几种杀虫剂对分月扇舟蛾离体酶活性的影响1)
2014-08-08刘晓露古丽吉米拉米吉提王志英吴红渠
刘晓露 古丽吉米拉·米吉提 王志英 吴红渠
(东北林业大学,哈尔滨,150040) (湖北生物农药工程研究中心)
几种杀虫剂对分月扇舟蛾离体酶活性的影响1)
刘晓露 古丽吉米拉·米吉提 王志英 吴红渠
(东北林业大学,哈尔滨,150040) (湖北生物农药工程研究中心)
以分月扇舟蛾(Closteraanastomosis)3龄幼虫为研究对象,比较了几种杀虫剂对其离体保护酶(SOD和CAT)和解毒酶(CarE和GST)活性的影响。结果表明:0.100 mg·L-1高效氯氟氰菊酯、0.010 mg·L-1吡虫啉、0.001 mg·L-1灭幼脲对CAT活性的抑制作用较强,抑制率分别为63.50%、68.89%和62.11%,而敌敌畏、氧化乐果对CAT活性的抑制作用较小。0.100 mg·L-1高效氯氟氰菊酯对SOD活性的抑制作用最强,抑制率为60.82%,氧化乐果对SOD活性有较强的促进作用,激活率可达168.72%(10.000 mg·L-1)。10.000 mg·L-1敌敌畏对CarE的抑制作用最强,抑制率为52.63%,1.000 mg·L-1高效氯氟氰菊酯对CarE的激活作用最强,激活率为43.06%。供试杀虫剂对GST活性均有抑制作用,1.000 mg·L-1灭幼脲对GST抑制率可达51.75%。由此可见,离体保护酶和解毒酶的活性与杀虫剂的类型有直接关系,并且同一类型不同化学结构的杀虫剂对离体酶活性的影响存在显著差异。
分月扇舟蛾;过氧化氢酶;超氧化物歧化酶;羧酸酯酶;谷胱甘肽-S-转移酶
We studied the effects of several pesticides on the activities of SOD, CAT, CarE and GST in three instars ofClosteraanastomosis. The activities of CAT were highly inhibited by 0.1 mg·L-1lambda-cyhalothrin, 0.010 mg·L-1imidacloprid, and 0.001 mg·L-1diflubenzuron, and the inhabitation rate were 63.50%, 68.89% and 62.11%, respectively. However, the activities of CAT were lowest inhibited by dichlorvos and omethoate, and highest inhibited by 0.1 mg·L-1lambda-cyhalothrin. The inhabitation rate of 0.100 mg·L-1lambda-cyhalothrin was 60.82%. The activities of SOD were highly increased by 10.000 mg·L-1omethoate, and the increasing rate was 168.72%. The activities of CarE were highly inhibited by 10.000 mg·L-1dichlorvos, and the increasing rate was 52.63%. The activities of CarE was highest increased by 1.000 mg·L-1lambda-cyhalothrin, and the increasing rate of 1.000 mg·L-1lambda-cyhalothrin was 43.06%. The activities of GST were all inhibited by the tested pesticides and the inhabitation rate of 1.000 mg·L-1lambda-cyhalothrin reached 51.75%. Thus, the activities of protective and detoxification enzymes have direct relation to the type of pesticides, and the effects on the activities of protective and detoxification enzymes are significant different by the same type of pesticides with different chemical structures.
