牛瑟氏泰勒虫p23-IL-18融合基因的原核表达
2014-11-23许应天吴艳丽于志云
白 杨,许应天,吴艳丽,于志云
(1.吉林正业生物制品股份有限公司,吉林吉林132101;2.延边大学,吉林延吉133002)
牛瑟氏泰勒虫病是由瑟氏泰勒虫寄生于牛的红细胞和单核巨噬系统细胞内引起的一种蜱传播性血液原虫病[1]。该病在本地牛多数呈带虫,染虫率较低,外地引进牛和改良牛染虫率较高,死亡率也很高[2]。牛瑟氏泰勒虫p23主要表面蛋白是红细胞感染阶段的一种虫体表面蛋白,当用孢子持续感染牛时,虽然瑟氏泰勒虫表面抗原不断发生变化,却始终都能检测到 p23表面蛋白的表达[3]。p23因具有较好的免疫原性[4],故是诊断和预防该病的理想抗原之一[5]。
白细胞介素18(interleukin,IL-18)是近年来新发现的一种重要的细胞免疫调节因子,具有多种生物学功能,其应用研究十分活跃[6-9]。IL-18属于IL-1家族成员,是机体先天性和获得性免疫的重要调节因子。IL-18在慢性炎症、自体免疫性疾病、各种各样的癌变及众多传染病的发生过程中都有表达,所以IL-18自发现以来就一直受到各国研究人员的关注。1999年,IL-18基因的克隆和IL-18活性分析由 Shoda[10]完成。2002 年 Nagata T 等[11]将该基因在家蚕中表达,实验是通过杆状病毒系统完成。目前,研究较多的也是其在参与机体免疫方面的功能,在提高牛的抗感染力、增强灭活苗或基因工程苗免疫作用方面具有广阔的应用前景。本研究以pMD18-T-p23-IL-18质粒为材料,构建牛瑟氏泰勒虫原核表达质粒pET-28a-p23-IL-18,并进行表达,以期为牛瑟氏泰勒虫病基因工程亚单位疫苗的制备提供了理论依据。
1 材料与方法
1.1 质粒、菌种和载体 质粒pVAXI-p23、pMD18-IL-18、DH5α菌种、pET-28a载体菌液均由延边大学预防兽医实验室保存。
1.2 主要试剂 DNA Ligation Kit Ver.2.1,pMD18-T-Simple 载体,T4 DNA Lingase,BamH I,EcoR I,ExTaq酶,质粒小量抽提试剂盒,逆转录酶,DNA Maker,FITC标记的羊抗牛IgG,RNase和巯基乙醇,均为Sigma公司产品。
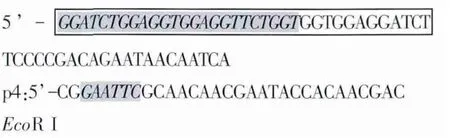
1.3 引物的设计与合成 根据GenBank中报道的牛瑟氏泰勒虫p23序列(D84447)及牛白细胞介素18cDNA序列(BC102879)设计了两对特异性的引物。其中含有长度为45 bp的linker序列为中间接头(Gly4Ser)3核苷酸,由p2和p3共同编码,即方框的部分。p2引物与p3引物的5’端有24 bp的碱基互补,为阴影部分。在p1与p4引物上含有酶切位点,设计的引物由上海生工生物工程技术服务有限公司合成。p1和p2用于扩增IL-18基因序列,预测扩增长度为526 bp,重组后目的片段长度约为1048 bp。
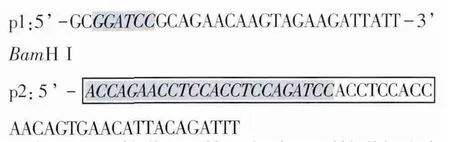
p3和p4用于扩增p23基因序列,预测扩增长度为522 bp。p3:
