突变和野生型TDP-43对运动神经元电压门控钠通道的影响
2014-09-04王小新吴力娟段伟松河北医科大学第二医院神经内科河北省神经病学重点实验室河北石家庄050000北京市密云县医院神经内科北京密云0500河北省邢台市第三医院神经内科河北邢台054000
董 惠,王小新,吴力娟,段伟松,许 蕾*(.河北医科大学第二医院神经内科,河北省神经病学重点实验室,河北 石家庄 050000;.北京市密云县医院神经内科,北京 密云 0500;.河北省邢台市第三医院神经内科,河北 邢台 054000)
·论著·
突变和野生型TDP-43对运动神经元电压门控钠通道的影响
董 惠1,王小新2,吴力娟3,段伟松1,许 蕾1*
(1.河北医科大学第二医院神经内科,河北省神经病学重点实验室,河北 石家庄 050000;2.北京市密云县医院神经内科,北京 密云 101500;3.河北省邢台市第三医院神经内科,河北 邢台 054000)
目的观察反式激活应答区域DNA结合蛋白43(transactivating response region DNA binding protein,TDP-43)对运动神经元电压门控钠离子通道(voltage-gated sodium channels,VGSCs)特性的影响,评价突变(M337V)和野生型TDP-43与运动神经元变性的关系。方法选择运动神经元样细胞系,稳定转染空的pCI-neo质粒(对照组)或者携带M337V突变(M337V组)和人野生型(WT组)TDP-43 cDNA的质粒;采用全细胞膜片钳技术记录M337V组、WT组和对照组3类细胞VGSCs激活和失活电流,分析VGSCs特性的变化。结果WT组VGSCs激活曲线的半数激活电压以及缓慢失活后恢复曲线的时间常数明显小于M337V组和对照组(P<0.05或<0.01),提示野生型TDP-43加快了VGSCs激活和慢失活恢复的特性;而M337V组VGSCs稳态失活曲线的半数失活电压以及快速失活后恢复曲线的时间常数明显大于WT组和对照组(P<0.05或<0.01),提示M337V突变TDP-43抑制运动神经元样细胞VGSCs快速失活和激活后恢复能力。结论M337V突变或人野生型TDP-43可导致VGSCs功能改变,增高运动神经元兴奋性,但二者从不同角度影响VGSCs的特性,提示这两种TDP-43通过不同的机制介导和参与运动神经元变性。
运动神经元;DNA结合蛋白质类;钠通道,电压门控
10.3969/j.issn.1007-3205.2014.07.021
反式激活应答区域DNA结合蛋白43(transactivating response region DNA binding protein,TDP-43)是大多数肌萎缩侧索硬化(amyotrophic lateral sclerosis,ALS)和额颞叶痴呆(frontotemporal lobar degeneration,FTLD)患者泛素化包涵体的主要成分[1]。编码TDP-43的基因TARDBP与散发型和家族型ALS相关[2-3],提示TDP-43的改变可直接引起神经元变性。临床研究[4-7]显示,ALS患者皮层运动神经元兴奋性增加同时皮层间抑制减小,甚至这种高兴奋性是散发型ALS的早期特点,也可预示家族型ALS的临床发病,但是TDP-43如何影响运动神经元兴奋水平还不清楚。细胞产生动作电位的能力被称为兴奋性,电压门控钠离子通道(voltage-gated sodium channels,VGSCs)是可兴奋细胞暴发动作电位以及动作电位延细胞膜传递的关键,其激活和失活功能改变直接影响细胞兴奋水平。本研究通过观察表达人类突变(M337V)和野生型TDP-43的运动神经元样细胞VGSCs性质的改变,探讨VGSCs在TDP-43所致神经元变性中的作用。
