模拟酸雨对3种本地草本植物的胁迫效应
2014-08-28柯展鸿王远智宋莉英孙兰兰彭长连
柯展鸿, 王远智, 宋莉英, 孙兰兰, 彭长连*
(1.华南师范大学生命科学学院,广州 510631; 2.广州大学生命科学学院,广州 510006)
酸雨污染对生态系统和人类生存环境造成严重影响,已成为全球性重要的环境问题. 目前,对酸雨的研究,大多涉及酸雨成因、时空分布和治理对策,对农作物、经济作物和针叶树种等植物的伤害机理以及对水陆生态系统的影响等方面[1]. 但是,关于酸雨危害程度的大小和作用机理方面仍有许多未了解,如酸雨危害下植物的光合生理和适应性反应就是其中的问题[2];在酸雨高发区,环境绿化及生态恢复过程中还有许多问题亟待解决,如土壤酸化,土壤盐基离子大量淋失、土壤微生物生长受到影响以及植物难以生长等. 因此,找到适应性强、缓冲能力高的植物无疑具有重要意义.
华泽兰(EupatoriumchinenseLinn.)、鸡矢藤[Paederiascandens(Lour.) Merr.]和蟛蜞菊[Wedeliachinensis(Osbeck.)Merr.]为华南地区常见的几种本地药用草本植物[3-5],具有草本植物典型的生长迅速, 茎叶繁茂,根系发达等特点,对水土保持和环境绿化具有重要作用.
本研究通过比较3种草本植物在光合生理和抗性生理方面对酸雨胁迫的响应差异,探讨草本植物对酸雨胁迫的敏感性和耐受能力,阐明酸雨对植物光合生理和抗性生理的影响机制,为酸雨地区开展植物绿化及生态恢复提供理论参考依据.
1 材料与方法
1.1 材料
试验所用植物材料华泽兰(EupatoriumchinenseLinn.)采自广州市华南植物园内的自然种群,鸡矢藤[Paederiascandens(Lour.) Merr.]和蟛蜞菊[Wedeliachinensis(Osbeck.)Merr.]采自华南师范大学标本园.
选取扦插2~4周后生长健壮、长势一致的再生小苗移栽到小花盆中(直径10 cm, 高5 cm, 容积250 mL),基质采用营养土、河沙和塘泥(体积比为1∶1∶1)的混合物,设置3个酸雨浓度梯度,每个梯度6盆,每盆移栽1株小苗.
1.2 方法
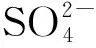
1.2.2 叶片伤斑面积测定及受害等级划分 酸雨处理30 d后,每个处理选取4株植物,摘取同一叶位的成熟叶片,用透明方格坐标纸测得叶片伤斑面积和总面积,以伤斑面积占总叶面积的百分比表征叶片的受伤害程度;参考宋莉英等[7]的分级标准,将叶片受酸雨胁迫伤害分为5个等级:N:叶片没有可见伤害;Ⅰ:叶片受害轻微,受害叶面积<5%;Ⅱ:叶片受害较明显,受害面积约5%~10%;Ⅲ:叶片受害明显,受害面积约10%左右;Ⅳ:叶片受害较重,受害面积约10%~20%.
1.2.3 光合特性测定 酸雨处理30 d后,每个处理选取4株,每株1片成熟叶,于晴天上午9:00—11:30测定叶片净光合速率(Pn)-光强响应曲线.设定进入叶室的CO2浓度(Ca)为 380 μmol/mol,光量子通量密度(photosynthetic photon quanta flux density, PPFD)依次为1 500、1 200、800、600、300、150、100、80、50、20、0 μmol/(m2·s),自高光强开始测定. 依据 Bassman和Zwier[8]的方法拟合Pn-PPFD的曲线方程,并计算最大光合速率(the maximum net photosynthetic rate,Pmax)、光饱和点(light saturati on point, LSP)、光补偿点(light compensation point, LCP)、表观量子效率(apparent quantum yield, AQY)和暗呼吸速率(dark respiration rate, Rd).
1.2.4 叶绿素含量测定 叶绿素含量参照Arnon[9]的方法测定.
