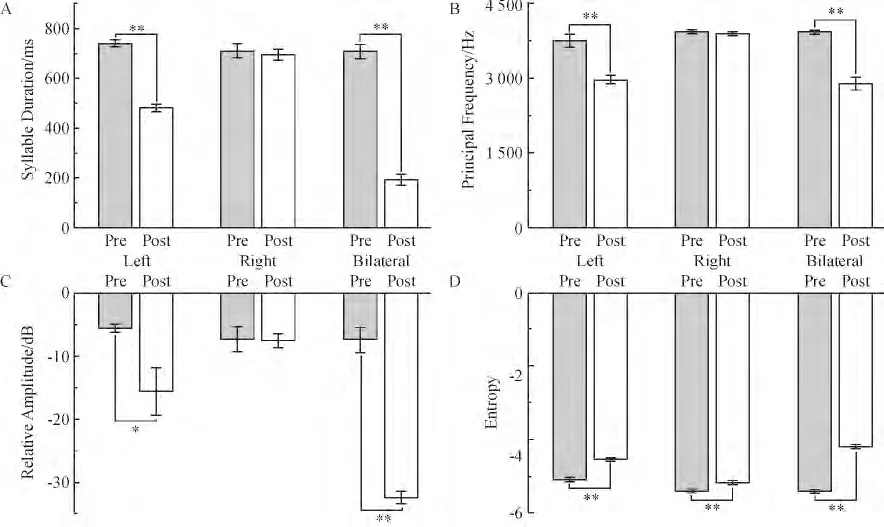
图5 雄鸟断神经前后各声学参数的变化Figure 5 Comparison of acoustic analysis before and after section of NXIIts in male White-rumped Munia
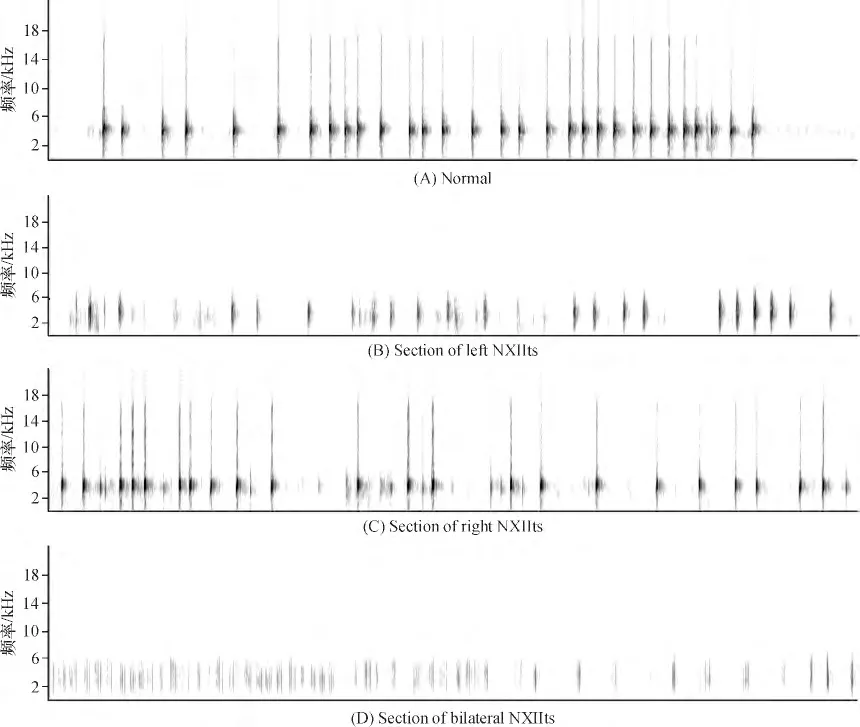
图6 雄鸟断鸣管神经支前后鸣曲的语图对比Figure 6 Songs before and after section of NXIIts in adult male White-rumped Munia
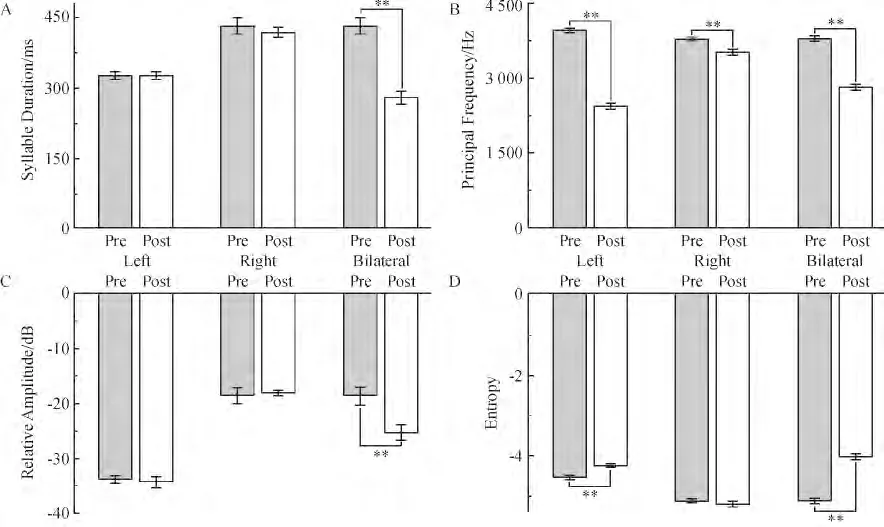
图7 雌鸟断神经前后各声学参数的变化Figure 7 Comparison of acoustic analysis before and after section in female White-rumped Munia
3 讨论
3.1 成年雄性白腰文鸟鸣管神经的左侧优势
鸣禽发声的侧别优势一直为鸟声研究领域所关注.NXIIts 严格的侧别优势使得鸣禽控制声音的能力加强,鸣声变化幅度大[2]. 本实验的研究结果发现,对于雄鸟的鸣叫声,断左侧鸣管神经支将导致音节时长变短、音调降低、主频率及音节整体音量下降;断右侧鸣管神经支则导致音节整体音量增加、调频一致性和音节稳定性增强;断双侧鸣管神经支后音节时长变短、音调降低、主频率及音节整体音量下降.无论切断哪侧鸣管神经支,都对成年雄性白腰文鸟调节频率的能力没有显著影响. 出现上述结果可能是因为左侧鸣管神经支能产生较高的音量和较高的频率,而右侧则主要产生较低的音量和2 kHz 以下的较低的频率[6],因此,断左侧音节整体音量下降且音调较低,而断右侧音节整体音量增加且对音调没有较大影响.正常雄鸟发声时由于两侧鸣管神经的协调作用,因此叫声的频率变化较大,而断右侧鸣管神经后频率和音节更趋于一致,因此调频一致性和音节稳定性增强.