1 材料与方法
1.1 建立稳定表达突变和人野生型TDP-43的运动神经元样细胞系:NSC34细胞是一种保留增殖功能和表达多种运动神经元特性的杂交细胞系。NSC34细胞分别转染pCl-neo空质粒(对照组)、克隆人类野生型TDP-43 cDNA质粒(WT组)和克隆人M337V突变的TDP-43 cDNA质粒(M337V组),3类质粒均携带HA标签,转染方法与我们以往的研究[8]相同。
1.2 VGSCs的记录:采用全细胞模式电压钳方法记录对照组、WT组和M337V组NSC34细胞的VGSCs电流(INa),将硼硅酸玻璃电极拉制成电阻4~6MΩ的微电极,尖端热抛光处理。电极內液,130mmol/L CsCl,20mmol/L氯化四乙胺,1mmol/L MgCl2,0.24mmol/L CaCl2,10mmol/L右旋葡萄糖,5mmol/L乙二醇双四乙酸,2mmol/L三磷酸腺苷二钠,10mmol/L羟乙基哌嗪乙磺酸,用CsOH调pH值至7.2~7.3。细胞外液成分为120mmol/L NaCl,3mmol/L KCl,2mmol/L MgCl2,2mmol/L CaCl2,10mmol/L右旋葡萄糖,10mmol/L羟乙基哌嗪乙磺酸,用NaOH调pH值至7.2~7.3,加入0.2mmol/L氯化铬和1mmol/L 4-氨基吡啶阻滞Ca2+和K+电流。所有试剂均购自Sigma公司。整个实验过程在室温(22~24°C)条件下进行,当电极尖端接触到细胞膜时,释放正压并稍施负压形成封接(电阻>2GΩ),吸破细胞膜,膜片钳放大器(德国HEKA EPC10)Patchmaster记录到膜电流,表明细胞膜已经吸破而形成全细胞记录模式,记录细胞电容(C),电容和串联电阻的补偿>85%,采用频率为50kHz。形成全细胞记录模式后,将电位钳制在-100mV,待电流稳定,去极化到-10mV时长为10ms,可记录到一个快速激活快速失活的内向电流,此电流可以被1μmol/L河豚毒素(Tetrodotoxin,TTX,Sigma,美国)完全阻断,证实为TTX敏感型INa。每个细胞所记录到的电流除以自身电容(INa/C),得到单位表面积上电流的大小,即电流密度(pA/pF),以减少误差。
1.3 INa数据拟合和性质分析:VGSCs电压依赖的激活和稳态失活,均采用Boltzmann方程进行拟合,I/C=1-{1/[1+exp(V-V1/2)/k]},其中I/C代表电流密度,V代表刺激电压,V1/2是半数激活或失活电压,k为曲线斜率,这些参数表现了VGSCs激活和失活电压依赖性的特征。VGSCs快速失活恢复以及慢失活恢复,均采用单指数方程进行拟合,I△t/Imax=A1×exp(-t/τ),其中I△t代表电流峰值,A1代表时间常数τ对应的电流恢复比例,t代表时间,Imax代表第1个去极化电流的幅度。
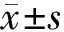