1.2.5 酶活性测定 超氧化物歧化酶(SOD)活性参考李合生[10]的方法测定,过氧化物酶(POD)活性参考张志良等[11]的方法测定.
1.2.6 数据统计 数据用SPSS 11.5 One Way ANOVA进行统计分析,对同一测定指标在不同处理间的差异进行方差分析,并用LSD法检验各处理与对照组的差异显著性.
2 结果与分析
2.1 酸雨胁迫对3种草本植物叶片的伤害作用
随着酸雨pH的降低,3种植物受伤害程度逐渐加深且受害面积逐渐增大(表1). 其中,重度酸雨(pH 2.5)下,华泽兰和鸡矢藤叶片受害明显,受害叶面积约5%~10%左右,而蟛蜞菊叶片受害最轻,受害叶面积<5%,说明华泽兰和鸡矢藤对酸雨胁迫的敏感性更高,而蟛蜞菊对酸雨胁迫的耐受性更强.
表1酸雨胁迫下3种草本植物叶片的伤害程度
Table 1 Injury degree of leaves treated with simulated acid rain at various pH values
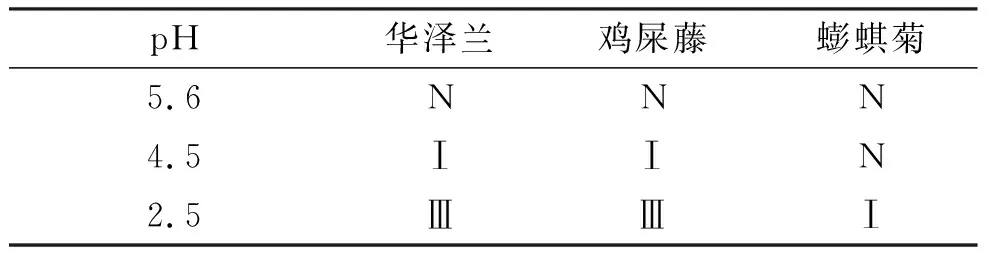
pH华泽兰鸡屎藤蟛蜞菊5.6NNN4.5ⅠⅠN2.5ⅢⅢⅠ
注: N: 叶片没有可见伤害; Ⅰ: 叶片受害轻微,受害叶面积<5%; Ⅱ: 叶片受害较明显,受害面积约为5%~10%; Ⅲ: 叶片受害明显,受害面积约为10% 左右; Ⅳ: 叶片受害较重,受害面积约为10%~20%.
2.2 酸雨胁迫对3种草本植物光合特性的影响
酸雨胁迫对3种草本植物叶片净光合速率(Pn)-光强响应曲线的影响不同(图1):重度酸雨(pH 2.5)条件下,当光合有效辐射大于200 μmol/(m2·s)时,华泽兰(图1A)和鸡矢藤(图1B)的Pn与对照相比明显降低,表明重度酸雨胁迫抑制了这2种植物的光合作用;相反,蟛蜞菊(图1C)的Pn在重度酸雨条件下与对照相比则明显升高,说明本实验中随着酸雨pH值的降低,蟛蜞菊的Pn有升高的趋势.
重度酸雨胁迫(pH 2.5)条件下,华泽兰和鸡矢藤的最大净光合速率(Pmax)与对照相比均显著下降(表2),相反,蟛蜞菊的Pmax与对照相比显著升高. 华泽兰和鸡矢藤的光饱和点(LSP)在各酸雨浓度条件下没有显著性变化、而光补偿点(LCP)逐渐升高,说明酸雨胁迫降低了华泽兰和鸡矢藤的光适应范围,而蟛蜞菊的LSP随酸雨浓度的降低而逐渐上升,但其LCP在各酸雨浓度条件下没有显著性变化,其光适应范围仍随酸雨浓度的降低而呈逐渐上升的趋势. 此外,中度(pH 4.5)和重度酸雨还显著增加了华泽兰的暗呼吸速率(Rd)(P<0.05).