1.4 重组基因的构建 参考文献刘娟等的试验方法[12]以现有质粒pMD18-IL-18为模板,以p1和p2为引物,扩增IL-18基因片段;以pVAXI-p23为模板,以p3和p4为引物,扩增p23基因片段。将以上2次PCR产物混合作为PCR反应模板,以pl和p4为引物,进行PCR扩增,重组基因p23-IL-18。
1.5 克隆载体pMD18-T-p23-IL-18的构建及鉴定 将纯化后的p23-IL-18与pMD18-TSimple Vector进行连接。连接体系:PCR回收产物4 μL,pMD18-T-Simple Vector 1 μL,Vector Buffer 5 μL,4℃链接过夜。连接产物转化感受态DH5α大肠杆菌。挑取经Amp抗性筛选的菌落后培养,提取质粒,经PCR、酶切鉴定正确后命名为pMD18-T-p23-IL-18。
1.6 原核表达载体pET-p23-IL-18的构建及鉴定 将pMD18-T-p23-IL-18与pET-28a载体分别用BamH I和EcoR I进行双酶切,酶切产物纯化后用T4 DNA连接酶连接。连接体系为:T4 DNA Lingase 1 μL,T4 DNA ligation buffer 2.5 μL,p23-IL-18 10 μL,pET-28a 2 μL,16 ℃连接过夜。连接产物转化感受态DH5α大肠杆菌,小量提取质粒。经 PCR(引物为 p1和 p4)、酶切(BamH I和EcoR I)、测序鉴定正确后的重组质粒命名为pET-p23-IL-18。
1.7 重组表达质粒的诱导表达 将鉴定为阳性的重组表达菌50 μL接种于5 mL LB培养液(含Amp 100 μg/mL)中,37℃过夜培养后,按1%接种于新鲜 LB培养液中,振摇至OD600nm为0.6,加入IPTG至终浓度为1 mmol/L,37℃诱导表达。分别在0、2、4、6和8 h各收集2 mL菌液,同时收集未诱导的菌液作为阴性对照。SDS-PAGE电泳后考马斯亮蓝染色观察,并作Western-blotting分析。
2 结果与分析
2.1 目的基因的扩增 取5 μL的PCR产物使用21.0%琼脂糖凝胶进行电泳检测。p23与IL-18基因扩增结果显示均在500 bp左右出现特异性条带,与预期大小相符合(图1);重组后基因扩增结果显示在1000 bp左右出现特异性条带,与预期大小相符合(图2)。
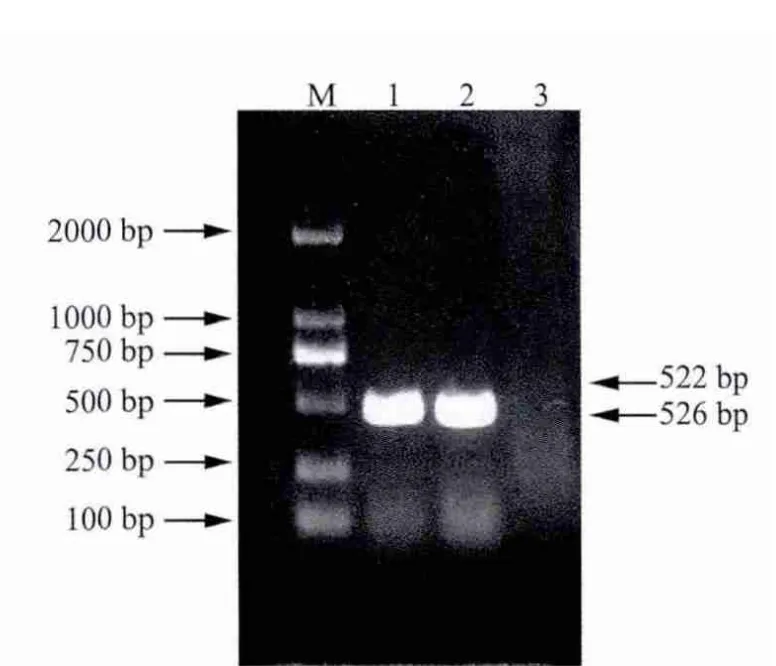
图1 p23与IL-18基因的PCR扩增
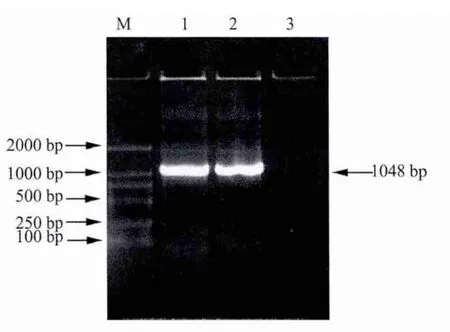
图2 p23-IL-18融合基因的PCR扩增
2.2 阳性克隆载体的鉴定 重组克隆质粒用特异性引物p1,p4扩增出1000 bp左右出现特异性条带,然后用 EcoRⅠ和 BamHⅠ酶切出长度为1000 bp左右的片段(图3),说明目的片段已成功扩增并正确插入到克隆载体pMD-18-T Simple中。测序结果表明,克隆得到的重组基因p23-IL-18基因片段大小为1048 bp。此基因片段与GenBank中已发表的牛瑟氏泰勒虫p23核苷酸序列、牛IL-18核苷酸序列同源性为99%。