2 结 果
2.1 VGSCs激活的电压依赖性:电位钳制在-100mV,给予-60~+45mV的阶梯去极化脉冲刺激,步幅为5mV,刺激时长为10ms,每两个刺激循环之间的时间间隔为500ms,可以记录到一系列VGSCs电流(图1A,B)。
电流密度-电压(INa-V)曲线(图1C)显示,对照组(n=12)、WT组(n=17)和M337V组(n=16)均存在电压依赖性,在-35mV脉冲刺激时可激活VGSCs产生INa,随着刺激电压的增大,INa也逐渐增大,在刺激电压为-10mV时INa达到峰值,分别是-95.00±8.83、-81.36±10.62和-81.92±10.86pA/pF,3组细胞的峰值差异无统计学意义(P>0.05),3组细胞在达到峰值后逐渐减小,刺激电压从-35~+45mV所对应的对照组细胞INa是从-10.04±9.61~-34.53±5.36pA/pF,WT组是从-26.25±16.61~-42.18±7.56pA/pF,M337V组是从-13.60±9.69~-29.50±6.36pA/pF,3组间各个刺激电压所对应的INa比较差异均无统计学意义(P>0.05)。
将3组细胞-50~-5mV所对应的INa进行Boltzmann方程拟合(图1D)。对照组、WT组和M337V组半数激活电压V1/2分别为-28.04±0.35、-32.75±0.69、-28.93±0.38mV,其中WT组与对照组比较差异有统计学意义(P<0.05),M337V组与WT组比较差异有统计学意义(P<0.05),而M337V组与对照组比较差异无统计学意义(P>0.05);对照组、WT组和M337V组的斜率κ分别为3.68±0.31、3.58±0.62和3.81±0.35,3组之间比较差异均无统计学意义(P>0.05)(图1E)。

图1 VGSCs激活的电压依赖性(电流密度-电压曲线)
*P<0.05与WT组比较 #P<0.05与对照组比较(q检验)
A.电压刺激模式图;B.M337V组,WT组和对照组细胞的VGSCs电流图;C.3组VGSCs电流密度-电压曲线;D.3组VGSCs激活拟合曲线;E.3组半数激活电压V1/2和斜率k
2.2 VGSCs的稳态失活:钳制电压-100mV,给予从-100~-20mV时长为40ms系列预刺激,步幅为10mV,再去极化到+10mV,时长10ms,最后恢复到钳制电压,每个循环间隔500ms(图2A)。图2B显示记录到的对照组、WT组和M337V组VGSCs电流。预刺激为-100mV时对应的电流最大(Imax),其他预刺激所对应的INa除以Imax得到电流比例值(I/Imax),对3组I/Imax值进行Boltzmann方程拟合得到图2C。可见对照组、WT组和M337V组均随着预刺激电压的增大VGSCs逐渐失活,至-20mV时VGSCs完全失活。在预刺激电压为-50、-40和-30mV时,WT组细胞I/Imax值分别为0.60±0.08、0.29±0.10和0.08±0.07,对照组分别为0.58±0.05、0.31±0.06和0.08±0.04,2组比较差异无统计学意义(P<0.05)。在预刺激电压为-40和-30mV时M337V组I/Imax值分别为0.70±0.04,0.46±0.07和0.13±0.06,明显大于对照组,差异有统计学意义(P<0.05)。
