稳定同位素在植物水分来源及利用效率研究中的应用
2014-07-25吴骏恩刘文杰朱春景
吴骏恩 刘文杰 朱春景
(1.中国科学院西双版纳热带植物园热带森林生态学重点实验室, 云南 勐仑 666303;2.中国科学院大学, 北京 100049)
稳定同位素在植物水分来源及利用效率研究中的应用
吴骏恩1,2刘文杰1朱春景1,2
(1.中国科学院西双版纳热带植物园热带森林生态学重点实验室, 云南 勐仑 666303;2.中国科学院大学, 北京 100049)
介绍了稳定氢、氧、碳同位素技术在定量区分植物水分来源及利用效率研究中的应用。以往大量研究结果表明,由于植物根系在吸收水分过程中没有发生同位素分馏,所以通过比较和分析植物木质部水分及其可利用水源的氢氧同位素δ值,就可以定量判断植物的水源;植物在光合作用过程中会产生碳同位素的分馏,从而使得植物叶片中的碳同位素组成能够成为指示植物水分利用效率的一个间接指标,是目前研究植物叶片长期水分利用效率的最佳方法;稳定同位素技术作为安全的示踪剂,有助于人类了解植物的生理生态功能,随着该技术的不断完善和研究的不断深入,同位素技术将会在生态学研究的许多领域得到广泛的应用。
稳定同位素;植物;水分来源;利用效率
伴随着极端天气事件的增加,陆地生态系统对全球变化的响应已经成为当前科学研究的热点。全球变化的一个重要内容是区域降雨格局的变化,它对生态水分平衡与植被分布都产生了极为重要的影响[1],尤其是植物吸收和利用水分的模式。因此,定量研究植物的水分利用模式,不仅有助于了解由降雨格局变化所导致的植被时空变化规律[2],还能指导人们选择合适的植物进行植被建设与恢复工作。
传统研究植物水源的方法一直都很困难,如根系挖掘法,虽然可以确定植物可利用的水源,但无法判断其主要水源,因为根系的存在并不意味着这些根在水分吸收方面活跃,植物群落中的根系分布策略被认为是生态位分化的一种形式,所以根系分布仅代表水分的可利用性而非水分本身分布的结果[3];此外还有根据地表以下各土层土壤含水量变化以及植物体水分指标的季节变化研究水源的方法,这些方法受环境因素影响大,很难精确地判断植物的水源[4]。相比较而言,稳定同位素技术具有较高的灵敏性与准确性,是目前判断植物水源较为先进和准确的方法。
水分利用效率(WUE)是耦合植物光合与水分生理过程的重要指标,是指植被光合作用生产的干物质量与蒸散作用消耗的水量之比,是联系植被生态系统碳循环和水循环的关键因子,具有极为重要的生理学、生态学和水文学意义[5]。早前,国内外关于WUE的研究方法都比较直接和简单,常见的方法有田间测量法与气体交换法。其中田间测量法需要进行大量繁重的工作,且受人为因素影响太大;而气体交换法所测量的WUE为瞬时水分利用效率,只能反应植物在某一时刻的WUE,且受环境因素影响较大;相比之下,稳定同位素技术不仅可以克服传统方法中遇到的困难,更因其准确性,成为目前研究植物叶片长期水分利用效率的最佳方法。
1 稳定同位素技术发展概况
20世纪30年代,Urey等[6]发现了重氢(D),同时发现不同水体中重氢含量不同。然而,稳定同位素作为独特的示踪剂,在当时主要应用于岩石、生物、海洋、河流、地下水及各种矿床等领域的研究中[7]。直到20世纪50年代初,稳定同位素开始大量应用于水环境科学领域。从20世纪80年代到现在,这项技术被广泛地应用到植物水源的研究中,从植物个体到种群,再从种群到群落;从研究森林到各式各样的植物群落;从区分植物对雨水、土壤水、地表径流和地下水的利用到区分对雾水露水的利用。如今,稳定同位素技术已成为生态学研究的重要手段[8]。