杨树(Populusspp.)是优质速生的树种,在三北防护林和农田防护林建设中广泛栽植。随着杨树纯林的栽植,各种病虫害发生较为严重。分月扇舟蛾(Closteraanastomosis)是近年来严重危害杨树的重要森林害虫之一[1-2]。杨树被其危害后叶片枯黄,严重者树叶所剩无几,不仅影响当年枝条的木质化,而且严重影响翌年树木的生长发育,甚至导致树木死亡[3]。目前防治分月扇舟蛾的有效手段依旧是化学防治[4],造成了严重的环境污染和昆虫抗药性。羧酸酯酶(CarE)和谷胱甘肽-S-转移酶(GST)是昆虫体内重要的解毒酶,在分月扇舟蛾对化学药剂的解毒代谢中起着非常重要的作用。大量研究表明,超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)等与昆虫耐药性有关[5-6]。正常情况下,机体通过体内的保护酶系维持自由基的代谢平衡。当昆虫受到农药胁迫之后,昆虫体内会诱发细胞大量产生各种活性氧物质,而活性氧会导致体内的细胞膜脂质过氧化、影响细胞内的信号传导和基因表达等多种生理活动[7-8]。本研究以分月扇舟蛾为研究对象,通过探究杀虫剂对其CAT、SOD、CarE和GST活性的影响,探索害虫接触杀虫剂后的抗药性机制和解毒机理,为杀虫剂的管理、合理使用和安全性评价提供基础资料和理论指导。
1 材料与方法
1.1 供试虫源
于2013年6月,在东北林业大学示范林场采集大小一致、发育状态相近的3龄分月扇舟蛾幼虫。饥饿处理12 h,挑选健壮、活泼的幼虫,用液氮速冻后于冰箱中-80 ℃保存,用于杀虫剂对分月扇舟蛾离体酶活性抑制的测定。
1.2 供试化学试剂和杀虫剂
α-乙酸萘酯(α-NA)、毒扁豆碱、固蓝B盐、十二烷基硫酸钠(SDS)、乙二胺四乙酸二钠盐(EDTA-Na2)、二硫代苏糖醇(DTT)、还原型谷胱甘肽(GSH)、1-氯-2,4二硝基苯(DCNB)、核黄素(VB2)、苯基硫脲(N-Phenylthiourea)、聚乙烯吡咯烷酮(PVP)、氮蓝四唑(NBT)、甲硫氨酸(Met)等均购自Sigma公司;磷酸氢二钠(Na2HPO4)、磷酸二氢钠(NaH2PO4)、过氧化氢(H2O2),均购自天津永大化学试剂有限公司,均为分析纯。93.50%敌敌畏原药,购自郑州沙隆达伟新农药有限公司;82.8%氧化乐果原油,购自郑州沙隆达伟新农药有限公司;93.8%吡虫啉原药、97.0%杀铃脲原药、95.0%百树菊酯原药、94.3%灭幼脲原药、96.8%阿维菌素原药均购自吉林通化农药化工股份有限公司;95.0%高效氯氟氰菊酯原药,购自黑龙江省平山林业制药厂。
2 研究方法
2.1 杀虫剂配制
杀虫剂原药用丙酮溶解后配制成质量浓度为10 g·L-1的母液,然后用蒸馏水稀释并配制成质量浓度为10.000、1.000、0.100、0.010、0.001 mg·L-15个梯度。
2.2 酶液制备