2.3 重组表达质粒pET-28a-p23-IL-18的鉴定 用BamHⅠ和EcoRⅠ对初步筛选为阳性的重组质粒进行酶切鉴定,酶切产物于1%琼脂糖凝胶中电泳。结果显示,经BamHⅠ和EcoRⅠ酶切,得到了约3000 bp和1048 bp的两条片段(图4),说明目的基因片段已正确地克隆到pET-28a原核表达载体中。
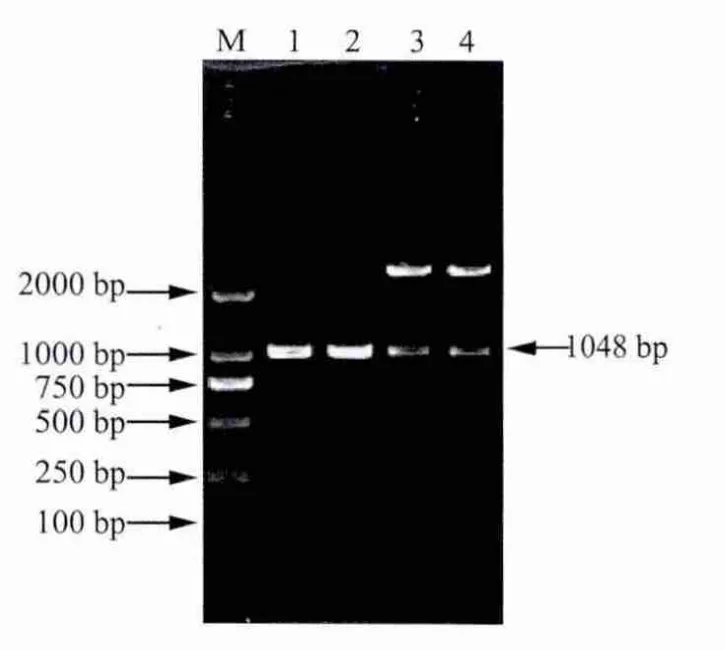
图3 重组质粒pMD18-T-p23-IL-18的PCR及酶切鉴定结果

图4 重组质粒pET-28a-p23-IL-18的PCR及酶切鉴定图
2.4 目的基因的诱导表达 诱导的pET-28ap23-IL-18阳性菌在约48 kDa处出现明显特异性条带(图5),与预期结果相符,重组表达菌在6 h时表达量最高。
2.5 表达产物的Western-blotting检测 表达产物经SDS-PAGE电泳后转移到PVDF膜上进行Western-blotting检测,在PVDF膜上出现特异性条带,表达产物能被牛瑟氏泰勒虫阳性血清所识别,证明目的基因的表达产物具有较好的反应原性(图6)。
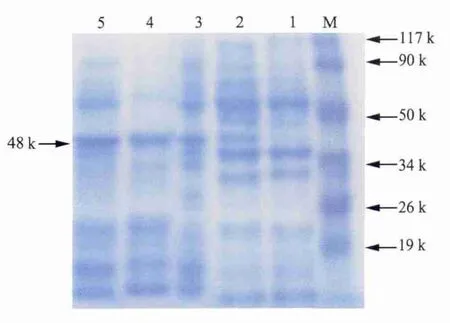
图5 表达产物的SDS-PAGE电泳

图6 表达产物的Western-blotting分析
3 讨论
细胞因子通过复杂网络调节机体免疫,细胞因子失衡或调节异常都与疾病有着密切的关系。IL-18是一种很强的IFN-γ诱导剂,其生物学活性是通过调节IFN-γ的表达来提高机体细胞免疫水平。因此BolIL-18可望成为疫苗的一种重要的细胞免疫佐剂[13-15],在提高牛的抗感染力、增强灭活苗或基因工程苗免疫作用方面具有广阔的应用前景。本实验利用现有质粒为模板,将牛白细胞介素18与牛瑟氏泰勒虫的p23表面蛋白基因通过重叠延伸拼接聚合酶链式反应(SOE-PCR)将两段基因串连在一起,并连接到原核表达载体上,构建原核表达重组质粒。Western-blotting结果证实表达产物能被牛瑟氏泰勒虫阳性血清所识别,证明表达产物具有较好的反应原性。
牛瑟氏泰勒虫p23基因全长672 bp,具有完整的开放阅读框,编码223个氨基酸,N末端有28个氨基酸残基的信号肽序列,疏水区多缬氨酸,C末端是跨膜区域,69~71位有一个潜在的N糖基化位点,为ASn-Ile-Ser,p23是单拷贝基因,并有等位基因。化学组成分析酪氨酸含量较多,弥补了p23作为抗原蛋白分子量较小的不足,且亲水性较强,膜表面抗原决定簇丰富,具有疫苗候选分子的基础。
在进行原核细胞表达时,多以融合蛋白为表达形式作为载体,这样既可防止宿主菌的蛋白醇对所表达目的蛋白的降解,也有利于表达蛋白的纯化,最终保护目的蛋白的活性。故本试验选用pET-28a为载体,可将外源蛋白表达在细胞质内,在N端和C端都能加His-tag,pET-28a没有本身溶解性髙的多肽融合蛋白,也没有催化二硫键形成的酶融合蛋白,而且不含信号肽序列,这为纯化目的蛋白提供了有利条件。同时pET-28a上的大部分酶切位点都不会造成阅读框移位。
基于上述研究,本实验将IL-18基因和p23基因融合在一起,成功构建了双基因融合的基因工程菌株,并探讨p23和IL-18双基因融合产物作为亚单位疫苗成分的可行性[16],为探索牛瑟氏泰勒虫双基因工程疫苗奠定了理论依据。
[1]蒋金书.动物原虫病学[M].北京:中国农业大学出版社,2001.