2 稳定同位素值的表示方法
自然界中稳定同位素的含量极低,要用绝对丰度来表达同位素的差异是非常困难的,所以同位素含量通常用相对量来表示,即:
δX= (Rsample/Rstandard- 1)×1 000
( 1 )
式中:Rsample是样品中元素的重轻同位素丰度比(如D/H,16O/18O);Rstandard是国际通用标准物的重轻同位素丰度之比(氢、氧稳定同位素采用v-SMOW标准,碳稳定同位素采用v-PDB标准,如13Cstandard/12Cstandard)。
在植物水分利用策略的研究中,主要的可利用稳定同位素有碳、氢、氧,它们的主要来源分别是CO2和H2O,其组成参考标准如表1所示。
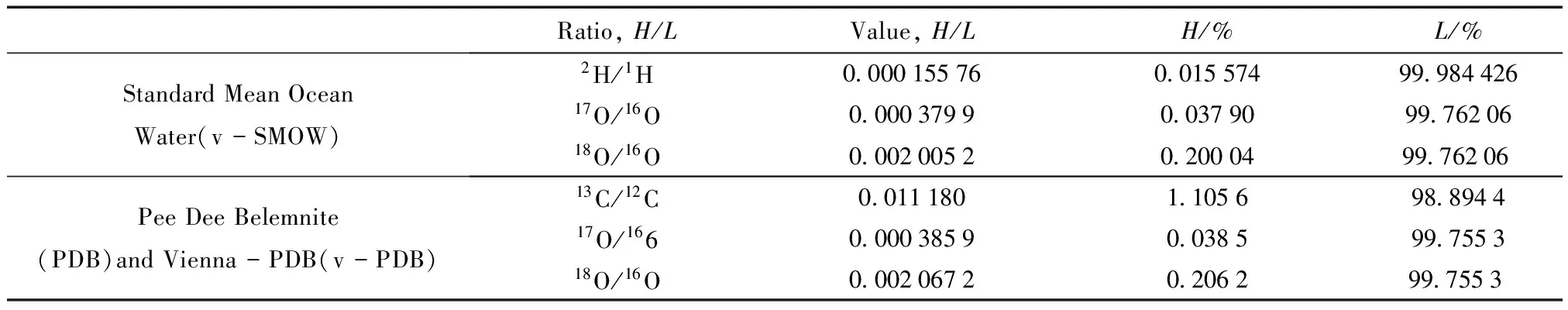
表1 同位素组成的国际参考标准[9]
注:H和L分别代表重同位素和轻同位素。
3 氢氧稳定同位素在定量判断植物水分来源中的应用
植物体内的水主要来自于大气降水、土壤水、地表径流和地下水等。由于水在循环过程中发生了包括蒸发、凝聚、降落、渗透等物理化学过程,从而引起了同位素的分馏[10-11],使得这些水体各自都具有不同的氢氧同位素值。然而植物根系在吸水过程中氢氧同位素是不会发生分馏的[12-13],即导管内水与根系吸收水的氢氧同位素在组成上是一致的。因此,通过分析植物木质部水和各水源的氢氧同位素组成就可以确定植物的水分来源。
3.1 氢氧稳定同位素研究植物水分来源的模型
3.1.1二项和三项线性混合模型 植物木质部水分的氢氧同位素值是多种水源氢氧同位素混合组合的结果。当植物有2种水源时,可用如下公式计算其贡献率[14]:
δD=f1δD1+f2δD2
(2)
δ18O =f1δ18O1+f2δ18O2
(3)
f1+f2=1
(4)
当存在3种水源时,计算公式依此类推:
δD =f1δD1+f2δD2+f3δD3
(5)
δ18O =f1δ18O1+f2δ18O2+f3δ18O3
(6)
f1+f2+f3=1
(7)
其中:δD (δ18O )由植物木质部水分的稳定氢(氧)同位素组成;δD1(δ18O1)、δD2(δ18O2)、δD3(δ18O3)为水源1、2、3的稳定氢(氧)同位素组成;f1、f2、f3为水源1、2、3在植物所利用的水分总量中所占的百分数,和值为1。
3.1.2IsoSource 模型 当植物可能利用的水源过多时,测定的同位素种类就不能满足线性混合模型的要求。据此,Robert Gibson公司开发了IsoSource模型[15]。该模型可以不受水源个数和测定同位素种类的限制,巩国丽等[16]曾用该模型研究格尔木白刺的水源,同时还将该模型与其他模型进行了对比。
运行模型前需要设置来源增量(source increment)和质量平衡公差(mass balance tolerance)2个参数[15,17],模型运行计算的结果是一个由最大值和最小值组成的范围值。2种模型都是计算植物各水源贡献率的常用方法,但需注意的是各可能水源之间同位素值差异越小,计算结果与实际情况的误差就越大。增加差异的方法有2种[18]:一种是人为地向各潜在水源添加不等量重水(D2O);另一种是增大各可能水源样品的数目。
3.2 植物水源的研究