华泽兰叶片叶绿素a+b含量随着酸雨浓度的降低而逐渐降低(表3),鸡矢藤叶片叶绿素a+b含量在各酸雨浓度下差异没有统计学意义,而蟛蜞菊叶片叶绿素a+b含量则随着酸雨浓度的降低反而呈现逐渐上升的趋势.此外,3种植物叶片叶绿素a/b在各酸雨浓度下有不同的差异,说明受试植物体内叶绿素a和叶绿素b对酸雨胁迫的敏感性不同.
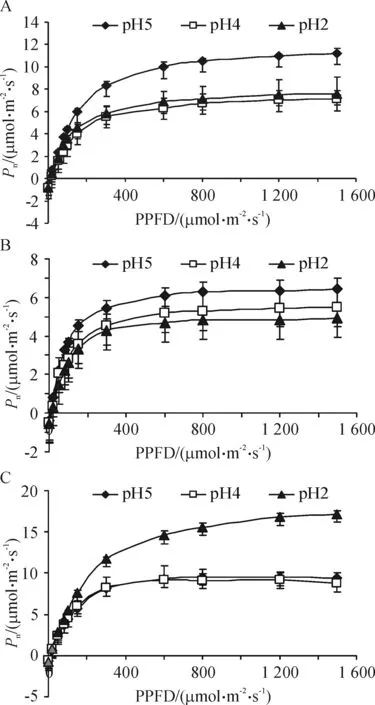
图1 不同酸雨处理下华泽兰(A)、鸡矢藤(B)和蟛蜞菊(C)的光响应曲线(平均值±标准误差n=4)
Figure 1 The light response curves ofEupatoriumchinense(A),Paederiascandens(B) andWedeliachinensis(C) under different treatments of simulated acid rain (mean±SEn=4)

表2 酸雨胁迫对3种草本植物光合特性的影响(平均值±标准误,n=4)
注:表中同一参数同一物种后相同字母表示在0.05水平上经LSD检验差异没有统计学意义(下同).
2.3 酸雨胁迫对3种草本植物叶片SOD和POD酶活性的影响
模拟酸雨处理后,3种植物叶片的SOD活性和POD活性均有不同程度的上升(图2). 重度酸雨处理下,鸡矢藤叶片SOD活性和POD活性与对照相比分别上升了236.78%和875.28%,且与对照的差异均有统计学意义(P<0.01),说明酸雨胁迫强烈的激发了鸡矢藤抗氧化酶系统,鸡矢藤具有较强的抗氧化胁迫能力.
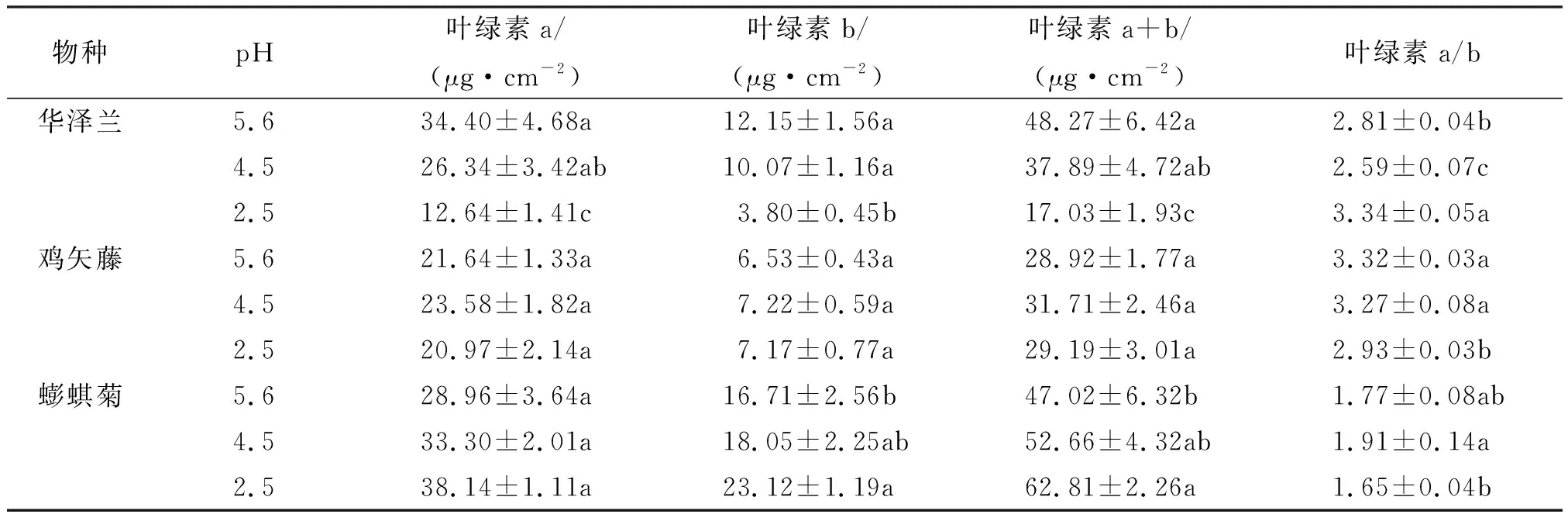