取6头3龄分月扇舟蛾幼虫,清除体内食物等杂质,加4 mL磷酸缓冲液,冰浴匀浆后于4 ℃,12 000 r·min-1离心15 min,2次,上清液即为原酶液,于冰箱中-80 ℃贮存,备用。其中,提取CarE的PBS为0.04 mol·L-1,pH=7.0;提取GST的PBS为0.10 mol·L-1,pH=6.5(含1 mmol·L-1EDTA-Na2,0.1 mmol·L-1DTT);提取CAT和SOD的PBS为0.05 mol·L-1,pH=7.0(含1% PVP,0.04%苯基硫脲,10 mmol·L-1EDTA)。
2.3 酶活性测定
CAT活性的测定:参照Cohen等[9]方法进行测定。取30% H2O20.5~0.6 mL,加水至50 mL,从中取出4 mL,加入26 mL 0.05 mol·L-1pH=7.0的PBS,测定240 nm的OD值,OD值为0.50~0.55时,即作为过氧化氢酶的底物溶液。在石英比色皿中依次加入3 mL底物溶液,20 μL稀释酶液,立即测定240 nm处的OD值,每隔30 s读一次,记录3 min,试验重复3次。以OD240减少0.1的酶量为一个酶活性单位。
CAT活性=(ΔA×V)/(0.1×T×VE)。
式中:ΔA为每分钟OD240的减小值;V为反应总体积;T为加过氧化氢到最后一次读数时间;VE为酶液体积。
SOD活性的测定:参照Beauchamp等方法[10]进行测定。在试管中依次加入3 mL反应液(含50 mmol·L-1pH=7.8的PBS,13 mmol·L-1Met,0.1 mmol·L-1EDTA,75 μmol·L-1NBT),25 μL稀释酶液,0.6 mL 0.5 mmol·L-1核黄素,以不加酶液管作为最大光还原管,4 000 lx下光照15 min后,立即避光迅速测定OD560,以未光照的相同反应管为对照管,重复3次。以达到50%抑制率所需的酶量为一个酶活性单位。
SOD活性=(A×V×60)/(T×VE×50%)。
式中:A为最大光还原管OD560-反应管OD560;V为反应总体积;VE为酶液体积;T为反应时间。
CarE活性的测定:参照Ibrahim等[11]方法进行测定。在试管中依次加入0.1 mL稀释酶液,2 mL 3×10-4mol·L-1α-NA(含毒扁豆碱1∶1)丙酮溶液,35 ℃水浴10 min后,加入1 mL显色剂(V(1%固蓝B盐)∶V(5% SDS)=2∶5)终止反应,以PBS代替酶液作为空白对照,测其在600 nm下的吸光值,重复3次。
CarE活力=(ΔOD600×V)/(ε×L)。
式中:ΔOD600为每分钟光吸收值的变化值;V为酶促反应总体积;ε为产物的消光系数(13.6 L·mmol-1·cm-1);L为比色杯的光程(1 cm)。
GST活性的测定:参照Habig等[12]方法进行测定。在石英比色皿中依次加入0.1 mL稀释酶液,2.3 mL 0.1 mol·L-1pH=6.5的PBS,0.5 mL 5 mmol·L-1还原型谷胱甘肽(GSH)和0.1 mL 1.0 mmol·L-1CDNB,以PBS代替酶液作为对照。记录其在340 nm下3 min内的吸光值变化,每隔30 s记录一次,重复3次。
GST活力=(ΔOD340×V)/(ε×L)。
式中:ΔOD340为每分钟光吸收值的变化值;V为酶促反应总体积;ε为产物的消光系数(9.6 L·mmol-1·cm-1);L为比色杯的光程(1 cm)。