3.2.1不同生活型植物的水源 植物生活型是与其生境相联系的,是植物体与环境间某种程度上统一性的反映。不同生活型的植物即使处于同一生境之中,其水分来源也是不同的[19-20],出现这一现象的原因可能是根系空间分布的不同以及植物叶片物候的差异[21]。水源的不同使得植物之间水分利用模式呈现互补性,比如同一生境的共生植物会分别利用不同深度的土壤水[22],这种用水策略能大大促进物种之间的共存,使群落更稳定[23]。若将植物水分利用模式与植物的生理生态状况、环境条件相联系,还能获得植物增强其水分管理的重要信息,从而指导人工群落的建设[24]。一般情况下,草本植物主要利用最近一次降雨所补给的浅层土壤水;除深根性灌木主要利用深层土壤水或地下水外,其余大部分灌木利用的是混合水源;落叶植物在夏季受水分胁迫时,利用地下水的比例比针叶植物更多。
3.2.2不同水源对植物水分的补给 水源在支撑生态系统植物群落生长的同时,会不断影响和塑造植物的特性,使得植物产生一系列获得与利用水分的特性,如根系空间分布结构,水力特征和气孔功能等。同时,植物可能会在遗传和个体发育等方面发展根系来利用水分[25],因此,研究不同水源对植物的补给具有重要的意义。
雨水作为植物的重要水源,是大部分学者研究的重点,有研究发现,干旱地区的植物会在雨季储存大量的雨水来满足旱季的用水量[26]。土壤水是植物能直接利用的水源,即使生长在河岸边的植物也并非直接将河水作为其主要的水源[27]。除了土壤水,有的植物还能利用具有储水能力的石灰岩基岩中的水[28]。而在干旱半干旱地区,地下水又会成为植物的主要水源[29]。除此之外,还有学者展开了雾水,露水和土壤、叶片吸附水的相关研究[30-32]。
值得一提的是植物的用水模式及其水分利用率是与水量相关的。在一定范围内,植物对某一水源的利用率是随着水量的增加而增加的[19,33-34],这种随水量变化的水分利用模式不但能增强植物的储水能力,还能维持其正常的蒸腾作用。
3.2.3不同季节植物对水源的选择 由于太阳辐射或季风变化,会使植物水源的水量及其氢氧同位素值发生季节变化,水量的变化会使植物相应地调整自身的用水策略,而同位素值的变化使得研究水源变化规律成为可能。一般来说,大部分植物在旱季主要吸收深层的土壤水和地下水,而到雨季水量充足时会转而吸收浅层的土壤水,一定阈值内,其吸收利用率与可获得的水量是成正比的[35-37]。
同时,由于雨水的季节性补给以及土壤表层水分的蒸发影响,土壤垂直剖面含水的氢氧同位素组成会出现梯度变化。据此可测定不同深度土壤水及植物木质部水分的氢氧同位素值,计算出各土层水分的贡献率,从而探索其季节变化规律[27,38-40]。
4 碳稳定同位素在研究植物水分利用效率中的应用
水分利用效率(WUE)是反映植物水分利用特性的一个重要参数,主要取决于植物的光合、呼吸与蒸腾3个作用的耦合过程[5]。其中,光合作用是自然界中产生分馏的最重要过程,光合作用途径不同的植物,即C3、C4和CAM植物,其不同的分馏方式使得其具有显著不同的δ13C值,据此可由δ13C值来区分植物的光合作用类型[41]。此外,环境对植物光合作用过程中CO2的羧化和叶片气孔导度的影响也会造成植物稳定同位素13C的分馏产生差异[42]。
4.1 稳定碳同位素研究植物水分利用效率的方法
稳定碳同位素Δ值是表示碳稳定同位素分馏效应大小的一种方法,表示如下[43]:
Δ= (δ13Ca-δ13Cp)/(1 +δ13Cp)≈
δ13Ca-δ13Cp
(8)
对于C3植物而言,δ13C分析是评估植物叶片中细胞间平均CO2浓度的有效方法。C3植物的δ13C值可由下式来表示[43]:
δ13Ca=δ13Ca-a -(b3-a ) ×Ci/Ca
(9)
即ΔC3=a+(b3-a)×Ci/Ca
(10)
其中:δ13Cp和δ13Ca分别为植物叶片组织及大气中CO2的碳同位素比率;a是气孔扩散过程中发生的分馏,其分馏值约为4.4‰;b3为羧化反应过程中发生的同位素分馏,其值约为30‰;而Ci和Ca分别为细胞间和大气的CO2浓度。