对照组、WT组和M337V组半数失活电压V1/2分别为-45.61±0.97、-45.98±0.65和-36.81±0.92mV,M337V组较WT组和对照组均有明显的增加,差异有统计学意义(P<0.05);对照组、WT组和M337V组的斜率k分别为10.60±0.83、9.33±0.56和12.11±0.56,M337V组较WT组和对照组均有明显的增加,差异有统计学意义(P<0.05)(图2D)。
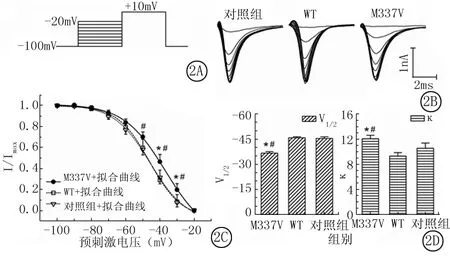
图2 VGSCs稳态失活
*P<0.05与WT组比较 #P<0.05与对照组比较(q检验)
A.电压刺激模式图;B.M337V组,WT组和对照组细胞VGSCs稳态失活电流图;C.3组VGSCs稳态失活拟合曲线;D.3组半数失活电压V1/2和斜率k
2.3 VGSCs快速失活的恢复:钳制电位-100mV,给予去极化到-10mV时长10ms的方波刺激,间隔后给予同样的方波刺激,2个方波之间的间隔时间从0.5ms开始,每个循环增加0.5ms,逐渐递增至最大间隔45ms(图3A)。图3B显示记录到的对照组、WT组和M337V组VGSCs快速恢复电流。
每个循环,均以第二个方波刺激所引出的电流峰值(I)除以第一个方波刺激所引出的电流峰值(Imax),得到比例值(I/Imax),以I/Imax值与对应的时间间隔作图,并对各组数据进行单指数方程拟合,得到图3C,显示M337V组与对照组比较曲线右移。对照组、WT组和M337V组VGSCs的快速失活恢复的时间常数(τ)分别为5.62±0.10、6.47±0.14和7.42±0.08ms,其中M337V组与对照组比较有明显的增大,差异有统计学意义(P<0.01)(图3D)。
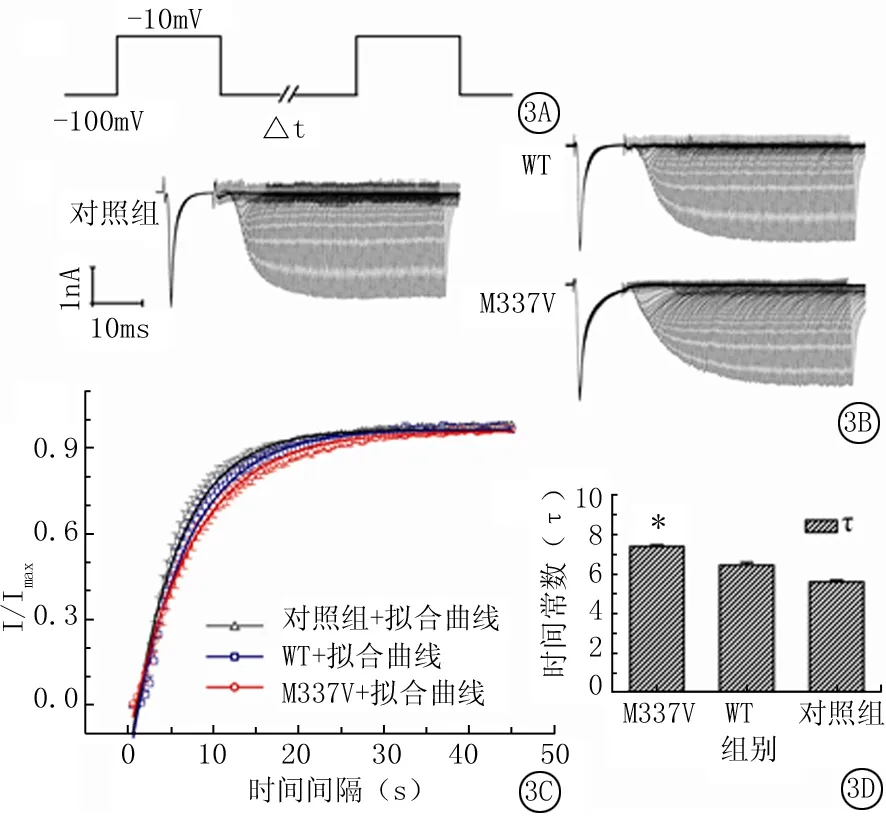
图3 VGSCs快速失活的恢复
*P<0.01与对照组比较(q检验)
A.电压刺激模式图;B.M337V组,WT组和对照组细胞VGSCs快速失活后恢复的电流图;C.3组VGSCs快速失活后恢复的拟合曲线;D.3组VGSCs快速失活后恢复的时间常数τ
2.4 VGSCs缓慢失活的恢复:钳制电压-100mV,给予-10mV、10s长时程方波刺激,间隔1s以使所有钠通道从快速失活状态恢复,再给予1个-10mV,5ms的短时程刺激,检测钠通道进入缓慢失活状态所占的比例,之后每间隔3s再次给予相同的短时程刺激,直到总间隔时间为25s(图4A)。
以长时程刺激所得到的电流幅度值为Imax,随后的不同时间间隔的短时程刺激得到的电流幅度(I)除以Imax,得到I/Imax。以间隔时间为横坐标,对1~25s对应的I/Imax值进行单指数方程拟合制成图4B,显示在间隔时间为1和4s时WT组I/Imax值(0.72±0.03vs0.90±0.02)明显大于对照组(0.50±0.03和0.80±0.02)和M337V组(0.48±0.04和0.80±0.02),差异有统计学意义(P<0.01或<0.05)。
对照组、WT组和M337V组缓慢失活恢复的时间常数(τ)分别为3.30±0.31、1.94±0.14和3.25±0.24s,WT组明显小于M337V组和对照组,差异有统计学意义(P<0.01)(图4C)。