2.4 杀虫剂对分月扇舟蛾活性的抑制
离体CAT活性抑制的测定:分别将不同质量浓度的8种杀虫剂与稀释酶液混合,总反应体系为3.12 mL。将0.1 mL不同质量浓度杀虫剂与20 μL稀释酶液混合均匀,于35 ℃温浴5 min后取出,加入3 mL底物溶液,测定残余活性。每种杀虫剂的每个质量浓度3次重复。
离体SOD活性抑制的测定:分别将不同质量浓度的8种杀虫剂与稀释酶液混合,总反应体系为3.95 mL。将0.1 mL不同质量浓度杀虫剂与25 μL稀释酶液混合均匀,于35 ℃温浴5 min后取出,在试管中依次加入3 mL反应液,与农药反应后的酶液,0.6 mL 0.5 mmol·L-1核黄素,测定残余活性。每种杀虫剂的每个质量浓度3次重复。
离体CarE活性抑制的测定:分别将不同质量浓度的8种杀虫剂与稀释酶液混合,总反应体系为3.2 mL。将0.1 mL不同质量浓度杀虫剂与0.1 mL稀释酶液混合均匀,于35 ℃温浴5 min,加入底物2 mL,35 ℃温浴10 min后取出,加显色剂1 mL终止反应,测定残余活性。每种杀虫剂的每个质量浓度3次重复。
离体GST活性抑制的测定:分别将不同质量浓度的8种杀虫剂与稀释酶液混合,总反应体系为3.1 mL。将0.1 mL不同质量浓度杀虫剂与0.1 mL稀释酶液混合均匀,于35 ℃温浴5 min,加入2.3 mL 0.1 mol·L-1pH=6.5的PBS,0.5 mL 5 mmol·L-1GSH和0.1 mL 45 mmol·L-1CDNB,测定残余活性。每种杀虫剂的每个质量浓度3次重复。
2.5 数据处理
采用SPSS16.0统计软件进行方差分析,采用Duncan’s新复极差法比较各处理间的差异显著性(p<0.05);试验数据用平均值±标准误表示,酶活抑制率的计算公式为:酶活性抑制率=((对照组酶活性-残留酶活性)/对照组酶活性)×100%。
3 结果与分析
3.1 杀虫剂对分月扇舟蛾CAT体外活性的影响
8种杀虫剂对分月扇舟蛾3龄幼虫CAT活性的影响见表1。高效氯氟氰菊酯、吡虫啉、杀铃脲、灭幼脲、阿维菌素对分月扇舟蛾CAT活性都有一定的抑制作用,其中高效氯氟氰菊酯、吡虫啉、灭幼脲对CAT活性有较强的抑制作用,抑制率分别为41.91%~63.50%、37.31%~68.89%、37.51%~62.11%。高效氯氟氰菊酯处理组对CAT活性的抑制作用表现为:随质量浓度增加抑制率呈先升后降的趋势,在0.100 mg·L-1抑制率达到最大,为63.50%;吡虫啉处理组离体酶活性的抑制率表现为低质量浓度(0.010 mg·L-1)强抑制,而在0.100、1.000、10.000 mg·L-1处理时,其梯度间抑制作用差异不显著(p>0.05);杀铃脲处理组对分月扇舟蛾CAT酶活性影响表现出随质量浓度的升高(除0.100 mg·L-1外),抑制作用逐渐增强,当高剂量(>1.000 mg·L-1)处理时,抑制率超过40.00%,而在低质量浓度时,抑制率较低,与高剂量间差异显著(p<0.05);农用抗生素阿维菌素对CAT活性也有一定的影响,但抑制作用较弱。而百树菊酯对CAT活性的抑制作用不大,抑制率为8.87%~18.33%;敌敌畏、氧化乐果甚至有时起到低激活的作用(敌敌畏1.000 mg·L-1、氧化乐果10.000 mg·L-1)。在供试的8种杀虫剂中,仅杀铃脲处理组对CAT酶活性抑制作用与质量浓度呈正相关,其余7种药剂处理后,对分月扇舟蛾CAT酶活性的影响并未表现出与质量浓度的相关性。