对于C4以及CAM植物而言,13C的分馏要显得更为复杂。首先,进入到C4和CAM植物叶片中的CO2一开始是由PEP羧化酶固定的,不同于C3植物是先由RuBisCO固定CO2;其次,C4和CAM植物还涉及到CO2通过气孔的扩散效应,以及溶解和转化为HCO3-等复杂过程的分馏[44]。因此,它们各自表现出不同δ13C值。一般来说,C3植物的δ13C值介于-22‰ ~-38‰;C4植物的δ13C值则介于-9‰ ~-16.8‰;而CAM植物固定CO2的模式虽然与C4植物相似,但在不同的干旱条件下,它能在C3和CAM途径间进行任意切换,所以其δ13C值介于典型的C3和C4植物之间[18]。
对于C4植物而言,其δ13C值的表示可以简化如下[43]:
δ13Cp=δ13Ca-a-(b4+b3φ-a)×
Ci/Ca
(11)
即ΔC4=a+(b4+b3φ-a)×Ci/Ca
(12)
其中:b4表示的是PEP羧化酶羟化反应过程中发生的同位素分馏,其值约为-5.7‰;b3为RuBisCO酶发生羟化反应产生的同位素分馏,其值约为30‰,φ为维管束鞘中的CO2渗漏到叶肉细胞间隙中,没有被PEP羟化酶所固定的比例,约为0.21。
计算植物水分利用效率的公式如下[43]:
A=g×(Ca-Ci)/1.6
(13)
E=g×ΔV
(14)
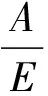

(15)
式中:A代表光合速率;E代表蒸腾速率;g代表气孔传导率;ΔV代表叶片内外水气压差;而数值1.6表示的是水蒸汽和CO2在空气中的扩散比率。
由上述所有公式可以看出,植物叶片δ13C值(δ13Cp)与胞间CO2浓度(Ci)相关,δ13C值随Ci值增大而降低,又由于WUE与Ci值之间显著相关,可发现高的WUE对应着较高的δ13C值,即WUE与δ13C值呈正相关。因此,通过测定植物叶片δ13C值就可用来间接反映植物的WUE了。由于所测定的δ13C值是植物整个生长周期中对碳的吸收,所以它表示的是植物的长期水分利用效率。
4.2 植物水分利用效率的研究
4.2.1不同光合作用途径植物δ13C与WUE的关系 Farquhar等认为,植物组织的δ13C和Δ13C与C3植物的WUE具有很强的相关性,可以作为C3植物WUE的间接测定指标[45],这一观点也得到了学术界普遍的认同。国际上,碳稳定同位素技术比较广泛地应用于指导C3作物的生产,Tambussi等[46]就对C3作物的Δ值与产量、生理特征及其与WUE的关系进行了整合报道。相比之下,目前对于C4植物的研究还比较少,常见的主要是对玉米等C4作物的研究[47-48],这主要是因为很多C4植物的δ13C值与其WUE的相关性不太明显[49]。而关于CAM植物WUE的研究,Gravatt等[50]]曾发现CAM植物苹果酸波动与WUE的增加没有相关性,但植物叶片的δ13C却能反应植物在不同水分条件下光合途径以及WUE的变化,但仍需大量的试验研究与调查来证实。
4.2.2不同生长型植物δ13C与WUE的关系 不同生长型植物的WUE是不同的,乔木、灌木、草本和藤本植物叶片之间的δ13C值存在显著差异[51],水分利用状况也因此而不同。一般来说1年生植物δ13C值低于多年生植物;早开花植物低于晚开花植物;常绿乔木>落叶乔木=常绿和落叶灌木=常绿草本>落叶草本=苔藓[52-56],其水分利用效率也呈现同样特征。同时,不同植物种δ13C值年间变化差别很大,受环境影响各不相同。有的主要是由于降水的变化引起的,其δ13C值随降雨量的增加而降低[56];有的则是因为温度变化和海拔高度等因素引起的,WUE均随年均温或海拔的增加而呈不同程度的降低[57],反应了不同生长型植物WUE对环境变化的不同响应。