表3 酸雨胁迫对3种草本植物叶片叶绿素含量的影响(平均值±标准误,n=5)Table 3 Effects of simulated acid rain on chlorophyll contents of three herbaceous plants leaves(mean±SE, n=5)
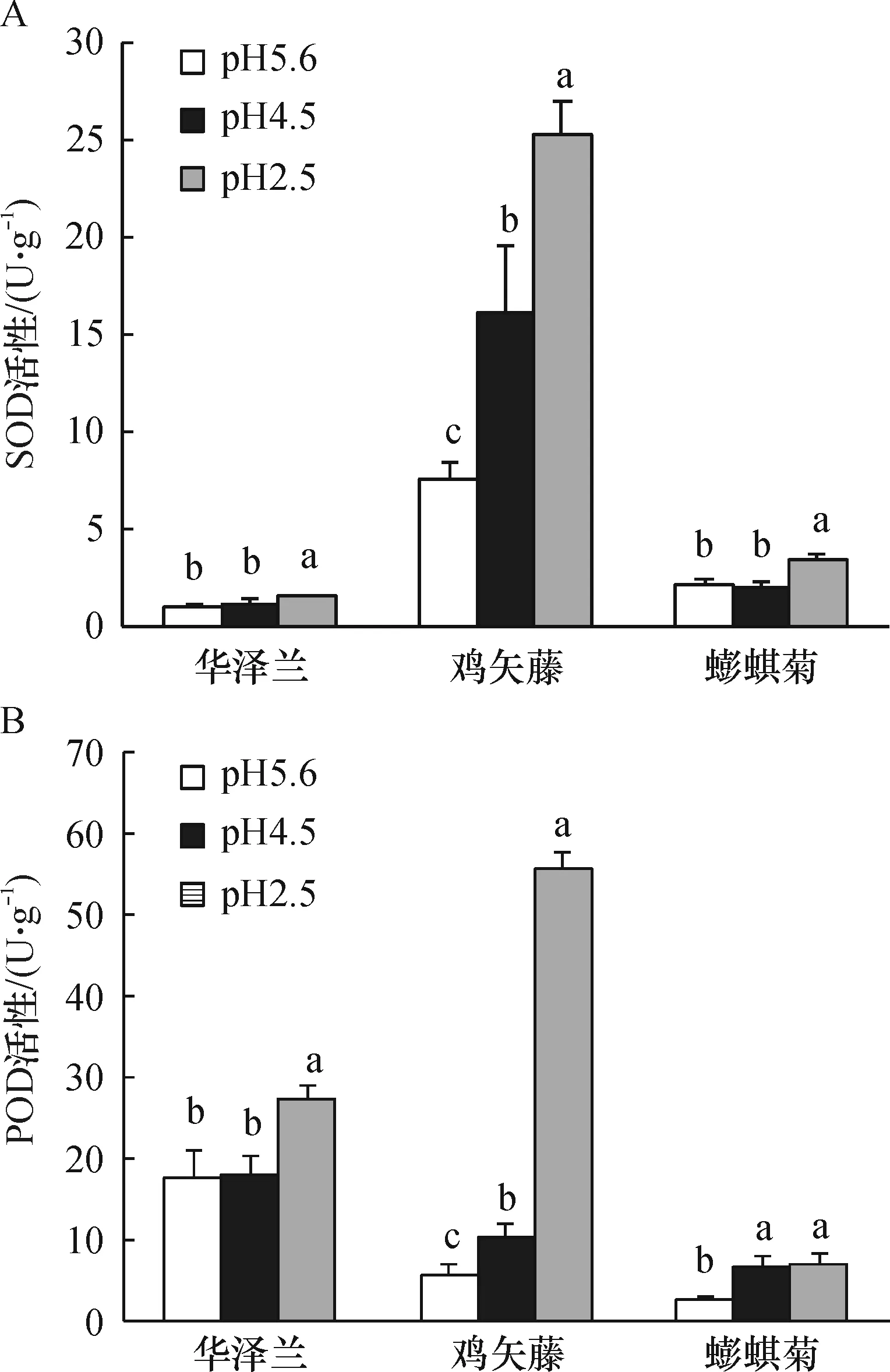
图2 酸雨胁迫对3种草本植物叶片SOD(A)、POD(B)活性的影响 (平均值±标准误差n=5)
Figure 2 Effects of simulated acid on activities of SOD (A) and POD (B) of three herbaceous plants leaves (mean±SEn=5)
3 讨论
研究表明酸雨会对植物的叶片结构产生伤害,使植物叶片出现黑斑和枯斑,严重时导致植物叶片坏死等[12]. 造成这种危害影响的原因可能是酸雨水附着于叶面时,损伤了叶表皮,破坏了细胞膜的结构与功能,造成细胞内电解质大量外渗,最终形成小的坏死斑[13]. 试验中蟛蜞菊叶片受酸雨胁迫的影响小于华泽兰和薇苷菊(表1),可能原因是蟛蜞菊叶片两面密被伏毛[14],酸雨无法直接通过叶片气孔进入叶肉细胞,进而减少了酸雨对其本身的伤害作用.
当酸雨突破植物叶片表层的防御(如角质层和蜡质层)后,能够通过植物叶片气孔或表皮扩散进入到植物体内进而影响植物正常的生长发育过程,这种影响存在一定的复杂性.如有研究报道酸雨能够降低髯毛箬竹(Indocalamusbarbatus)叶片叶绿素含量并抑制其光合速率[15],也有研究得到相反的结果[16]. 造成这种差异的原因,可能与植物光合色素含量的变化和光合机构的稳定性有关[17]. 本试验表明蟛蜞菊光合结构对酸雨的耐受能力较华泽兰和鸡矢藤更强(表2、表3),因此酸雨胁迫下仍能维持较高的光合作用. 此外,中度和重度酸雨显著增加了华泽兰的暗呼吸速率(P<0.05),可能是酸雨破坏了叶片线粒体的结果[17].