本研究对雄鸟的鸣曲进行断神经前后对比分析后发现,断左侧鸣管神经后音量、音调和音色的损伤程度都比断右侧的严重,这与Secora 等[6]和Urbano等[7]的研究结果一致,进一步验证了成年雄性白腰文鸟发声的左侧优势.
对于雌鸟的鸣叫声,左侧断神经对其声音的损伤也较大,但具体情况有些不同. 经过对比分析,可以推测雌鸟左侧鸣管神经支主要产生高频率的声音,并具有较强的调节音调和音量的能力,而右侧鸣管神经支的作用则比较小. 而雌鸟的音节时长和音量则受左右侧鸣管神经共同作用,只损伤一侧鸣管神经对音节时长和音量影响并不大,这可能是因为两侧神经控制音节时长和音量的能力差异不同所致,当损伤一侧神经时,另一侧神经可以补偿.
成年白腰文鸟的NXIIts 可单侧支配鸣肌,并且具有左侧优势.两性相同点在于两侧鸣管神经支既相互协调又各有侧重,且左侧鸣管神经支对声音的调控作用比右侧强,左侧侧重于产生较高频率的声音,而右侧鸣管神经则能产生较高的熵值,使得音节稳定性和调频一致性增强.两性不同点在于,雄鸟左侧鸣管神经能产生较高的音量和较长的音节,而雌鸟的音量和音节时长两侧均可以独立调节,并且两侧神经对这方面的调节能力差异不大,只有断双侧才能对音量,音调,音色和音节时长产生明显影响.
鸣禽发声的左侧优势,与其高位发声中枢有左侧优势有关,即左侧HVC 和RA 的体积都大于右侧[8].中枢的差异直接导致外周神经支配的差异.
3.2 成年白腰文鸟鸣声的性别差异
本研究证实了白腰文鸟的鸣声存在两性差异.正常情况下,雄鸟能鸣唱而雌鸟不能,并且雄鸟鸣叫的音节时长比雌鸟长,音节的幅值比雌鸟高,音节的频率范围比雌鸟广,调节音量的能力比雌鸟强,但调频一致性、频率调节能力和音节稳定性比雌鸟差.总体上雄鸟的鸣声更具优势.