4.2.3不同时间尺度上植物δ13C与WUE的关系 植物组织的碳同位素组成是植物在生长过程中发生的一系列复杂的整合CO2的生理生化反应所形成的,分析不同植物碳水化合物和碳库的δ13C值,研究的时间跨度可以为1个生长季或植物的整个生活史(结构性碳),甚至对1个月或1个世纪的平均WUE进行研究。
大尺度WUE的研究,比如说近百年来全球大气CO2浓度的持续增高使得植物δ13C值升高,WUE普遍提高[58-60]。
中等尺度,比如20世纪中后期,我国暖温带落叶阔叶林和亚热带常绿阔叶林大部分植物δ13C值持续降低比较明显[61],植物WUE呈下降趋势。但青藏高原高寒植被区域中辽东栎δ13C呈增加趋势,这可能与青藏高原高海拔的地理位置以及高温、强光照的特殊气候条件有关[62-63]。
小尺度,比如研究幼树与成年树种的WUE,由于植物WUE与水分在树体内传输的距离有关,针阔叶林内幼树的WUE和δ13C显著低于成年树[64]。
尺度再小如植物生长季WUE的变化,很多研究表明,越往生长后期,WUE与植物δ13C值的正相关性越强,WUE呈上升趋势[65-66]。
4.2.4不同环境条件下植物δ13C与WUE的关系 植物δ13C值的差别取决于其遗传特性,在良好而稳定的生态环境中,δ13C值的变动较少。然而,δ13C值的微小差别往往反映了植物生理活动发生较大的差异,有研究表明,非胁迫环境条件的变化可导致样品高达±3‰~±5‰的变幅[67]。影响δ13C值的外界因子有很多(如光照、降水、温度、湿度、CO2浓度、土壤水分状况等),且各因子的影响程度相同[8]。目前,很多研究结果均表明,一定程度的水分胁迫下,植物的δ13C、蒸腾效率和WUE会提高,与叶片的δ13C值呈正相关[44,68]。简而言之,温暖湿润环境下的植物δ13C值比生长在寒冷干旱环境下的植物低,其WUE也同样偏低[61,69-73]。
5 存在问题与展望
在植物水分来源的研究中,氢氧稳定同位素技术是了解植物水分来源很好的手段,但在判别不同深度土壤水的贡献率时,应避免水样之间氢氧同位素δ值的相对差异在10%以内[74],在土壤水取样之前必须考虑好取样土层的深度。同时,氢氧稳定同位素对环境极其敏感,所以在采样、处理样品的过程中一定要注意避免样品的蒸发以及其他水分对样品的污染,否则会对计算结果造成极其严重的影响。
在植物水分利用效率的研究中,由于植物WUE和δ13C值是受到多个环境因子和植物因子共同影响的,这可能造成植物δ13C值和WUE 间相关性不强而表现出彼此独立的变化。此外,在不同尺度上,如从植物叶片水平到景观水平,以及昼夜变化到年际变化,简单的δ13C与Ci/Ca以及植物水分利用效率WUE之间的线性关系是不可靠的,还需寻求分析各种因素影响大小的方法。同时,对C4和CAM植物的δ13C与WUE关系的研究还需加强。
如今,稳定同位素技术在生态学研究中的运用越来越广,它不仅可以研究植被在全球气候变化下的响应机制,还能为指导植被的恢复工作提供理论依据,具有重要的理论与实践意义以及较为广阔的发展前景。
[1] Neilson R P. A model for predicting continental-scale vegetation distribution and water balance[J]. Ecological Applications, 1995, 5(2): 362-385.