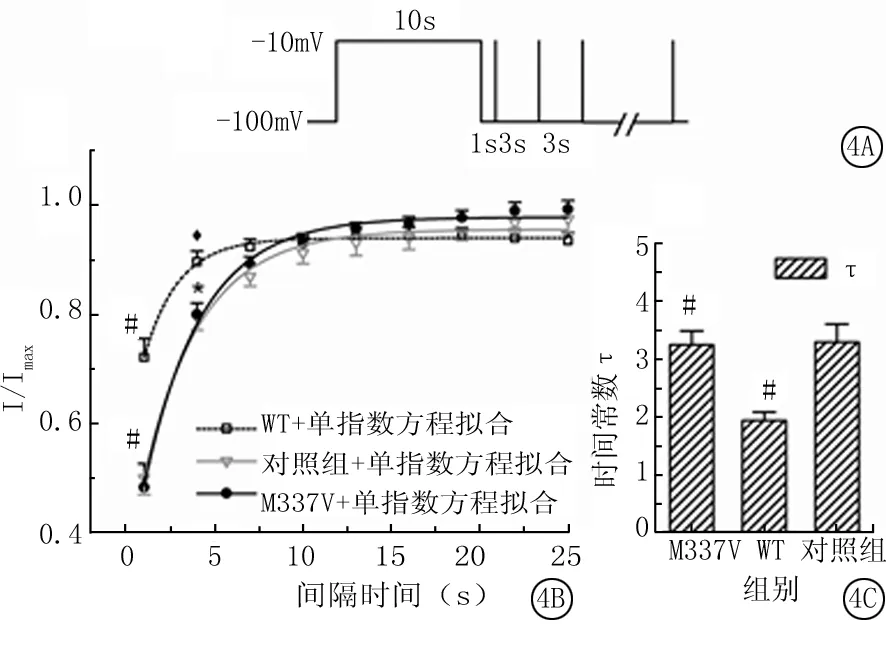
图4 VGSCs缓慢失活的恢复
*P<0.01与WT组比较 #P<0.01与对照组比较(q检验)
A.电压刺激模式图;B.M337V组,WT组和对照组细胞VGSCs缓慢失活恢复的拟合曲线;C.3组VGSCs缓慢失活恢复的时间常数τ
3 讨 论
人TDP-43是一种高保守、普遍存在的核蛋白,可结合DNA和RNA[9],主要参与RNA的代谢,包括剪切、转录抑制、miRNA的合成、mRNA胞核胞浆穿梭,以及RNA转运和稳定[10],它的功能障碍可导致其下游多种功能蛋白异常,到目前为止,已发现超过40种TDP-43的结构域突变与家族性或散发性ALS相关[11]。本研究发现,表达TDP-43 M337V突变的运动神经元VGSCs激活后稳态失活速度以及快速失活后的恢复速度均明显减慢。提示M337V突变的TDP-43主要影响运动神经元样细胞VGSCs失活和激活后恢复的过程。VGSCs失活和激活后恢复能力的下调,必然会导致神经元动作电位时间特性的改变,重要的是在神经信号的传导过程中,动作电位频率和时间特性在编码神经冲动中起关键作用[12],即使很小的改变也可能导致神经元间信息传递异常,进而影响正常生理过程。
然而大部分的ALS和FTLD患者都没有合并TDP-43突变,同时有研究[13-14]发现,过表达人类野生型TDP-43的转基因小鼠表现出脊髓和皮层运动神经元变性,野生型TDP-43可下调内源性TDP-43,导致TDP-43蛋白截断和聚集,最终提高胞浆和胞核泛素化水平、线粒体异常聚集以及轴索变性。本研究结果发现,表达人类野生型TDP-43的运动神经元样细胞VGSCs半数激活电压减小,说明细胞膜电位去极化较小的范围即可使50%VGSCs开放。同时这类细胞VGSCs进入缓慢失活的比例明显减少,其随后缓慢失活恢复的速度也明显高于M337V组和对照组,均提示表达人类野生型TDP-43可使运动神经元易产生动作电位,处于一种高兴奋水平,进而加重氧化应激[15],改变线粒体功能[16]导致能量耗竭,推进临床症状的发展。同样临床研究[17]也证实,无论是家族性还是散发性ALS患者均表现出皮层高兴奋性,推测细胞膜离子通道功能变化损害了沿运动皮层传导的抑制和兴奋通路,并且这种损害与病程和临床分期有关。
VGSCs拥有2种失活特性,快失活和慢失活。神经元短时间(数毫秒)去极化过程中钠电流的快速衰减即为快失活,而慢失活发生在细胞膜长时间(数秒或数分钟)去极化后。快失活保证了动作电位的短暂性以及不应期的产生,本研究发现M337V突变TDP-43主要抑制了运动神经元VGSCs快失活功能,而人野生型TDP-43主要改变慢失活的程度,这似乎也提示这两种蛋白在ALS致病通路中起不同的作用,但最终均导致选择性运动神经元变性。
综上所述,M337V突变TDP-43抑制运动神经元样细胞VGSCs快速失活和激活后恢复能力,人野生型TDP-43加快了VGSCs激活和慢失活恢复的特性,提示这两种TDP-43通过不同的机制导致和参与ALS的病理生理过程。
[1] NEUMANN M,SAMPATHU DM,KWONG LK,et al.Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis[J].Science,2006,314(5796):130-133.