鸟类的发声是由高级发声中枢(HVC)通过延髓舌下神经气管鸣管亚核(nXIIts)支配鸣肌收缩而实现的.因此,白腰文鸟鸣声的这种性别差异与其发声控制的核团必然存在联系. 李东风等认为鸣禽鸣啭能力表现出显著的性别差异,原因在于发声学习相关的核团体积存在显著的性二态性,并且与体内的激素调节存在一定的联系[9-10].曾少举等[5]和董银兰等[11]研究也表明,白腰文鸟的发声控制核团存在性双态性,具体表现在,雄鸟的3个发声控制核团均大于雌鸟,并且核团中的神经元的数量均超过雌鸟,但神经元的密度小于雌鸟.正是由于雄鸟在核团体积和神经元上数量及密度上的优势,使得雄鸟拥有更多的神经元参与鸣声学习,同时由于血液中具有较高的睾酮水平,使得雄鸟不仅更善于鸣叫,也具有区别于雌鸟的鸣唱行为.
正常雄鸟的调频一致性、频率调节能力和音节稳定性比雌鸟差,说明雌鸟在频率控制方面更具优势,出现此种差异可能是雌鸟频率变化范围小的原因造成的.正是由于频率变化范围较小,因此频率的一致性和音节的稳定性相对较高,FM 的值也因此而提高.此外鸣肌的发育程度差异可能也是影响雌雄白腰文鸟鸣声差异的重要因素[2].
目前关于鸣禽鸣管神经的侧别优势存在左右侧之争.曾贤燕等曾报道斑胸草雀存在右侧优势[12].不同鸣禽侧别支配的差异原因尚有待阐明.
[1]李东风,蓝书成. 鸟类鸣叫学习机理研究回顾[J]. 生理科学进展,1991,22(4):324-328.Li D F,Lan S C. Review of study of song learning in bird[J]. Progress in Physiological Science,1991,22(4):324-328.
[2]耿慧,李东风,蒋锦昌. 鸟类的发声器官与调控机制[J]. 生物物理学报,2005,21(6):397-403.Geng H,Li D F,Jiang J C. Avian vocal organ and vocal control mechanism[J]. Acta Biophysica Sinica,2005,21(6):397-403.
[3]Okanoya K. The Bengalese Finch:A window on the behavioral neurobiology of birdsong syntax[J]. Annals of the New York Academy of Sciences,2004,1016:724-735.
[4]左明雪,曾少举,彭卫民,等. 白腰文鸟发声行为的神经发育[J].动物学报,2002,48(1):50-57.Zuo M X,Zeng S J,Peng W M,et al. Neural development of vocal behavior in Striated manikin (Lonchura striata swinhoei)[J]. Acta Zoological Sinica,2002,48(1):50-57.
[5]曾少举,左明雪,张信文.白腰文鸟发声行为的性别差异及其机制[J].动物学研究,2001,22(1):51-57.Zeng S J,Zuo M X,Zhang X W. The mechanism of sexual difference of vocalization in Lonchura striata swinhoei[J]. Zoological Research,2001,22(1):51-57.
[6]Secora K R,Peterson J R,Urbano C M,et al. Syringeal specialization of frequency control during song production in the bengalese finch (Lonchura striata domestica)[J].PLoS ONE,2012,7(3):e34135.
[7]Urbano C M,Peterson J R,Cooper B G. Exploring vocal recovery after cranial nerve injury in bengalese finches[J]. Neuroscience Letters,2013,534:112-116.
[8]左明雪,蓝书成. 鸣禽发声控制神经通路的左侧优势[J]. 生物学通报,1989(3):7-9.Zuo M X,Lan S C. Left side dominance of vocal control pathway in songbirds[J]. Bulletin of Biology,1989(3):7-9.
[9]李东风,左明雪,蓝书成,等. 鸣禽前脑发声控制核团的雌雄差别[J].动物学报,1992,38(3):298-301.Li D F,Zuo M X,Lan S C,et al. Sex differences of vocal control nuclei in the forebrain of songbirds[J]. Acta Zoological Sinica,1992,38(3):298-301.
[10]李东风,李杰. 鸣禽鸟发声行为的激素调节[J]. 动物学研究,1999,20(1):62-66.Li D F,Li J. The hormonal regulation of vocal behavior in songbirds[J]. Zoological Research,1999,20(1):62-66.
[11]董银兰,张海珠. 白腰文鸟前脑发声控制核团的雄雌差异研究[J].山西师范大学学报:自然科学版,2006,20(1):76-79.Dong Y L,Zhang H Z. The sexual difference of vocal control nuclei of forebrain in White-rumpoed Munia[J]. Journal of Shanxi Teachers University,2006,20(1):76-79.
[12]曾贤燕,李东风. 成年雄性斑胸草雀前脑与中脑对习得性发声控制的侧别差异[J]. 动物学研究,2013,34(1):1-7.Zeng X Y,Li D F. Lateral differences in the forebrain and midbrain control of learned vocalizations in adult male Zebra Finch (Taeniopygia guttata)[J]. Zoological research,2013,34(1):1-7.