[2] Schimel D S.Population and community processes in the response of terrestrial ecosystems to global change[C]//Kareiva P M,Kingsolver J G,Huey R B.Biotic interactions and global change.Sunderland,USA:Sinauer Associates,1993:45-54.
[3] Flanagan L B,Ehleringer J R,Marshall J D.Differential uptake of summer precipitation among co-occurring trees and shrubs in a pinyon-juniper woodland[J].Plant,Cell & Environment,1992,15(7):831-836.
[4] 聂云鹏, 陈洪松, 王克林. 土层浅薄地区植物水分来源研究方法[J].应用生态学报, 2010, 21(9): 2427-2433.
[5] 李机密, 黄儒珠, 王健, 等. 陆生植物水分利用效率[J]. 生态学杂志, 2009, 28(8): 1655-1663.
[6] Urey H C, Brickwedde F G, Murphy G M. A hydrogen isotope of mass 2 and its concentration[J]. Physical Review, 1932, 40(1): 1.
[7] 丁悌平. 稳定同位素地球化学研究新况[J]. 地学前缘, 1994, 1(4): 191-203.
[8] Dawson T E, Mambelli S, Plamboeck A H, et al. Stable isotopes in plant ecology[J]. Annual Review of Ecology and Systematics, 2002: 507-559.
[9] Fry B. Stable Isotope Ecology[M]. New York: Springerverlag , 2006.
[10] 陈世苹, 白永飞, 韩兴国. 稳定性碳同位素技术在生态学研究中的应用[J]. 植物生态学报, 2002, 26(5): 549-560.
[11] Máguas C, Griffiths H. Applications of stable isotopes in plant ecology[M]//Progress in Botany. Springer Berlin Heidelberg, 2003: 472-505.
[12] Gonfiantini R, Gratziu S, Tongiorgi E. Isotopes and radiation in soil-plant nutrition studies[M]// Oxygen Isotopic Compotition of Water in Leaves. Vienna: International Atomic Energy Agency, 1965: 405-410.
[13] Wershaw R L, Friedman I, Heller S J, et al. Hydrogen isotopic fractionation of water passing through trees[M]//Advance in Organic Geochemistry.New York:Pergamon,1966:405-410.
[14] Phillips D L. Mixing models in analyses of diet using multiple stable isotopes: a critique[J]. Oecologia, 2001, 127(2): 166-170.
[15] Phillips D L, Gregg J W. Source partitioning using stable isotopes: coping with too many sources[J]. Oecologia, 2003, 136(2): 261-269.
[16] 巩国丽, 陈辉, 段德玉. 利用稳定氢氧同位素定量区分白刺水分来源的方法比较[J]. 生态学报, 2011, 31(24): 7533-7541.
[17] Pate J S, Dawson T E. Assessing the performance of woody plants in uptake and utilisation of carbon, water and nutrients: implications for designing agricultural mimic systems[J]. Agroforestry Systems, 1999, 45(1/2/3): 245-275.
[18] Phillips D L, Gregg J W. Uncertainty in source partitioning using stable isotopes[J]. Oecologia, 2001, 127(2): 171-179.
[19] Duan D Y, Ouyang H, Song M H, et al. Water sources of dominant species in three alpine ecosystems on the Tibetan Plateau, China[J]. Journal of Integrative Plant Biology, 2008, 50(3): 257-264.
[20] Ewe S M L, Sternberg L S L, Busch D E. Water-use patterns of woody species in pineland and hammock communities of South Florida[J]. Forest Ecology and Management, 1999, 118(1): 139-148.
[21] Nie Y, Chen H, Wang K, et al. Seasonal water use patterns of woody species growing on the continuous dolostone outcrops and nearby thin soils in subtropical China[J]. Plant and Soil, 2011, 341(1/2): 399-412.
[22] Asbjornsen H, Shepherd G, Helmers M, et al. Seasonal patterns in depth of water uptake under contrasting annual and perennial systems in the Corn Belt Region of the Midwestern US[J]. Plant and Soil, 2008,308(112):69-92.