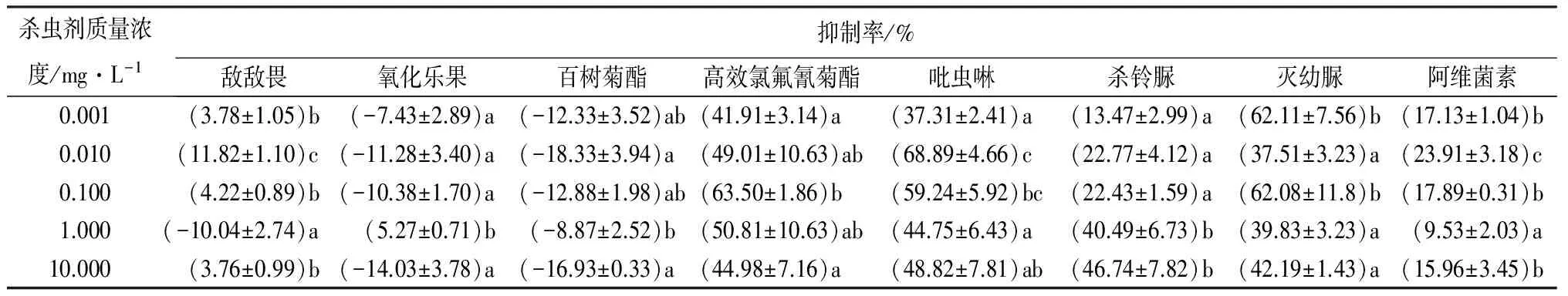
表1 杀虫剂对分月扇舟蛾3龄幼虫离体CAT活性的影响
注:表中数据为平均值±标准差;同列数字后小写字母不同表示不同处理间差异显著(p<0.05)。
3.2 杀虫剂对分月扇舟蛾SOD体外活性的影响
8种杀虫剂对分月扇舟蛾3龄幼虫SOD活性的影响见表2。敌敌畏、高效氯氟氰菊酯、百树菊酯、阿维菌素对分月扇舟蛾3龄幼虫SOD活性均有一定的抑制作用,其最高抑制率分别达43.12%、60.82%、41.81%、24.31%,其中敌敌畏、百树菊酯对SOD活性表现为较强的抑制作用,抑制率均在28.00%以上,其抑制率的高低与质量浓度的大小无关;而高效氯氟氰菊酯0.100、1.000 mg·L-1的抑菌率分别达60.82%、55.17%,其余各质量浓度抑制率较低,对离体SOD酶活性的抑制与质量浓度无必然关系;生物药剂阿维菌素总体来讲,对离体SOD酶活性的抑制率维持在较低的水平,当质量浓度高于0.010 mg·L-1时,抑制率与药剂质量浓度呈正相关,在中高质量浓度处理下(>1.000 mg·L-1),抑制率超过了22.28%,但差异不显著(p>0.05)。几丁质合成抑制剂杀铃脲对分月扇舟蛾3龄幼虫离体SOD酶活性的作用表现为:先抑制(<0.100 mg·L-1)再促进(1.000 mg·L-1)再抑制的变化趋势,当10.000 mg·L-1处理时,抑制率达最高。氧化乐果、高质量浓度的吡虫啉(>0.010 mg·L-1)、高质量浓度的灭幼脲(10.000 mg·L-1)对SOD活性均有促进作用。其中,氧化乐果和吡虫啉随质量浓度的升高对离体酶的促进作用呈正相关,且同种药剂不同质量浓度处理间差异显著(p<0.05)。
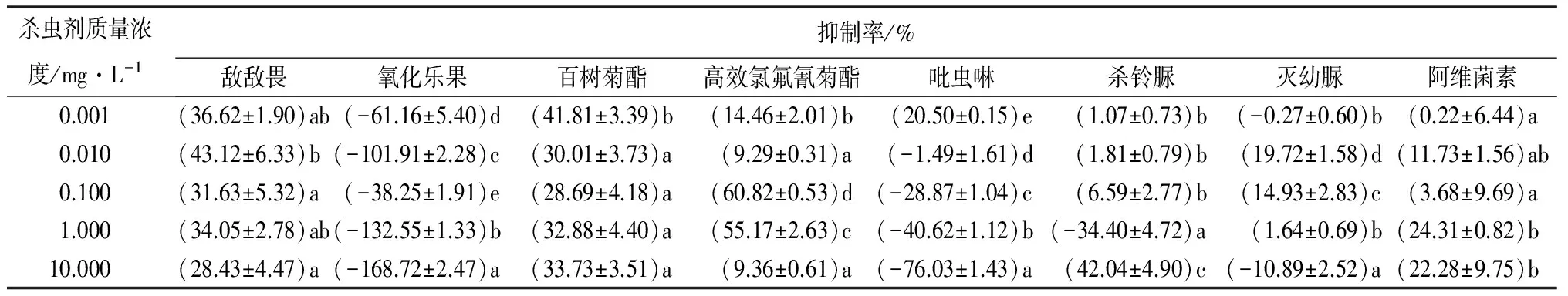
表2 杀虫剂对分月扇舟蛾3龄幼虫离体SOD活性的影响
注:表中数据为平均值±标准差;同列数字后小写字母不同表示不同处理间差异显著(p<0.05)。
3.3 杀虫剂对分月扇舟蛾CarE活性的影响
由表3可知,8种杀虫剂对分月扇舟蛾3龄幼虫离体CarE的活性影响各不相同。阿维菌素表现为较强的抑制作用,抑制率为19.71%~31.65%;敌敌畏在低质量浓度表现为促进作用,随着质量浓度的升高,促进作用减弱,当质量浓度高于0.100 mg·L-1时,表现为抑制作用,且随着质量浓度的升高,抑制率逐渐增大,当10.000 mg·L-1时,抑制率达52.63%;而氧化乐果、百树菊酯、高效氯氟氰菊酯、吡虫啉、灭幼脲处理时,表现为不同程度的激活作用。