[2] SREEDHARAN J,BLAIR IP,TRIPATHI VB,et al.TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis[J].Science,2008,319(5870):1668-1672.
[3] KABASHI E,VALDMANIS PN,DION P,et al.TARDBP mutations in individuals with sporadic and familial amyotrophic lateral sclerosis[J].Nat Genet,2008,40(5):572-574.
[4] KANAI K,KUWABARA S,MISAWA S,et al.Altered axonal excitability properties in amyotrophic lateral sclerosis: impaired potassium channel function related to disease stage[J].Brain,2006,129(Pt 4):953-962.
[5] SHIBUYA K,MISAWA S,ARAI K,et al.Markedly reduced axonal potassium channel expression in human sporadic amyotrophic lateral sclerosis: an immunohistochemical study[J].Exp Neurol,2011,232(2):149-153.
[6] VUCIC S,ZIEMANN U,EISEN A,et al.Transcranial magnetic stimulation and amyotrophic lateral sclerosis: pathophysiological insights[J].J Neurol Neurosurg Psychiatry,2013,84(10):1161-1170.
[7] VUCIC S,CHEAH BC,YIANNIKAS C,et al.Cortical excitability distinguishes ALS from mimic disorders[J].Clin Neurophysiol,2011,122(9):1860-1866.
[8] DUAN W,LI X,SHI J,et al.Mutant TAR DNA-binding protein-43 induces oxidative injury in motor neuron-like cell[J].Neuroscience,2010,169(4):1621-1629.
[9] BURATTI E,BARALLE FE.Characterization and functional implications of the RNA binding properties of nuclear factor TDP-43,a novel splicing regulator of CFTR exon 9[J].J Biol Chem,2001,276(39):36337-36343.
[10] LAGIER-TOURENNE C,POLYMENIDOU M,CLEVELAND DW.TDP-43 and FUS/TLS: emerging roles in RNA processing and neurodegeneration[J].Hum Mol Genet,2010,19(R1):R46-64.
[11] JANSSENS J,KLEINBERGER G,WILS H,et al.The role of mutant TAR DNA-binding protein 43 in amyotrophic lateral sclerosis and frontotemporal lobar degeneration[J].Biochem Soc Trans,2011,39(4):954-959.
[12] HARRIS KD,HENZE DA,HIRASE H,et al.Spike train dynamics predicts theta-related phase precession in hippocampal pyramidal cells[J].Nature,2002,417(6890):738-741.
[13] WILS H,KLEINBERGER G,JANSSENS J,et al.TDP-43 transgenic mice develop spastic paralysis and neuronal inclusions characteristic of ALS and frontotemporal lobar degeneration[J].Proc Natl Acad Sci USA,2010,107(8):3858-3863.
[14] XU YF,GENDRON TF,ZHANG YJ,et al.Wild-type human TDP-43 expression causes TDP-43 phosphorylation,mitochondrial aggregation,motor deficits,and early mortality in transgenic mice[J].J Neurosci,2010,30(32):10851-10859.
[15] HAND CK,ROULEAU GA.Familial amyotrophic lateral sclerosis[J].Muscle Nerve,2002,25(2):135-159.
[16] HEATH PR,SHAW PJ.Update on the glutamatergic neurotransmitter system and the role of excitotoxicity in amyotrophic lateral sclerosis[J].Muscle Nerve,2002,26(4):438-458.
[17] ZANETTE G,TAMBURIN S,MANGANOTTI P,et al.Different mechanisms contribute to motor cortex hyperexcitability in amyotrophic lateral sclerosis[J].Clin Neurophysiol,2002,113(11):1688-1697.
2014-05-15;
2014-07-10
国家自然科学基金项目(81171210);河北省卫生厅指导性课题(20110327)
董惠(1978-),女,河北唐山人,河北医科大学第二医院主治医师,医学博士,从事神经内科疾病诊治研究。
*通讯作者。E-mail:xldoc@126.com
R338.1
B
1007-3205(2014)07-0801-05