酸雨与植物互作是个复杂的过程,一方面酸雨破坏植物叶片表皮和细胞膜结构,一方面植物在受到酸雨胁迫时,体内的抗氧化酶系统受到激活,进而清除体内由于酸雨胁迫引起的过量的活性氧(ROS). 模拟酸雨处理后3种植物叶片的SOD活性和POD活性均有不同程度的上升,可能是参与抵抗酸雨胁迫过程的调节的结果.
综上所述,蟛蜞菊对酸雨的敏感程度较华泽兰和鸡矢藤更低,而华泽兰和鸡矢藤虽然对酸雨胁迫表现出一定的抵抗能力,但随着酸雨酸度增强,其抵抗能力逐渐减弱. 生产应用上,在酸雨污染严重的地方,如果要用草本植物进行绿化或生态恢复,建议种植抗性较强的蟛蜞菊等本地草本植物.
参考文献:
[1] 樊后保. 世界酸雨研究概况[J]. 福建林学院学报,2002,22(4):371-375.
Fan H B. On worldwide acid rain research[J]. Journal of Fujian College of Forestry,2002,22(4):371-375.
[2] Bini C,Bresolin F.Soil acidification by acid rain in forest ecosystems:a case study in northern Italy[J].Science of the Total Environment,1998,222(1/2):1-15.
[3] 梁钻姬,潘超美,赖珍珍,等. 药用植物华泽兰组织培养和快速繁殖[J]. 植物生理学报,2012, 48 (1): 85-89.