低剂量的氧化乐果(<0.010 mg·L-1)随质量浓度的升高促进作用增强,当质量浓度高于0.010 mg·L-1时,促进作用减弱,10.000 mg·L-1处理组的促进率仅为1.55%,与处理组间差异显著(p<0.05);高效氯氟氰菊酯各处理组(除10.000 mg·L-1处理外)随质量浓度的升高,促进作用逐渐升高,10.000 mg·L-1处理组促进作用虽有所下降,但与1.000 mg·L-1差异不显著(p>0.05);百树菊酯与高效氯氟氰菊酯相似,促进作用随质量浓度的升高逐渐增强(除10.000 mg·L-1外);吡虫啉和中高质量浓度的百树菊酯(>0.100 mg·L-1)激活作用稳定,激活率分别为17.17%~29.03%、13.38%~22.82%;灭幼脲处理组,整体表现为激活作用,但随着质量浓度的升高,激活作用减弱,低质量浓度促进作用明显,当质量浓度大于0.100 mg·L-1仅有微弱的激活作用。
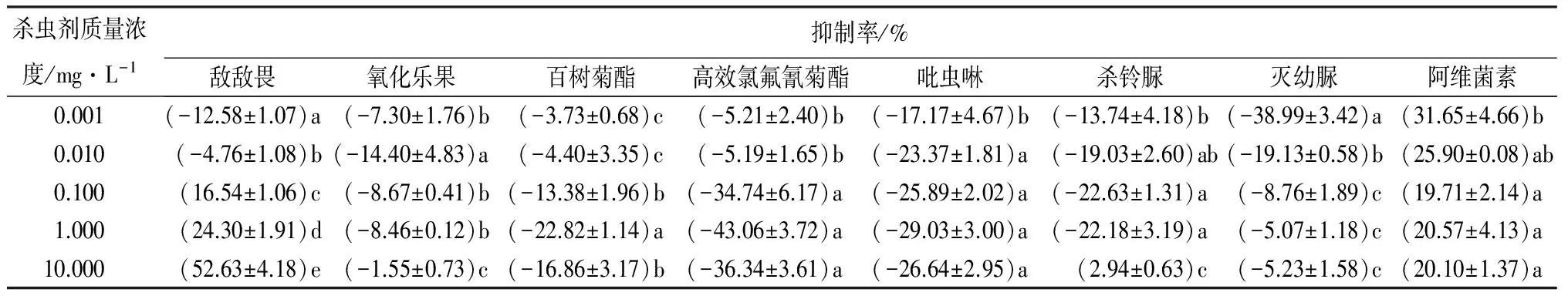
表3 杀虫剂对分月扇舟蛾3龄幼虫离体CarE活性的影响
注:表中数据为平均值±标准差;同列数字后小写字母不同表示不同处理间差异显著(p<0.05)。
3.4 杀虫剂对分月扇舟蛾GST活性的影响
表4显示,8种农药对分月扇舟蛾3龄幼虫离体GST活性均有抑制作用。其中,吡虫啉对GST活性影响表现出整体抑制作用较差,抑制率为2.82%~6.37%,抑制率呈先升高再下降的趋势,且不同质量浓度处理间差异不显著(p>0.05),敌敌畏、杀铃脲、阿维菌素对GST活性影响次之;低质量浓度(<1.000 mg·L-1)氧化乐果对GST活性具有较显著地抑制作用,大于1.000 mg·L-1时,抑制率明显下降,且与低质量浓度间差异显著(p<0.05)。高效氯氟氰菊酯和百树菊酯对GST活性的抑制作用随质量浓度的增加呈现先升高后降低的趋势,最高抑制率分别可达44.52%和38.25%。高质量浓度灭幼脲(>1.000 mg·L-1)对GST活性抑制作用较其他质量浓度处理差异显著(p<0.05),高达51.75%。
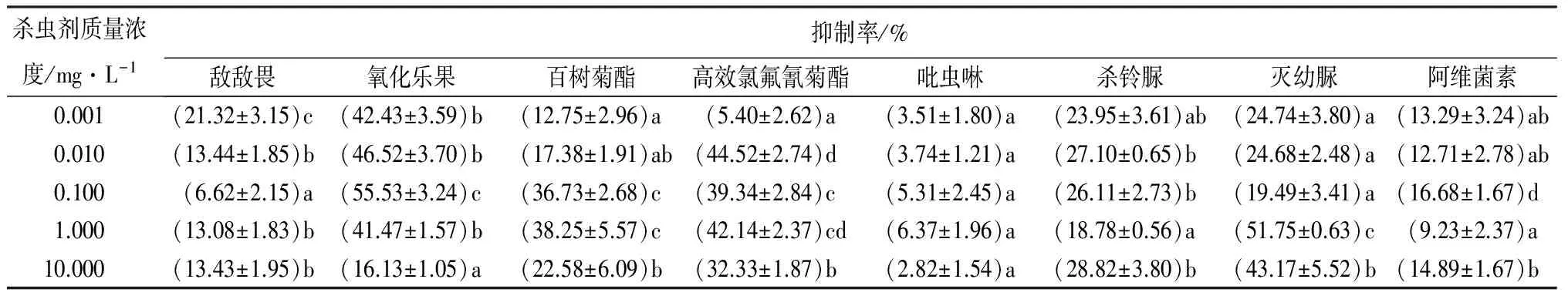
表4 杀虫剂对分月扇舟蛾3龄幼虫离体GST活性的影响
注:表中数据为平均值±标准差;同列数字后小写字母不同表示不同处理间差异显著(p<0.05)。
4 结论与讨论
离体酶在失去原有的生物膜保护系统时,无法进行自身的修复还原,其活性的抑制试验不仅对明确各种杀虫剂的杀虫机制及研发新的杀虫剂有着重要的作用,也为农药由仿制到创制的转变提供新思路,而且还对合理评价和利用杀虫剂具有重要的指导意义。本研究比较了8种杀虫剂原药对分月扇舟蛾3龄幼虫体外保护酶和解毒酶活性的影响,结果表明:分月扇舟蛾3龄幼虫体外保护酶和解毒酶的活性与杀虫剂的种类和质量浓度有着密切关系,且同类杀虫剂因化学结构不同,其对离体酶活性的影响也不尽相同,这为现阶段比较农药化合物构效关系数据库资料提供了切实的理论依据。