[2]Schnittger Leonhard,Katzer Frank,Biermann Reinhild,et al.Characterization of a polymorphic Theileria annulata surface protein(TaSP)closely related to PIM of Theileria parva:implications for use in diagnostic tests and subunit vaccines[J].Molecularandbiochemicalparasitology, 2002, 120(2):247-256.
[3]Wen Zhong Zhuang,Chihiro Sugimoto,Shuichi Kubota,et al.Antigenic alteration in major piroplasm surface proteins of Theileria sergenti during infection[J].Veterinary Parasitology,1995,60(3/4):191-198.
[4]金春梅,许应天,张守发,等.牛瑟氏泰勒虫P33主要表面蛋白基因的克隆与序列分析[J].中国预防兽医学报,2007,(2):112-114.
[5]Zhang W,Sugimoto C,Matsuba T,et al.Analysis of antigentic and genetic of Theileria sergenti major piroplasm surface proteins[J].Net Med Sci,1994,56:469-473.
[6]Bokarewa M,Hultgren O.Is interleukin-18 useful for monitoring rheumatoid arthritis?[J].Scandinavian journal of rheumatology,2005,34(6):433-436.
[7]Yano Ai,Nakao Kazushi,Sarai Ai,et al.Elevated serum interleukin-18 levels might reflect the high risk of hospitalization in patients on peritoneal dialysis[J].Nephrology(Carlton ,Vic.),2005,10(6):576-582.
[8]Thong-Ngam D,Tangkijvanich P,Lerknimitr R,et al.Diagnostic role of serum interleukin.1 8 in gastric cancer patients[J].World J Gastroenterol,2006,12(28):4473-4477.
[9]Shinoda Masahiro,Wakabayashi Go,Shimazu Motohide,et al.Increased serum and hepatic tissue levels of interleukin-18 in patients with fulminant hepatic failure[J].Journal of gastroenterology and hepatology,2006,21(11):1731-1736.
[10]Shoda L K,Zarlenda D S,Hirano A.Cloning of a cDNA encoding bovine interleukin-18 and analysis of IL-18 expression in macrophages and its IFN-gamma-inducing activity[J].J Interferon Cytokine Res,1999,(10):1169-1177.
[11]Nagata T,Ishikawa S,Shimokawa E,et al.High level expression and purification of bioactive bovine interleukin-18 using a baculovirus system[J].Vet Immunity,2002,87(8):65-72.
[12]刘 娟,刘明远,于 录,等.RV-G/LTB双基因融合真核质粒表达载体的构建并在vero细胞中的表达[J].中国生物制品学杂志,2011,24(3):255-258.
[13]邢秀娟.牛IL-18真核表达载体的构建及其对FMD疫苗免疫增强作用的研究[D].泰安:山东农业大学,2008.
[14]叶丽萍,王春凤.细胞因子免疫佐剂效应机制及其在兽医临床中的应用[J].中国畜牧兽医2012,(11):196-199.
[15]刘建文,李月辉,王健春.基因疫苗及其免疫佐剂研究进展[J].中国兽医杂志,2007,(3):41-43.
[16]孙平杰.牛瑟氏泰勒虫p23表面蛋白基因与牛白细胞介素18基因在原核和真核细胞中的串联表达[D].延边:延边大学,2012.