[23] Xu Q, Li H, Chen J, et al. Water use patterns of three species in subalpine forest, Southwest China: the deuterium isotope approach[J]. Ecohydrology, 2011, 4(2): 236-244.
[24] Kulmatiski A, Beard K H, Stark J M. Exotic plant communities shift water-use timing in a shrub-steppe ecosystem[J]. Plant and Soil, 2006, 288(1): 271-284.
[25] Weltzin J F, McPherson G R. Spatial and temporal soil moisture resource partitioning by trees and grasses in a temperate savanna, Arizona, USA[J]. Oecologia, 1997, 112(2): 156-164.
[26] Yang H, Auerswald K, Bai Y, et al. Complementarity in water sources among dominant species in typical steppe ecosystems of Inner Mongolia, China[J]. Plant and Soil, 2011, 340(1/2): 303-313.
[27] Sánchez Pérez J M, Lucot E, Bariac T, et al. Water uptake by trees in a riparian hardwood forest (Rhine floodplain, France)[J]. Hydrological Processes, 2008, 22(3): 366-375.
[28] Nie Y, Chen H, Wang K, et al. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China[J]. Journal of Hydrology, 2012, 420: 264-274.
[29] 赵良菊, 肖洪浪, 程国栋,等. 黑河下游河岸林植物水分来源初步研究[J]. 地球学报, 2008, 29(6): 709-718.
[30] Liu W J, Li P J, Duan W P, et al. Dry-season water utilization by trees growing on thin karst soils in a seasonal tropical rainforest of Xishuangbanna, Southwest China[J]. Ecohydrology, 2013,7(3):927-935.
[31] Dawson T E. Fog in the California redwood forest: ecosystem inputs and use by plants[J]. Oecologia, 1998, 117(4): 476-485.
[32] Liu W J, Liu W Y, Li P J, et al. Using stable isotopes to determine sources of fog drip in a tropical seasonal rain forest of Xishuangbanna, SW China[J]. Agricultural and Forest Meteorology, 2007, 143(1): 80-91.
[33] 周雅聃, 陈世苹, 宋维民, 等. 不同降水条件下两种荒漠植物的水分利用策略[J]. 植物生态学报, 2011, 35(8): 789-800.
[34] 苑晶晶, 袁国富, 罗毅, 等. 利用δ18O信息分析冬小麦对浅埋深地下水的利用[J]. 自然资源学报,2009, 24(2):360-368.
[35] Dawson T E, Pate J S. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation[J]. Oecologia, 1996, 107(1): 13-20.
[36] McCole A A, Stern L A. Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in water[J]. Journal of Hydrology, 2007, 342(3): 238-248.
[37] Snyder K A, Williams D G. Water sources used by riparian trees varies among stream types on the San Pedro River, Arizona[J]. Agricultural and Forest Meteorology, 2000, 105(1): 227-240.
[38] Eggemeyer K D, Awada T, Harvey F E, et al. Seasonal changes in depth of water uptake for encroaching treesJuniperusvirginianaandPinusponderosaand two dominant C4 grasses in a semiarid grassland[J]. Tree Physiology, 2009, 29(2): 157-169.
[39] Wang P, Song X, Han D, et al. A study of root water uptake of crops indicated by hydrogen and oxygen stable isotopes: a case in Shanxi Province, China[J]. Agricultural Water Management, 2010, 97(3): 475-482.
[40] 孙守家, 孟平, 张劲松, 等. 华北石质山区核桃-绿豆复合系统氘同位素变化及其水分利用[J]. 生态学报, 2010, 30(14): 3717-3726.
[41] Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis[J]. Annual Review of Plant Biology, 1989, 40(1): 503-537.
[42] 殷树鹏, 张成君, 郭方琴, 等. 植物碳同位素组成的环境影响因素及在水分利用效率中的应用[J]. 同位素, 2008, 21(1): 46-53.
[43] Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis[J]. Annual Review of Plant Biology, 1989, 40(1): 503-537.
[44] Hubick K T, Farquhar G D, Shorter R. Correlation between water-use efficiency and carbon isotope discrimination in diverse peanut(Arachis)germplasm[J]. Functional Plant Biology, 1986, 13(6): 803-816.
[45] Farquhar G D, Richards R A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes[J]. Functional Plant Biology, 1984, 11(6): 539-552.