CAT和SOD是昆虫体内关键的保护酶,它们在昆虫生长发育、杀虫剂渗入和病原微生物侵染等情况下发挥着清除自由基、保护机体免受或减轻伤害的作用[13]。查黎春等[14]研究发现高效氯氰菊酯、阿维菌素和辛硫磷对天幕毛虫SOD离体活性有显著地影响。吕林兰等[15-16]研究表明,马拉硫磷对双齿围沙蚕(Perinereisaibuhitensis)体内的SOD和CAT活性具有显著影响。这些研究结果均表明,许多杀虫剂对昆虫体内的CAT和SOD活性有一定的影响作用。本研究发现,拟除虫菊酯类杀虫剂(高效氯氟氰菊酯)、烟碱类杀虫剂(吡虫啉)、几丁质合成抑制剂(杀铃脲)和苯甲酰基类杀虫剂(灭幼脲)对分月扇舟蛾3龄幼虫CAT活性的体外抑制作用最强,抑制率分别达63.50%(0.100 mg·L-1)、68.89%(0.010 mg·L-1)、46.74%(10.000 mg·L-1)和62.11%(0.001 mg·L-1)。有机磷类杀虫剂(敌敌畏、氧化乐果)对CAT活性的抑制作用不大,甚至有时起到激活的作用(敌敌畏0.100 mg·L-1、氧化乐果1.000 mg·L-1)。本试验所用的另一类拟除虫菊酯类杀虫剂百树菊酯对离体CAT活性有较弱的激活作用,它与高效氯氟氰菊酯所属同一类杀虫剂,但其官能团上的氯离子被氟离子取代,由于氟原子的引入,提高了其脂溶性和疏水性,这样可能促进其与CAT酶的活性部位结合。笔者推测,这是导致其对CAT酶活抑制效果较高效氯氟氰菊酯低的主要原因之一。百树菊酯和高效氯氟氰菊酯对分月扇舟蛾3龄幼虫SOD活性的体外抑制作用最强,抑制率分别达41.81%(0.001 mg·L-1)和60.82%(0.100 mg·L-1)而氧化乐果对SOD活性的体外激活作用最强,激活率为61.16%~168.72%,且激活率与氧化乐果的质量浓度呈正相关。由此可见,离体CAT和SOD活性与杀虫剂的类型有直接关系,并且同一类型不同化学结构的农药对离体CAT和SOD活性的影响存在显著差异。在供试杀虫剂中,敌敌畏、拟除虫菊酯类杀虫剂(百树菊酯和高效氯氟氰菊酯)和抗生素类杀虫剂(阿维菌素)对分月扇舟蛾3龄幼虫的2种保护酶具有良好的抑制效果,相对而言,氧化乐果对其抑制效果最差。
CarE和GST是昆虫体内重要的解毒酶,在杀虫剂代谢和抗药性的形成方面有着重要意义[17]。它们的活性能够被各种外源化合物诱导,使昆虫迅速作出反应,从而存活下来[18]。王旭等[19]和许雄山等[20]先后报道了敌敌畏等有机磷杀虫剂对棉铃虫离体CarE有抑制作用。汤方等[21]研究表明,槲皮素、单宁酸和辛硫磷对分月扇舟蛾GST有抑制作用,抑制中浓度分别为2.19×10-5,2.62×10-5,1.49×10-4mol·L-1。本研究发现,敌敌畏和阿维菌素对分月扇舟蛾3龄幼虫CarE活性的体外抑制作用最强,而百树菊酯和高效氯氟氰菊酯对CarE活性的体外有激活作用。氧化乐果、百树菊酯、高效氯氟氰菊酯和灭幼脲对分月扇舟蛾3龄幼虫GST活性的体外抑制作用最大,而吡虫啉对GST活性的体外抑制作用最弱。由此可见,在供试杀虫剂中,有机磷类、激素类和抗生素类杀虫剂对分月扇舟蛾3龄幼虫的两种解毒酶具有良好的抑制效果,烟碱类杀虫剂(吡虫啉)对其抑制效果最差。
[1] 方杰,崔永三,赵博光,等.我国分月扇舟蛾的研究进展[J].中国森林病虫,2007,26(2):28-31.
[2] 孟庆庭,翟光耀,马蓓莉.杨树主要病虫害及防治技术[J].绿色科技,2013(3):69-71.