Liang Z J, Pan C M, Lai Z Z, et al. Tissue culture and rapid propagation of medicinal plantEupatoriumchinenseL.[J]. Plant Physiology Journal,2012,48(1):85-89.
[4] 余勉. 鸡矢藤化学成分及药理作用研究进展[J]. 卫生职业教育,2009,27(20):149-150.
Yu M. Advances in research of chemical constituents and pharmacological activities ofPaedenascanden[J]. Health Vocational Education,2009,27(20):85-89.
[5] 刘漫宇,朱家勇,金小宝. 蟛蜞菊活性成分的药理学研究进展[J]. 中国药房,2011,22(11):1048-1050.
Liu M Y, Zhu J Y, Jin X B. Advances in research of pharmacological activity ofWedeliachinensis[J]. China Pharmacy,2011,22(11):1048-1050.
[6] 刘大为,夏昊. 佛山市大气降水化学特征及其与广州市的比较[J]. 环境科学与技术,2007,30(增刊): 71-74.
Liu D W, Xia H.Chemical characteristics of precipitation in Foshan: comparing with Guangzhou[J]. Environmental Science and Technology,2007,30(supplement):71-74.
[7] 宋莉英,柯展鸿,孙兰兰,等. 模拟酸雨对3种菊科入侵植物光合特性的影响[J].植物学报,2013,48 (2):160-167.
Song L Y, Ke Z H, Sun L L, et al. Effect of simulated acid rain on gas exchanges of three compositae invasive plants[J]. Chinese Bulletin of Botany,2013,48(2):160-167.
[8] Bassman J,Zwier J C.Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltoidsandPopulustrichocarpa×P.deltoidsclone[J].Tree Physiology,1991,8(2):145-159.
[9] Arnon D. Copper enzymes in isolated chloroplasts polyphenol oxidase inbetavulgaris[J]. Plant Physiology,1949,24(1):1-15.
[10] 李合生. 植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:167-169.
[11] 张志良,翟伟菁. 植物生理学实验指导[M]. 北京:高等教育出版社,2003:123-126.
[12] 单运峰,冯宗炜,陈楚莹. 模拟酸雨对七种森林植物生物量的影响[J].生态学报,1989,9(3):274-276.
Shan Y F, Feng Z W,Chen C Y. The effects of simulated acid rain on biomass of seven species of forest plants[J]. Acta Ecologic Asinica,1989,9(3):274-276.
[13] 肖艳,黄建昌,刘伟坚,等. 12种园林植物对模拟酸雨的敏感性反应[J].广东园林,2004,26(3):651-655.
Xiao Y, Huang J C, Liu W J, et al. Sensitivity of twelve ornamental plant exposed to simulated acid rain[J]. Guangdong Landscape Architecture,2004,26(3):651-655.
[14] 聂呈荣,黎华寿,黄京华,等. 蟛蜞菊对花生和其它作物的化感作用[J]. 花生学报,2002,31(1):30-32.
Nie C R, Li H S, Huang J H, et al. The allelopathic effects ofWedeliaChinensisMerr. on peanut(ArachishypogaeaL.) and other crops[J]. Journal of Peanut Science,2002,31(1):30-32.
[15] 谢寅峰,杨万红,陆美蓉,等. 模拟酸雨胁迫下硅对髯毛箬竹光合特性的影响[J]. 应用生态学报,2008,19(6):1179-1184.
Xie Y F, Yang W H, Lu M R, et al. Effects of silicon on photosynthetic characteristics ofIndocalamusbarbatusunder simulated acid rain stress[J]. Chinese Journal of Applied Ecology,2008,19(6):1179-1184.
[16] Ferenbaugh R W. Effects of simulated acid rain onPhaseolusVulgarisL.(Fabaceae)[J]. American Journal of Botany,1976,63(3):283-288.
[17] 王玮. 模拟酸雨处理的青菜显微和亚显微结构观察及部分生理指标测定[J]. 环境科学,1988,9(3):12-17.
Wang W. Microstructure and Ultrastructure observation and some physiological indexes determination ofBrassicacompestristreated by the simulated acid rain[J].Environmental Science, 1988,9(3):12-17.