[3] 杨振德,朱麟,赵博光,等.实验室群体饲养条件下分月扇舟蛾生物学特性初步研究[J].广西科学院学报,2004,20(1):35-37.
[4] 苏元吉,高玉梅.分月扇舟蛾发生特点及防治探析[J].林业科技情报,2009,41(3):8-9.
[5] 李周直,沈惠娟,蒋巧根,等.几种昆虫体内保护酶系统活力的研究[J].昆虫学报,1994,37(4):399-403.
[6] 马义,霍新北,马民,等.氯氟氰菊酯对德国小蠊保护酶活性的影响及新型防治策略研究[J].中国媒介生物学及控制,2009,20(4):303-306.
[7] Thannickal V J, Fanburg B L. Reactive oxygen species in cell signaling[J]. Am J Physiology Lung Cell Mol Physiology,2000,279(6):L1005-L1028.
[8] Morel Y, Barouki R. Repression of gene expression by oxidative stress[J]. Biochem J,1999,342(Pt3):481-496.
[9] Cohen G P, Dembiec D, Marcus J. Measurement of catalase activity in tissue extracts[J]. Analytical Biochemistry,1970,34(1):30-38.
[10] Beauchamp C, Fridovich I. Superoxide dismutase: Improved assays and assay applicable to acrylamidae gels[J]. Analytical Biochemistry,1971,44(1):276-287.
[11] Ibrahim H, Kheir R, Helmi S, et al. Effects of organophosphorus, carbamate, pyrethroid and organochlorine pesticides, and a heavy metal on survival and cholinesterase activity of Chironomus riparius Meigen[J]. Bulletin of Environmental Contamination and Toxicology,1998,60(3):448-455.
[12] Habig W H, Pabst M J, Jakoby W B. Glutathione S-transferase AA from rat liver[J]. Archives of Biochemistry and Biophysics,1976,175(2):710-716.
[13] 邹传山.落叶松毛虫酶学特性及生化毒理学研究[D].哈尔滨:东北林业大学,2013.
[14] 查黎春,胡春祥,曹传旺,等.3种农药对天幕毛虫超氧化物歧化酶活性的影响[J].东北林业大学学报,2011,39(5):105-107.
[15] 吕林兰,董学兴,黄金田,等.马拉硫磷对双齿围沙蚕抗氧化酶的影响[J].安徽农业科学,2010,38(11):5719-5722.
[16] 吕林兰,杨家新,董学兴,等.马拉硫磷对双齿围沙蚕乙酰胆碱酯酶和过氧化氢酶的影响[J].农业环境科学学报,2010,29(3):431-436.
[17] 蒋晴,苏宏华,杨益众.昆虫对Bt毒素的抗性与中肠蛋白酶、解毒酶及保护酶系活性的关系[J].环境昆虫学报,2013,35(1):95-101.
[18] 唐振华.我国昆虫抗药性研究的现状及展望[J].昆虫知识,2000,37(2):97-103.
[19] 王旭,高希武,郑炳宗,等.有机磷药剂对棉铃虫羧酸酯酶的抑制作用[J].昆虫学报,1998,41(S1):12-18.
[20] 许雄山,韩召军,王荫长.羧酸酯酶与棉铃虫对有机磷杀虫剂抗性的关系[J].南京农业大学学报,1999,22(4):41-44.
[21] 汤方,李丽,高希武.植物次生物质和杀虫剂对分月扇舟蛾谷胱甘肽S-转移酶的抑制作用[J].应用昆虫学报,2012,49(6):1513-1518.
Effects of Several Pesticides on the Activities ofinvitroEnzymes inClosteraanastomosis
/Liu Xiaolu, Gulijimila·Mijiti, Wang Zhiying(Northeast Forestry University, Harbin 150040, P. R. China); Wu Hongqu
(Hubei Biopesticide Engineering Research Center)//Journal of Northeast Forestry University.-2014,42(11).-128~133
Closteraanastomosis; Catalase; Superoxide dismutase; Carboxylesterase; Glutathione-S-transferase
刘晓露,女,1989年5月生,东北林业大学林学院,硕士研究生。
王志英,东北林业大学林学院,教授。E-mail:Zyw0451@sohu.com。
2014年3月24日。
S763.306.7;S763.42
1) “十二五”农村领域国家科技计划课题(2011BAD37B01-4);林业公益性行业科研专项(201104069)。
责任编辑:程 红。