[46] Tambussi E A, Bort J, Araus J L. Water use efficiency in C3 cereals under Mediterranean conditions: a review of physiological aspects[J]. Annals of Applied Biology, 2007, 150(3): 307-321.
[47] Monneveux P, Sheshshayee M S, Akhter J, et al. Using carbon isotope discrimination to select maize (ZeamaysL.) inbred lines and hybrids for drought tolerance[J]. Plant Science, 2007, 173(4): 390-396.
[48] 张丛志, 张佳宝, 赵炳梓, 等. 玉米水分利用效率, 碳稳定同位素判别值和比叶面积之间的关系[J]. 作物学报, 2009, 35(6): 1115-1121.
[49] Wayne Polley H, Johnson H B, Derner J D. Soil- and plant-water dynamics in a C3/C4 grassland exposed to a subambient to superambient CO2gradient[J]. Global Change Biology, 2002, 8(11): 1118-1129.
[50] Gravatt D A, Martin C E. Comparative ecophysiology of five species of Sedum(Crassulaceae)under well-watered and drought-stressed conditions[J]. Oecologia, 1992, 92(4): 532-541.
[51] 渠春梅,韩兴国,苏波,等.云南西双版纳片断化热带雨林植物叶片δ13C值的特点及其对水分利用效率的指示[J].植物学报,2001,43(2):186-192.
[52] 周雅聃, 陈世苹, 宋维民, 等. 不同降水条件下两种荒漠植物的水分利用策略[J]. 植物生态学报, 2011, 35(8): 789-800.
[53] Kloeppel B D, Gower S T, Treichel I W, et al. Foliar carbon isotope discrimination inLarixspecies and sympatric evergreen conifers: a global comparison[J]. Oecologia, 1998, 114(2): 153-159.
[54] Smedley M P, Dawson T E, Comstock J P, et al. Seasonal carbon isotope discrimination in a grassland community[J]. Oecologia, 1991, 85(3): 314-320.
[55] Brooks J R, Flanagan L B, Buchmann N, et al. Carbon isotope composition of boreal plants: functional grouping of life forms[J]. Oecologia, 1997, 110(3): 301-311.
[56] 李明财,黎贞发, 易现峰, 等. 青藏高原东部高寒草甸植物δ13C 年间变化及其环境分析[J]. 生态环境, 2007, 16(4): 1205-1210.
[57] 苏波, 韩兴国, 李凌浩, 等. 中国东北样苦草原区植物δ13C值及水分利用效率对环境梯度的响应[J]. 植物生态学报, 2000, 4(6): 648-655.
[58] Duquesnay A, Breda N, Stievenard M, et al. Changes of tree-ringδ13C and water-use efficiency of beech (FagussylvaticaL.) in north-eastern France during the past century[J]. Plant, Cell & Environment, 1998, 21(6): 565-572.
[59] Bert D, Leavitt S W, Dupouey J L. Variations of woodδ13C and water-use efficiency ofAbiesalbaduring the last century[J]. Ecology, 1997, 78(5): 1588-1596.
[60] 郑凤英, 彭少麟, 赵平. 两种山黄麻属植物在近一世纪里气孔密度和潜在水分利用率的变化[J]. 植物生态学报, 2001, 25(4): 405-409.
[61] 郑淑霞, 上官周平. 辽东栎叶片气孔密度及δ13C值的时空变异[J].林业科学,2005,42(2):30-36.
[62] 旺罗, 吕厚远, 吴乃琴, 等. 青藏高原现生禾本科植物的δ13C 与海拔高度的关系[J]. 第四纪研究, 2003, 23(5): 573-580.
[63] 刘光琇, 陈拓, 安黎哲, 等.青藏高原北部植物叶片碳同位素组成特征的环境意义[J]. 地球科学进展, 2004, 19:749-753.
[64] 孙双峰, 黄建辉, 林光辉, 等. 三峡库区岸边共存松栎树种水分利用策略比较[J]. 植物生态学报, 2006, 30(1): 57-63.
[65] 苏培玺, 严巧嫡, 陈怀顺. 荒漠植物叶片或同化枝δ13C值及水分利用效率研究[J]. 西北植物学报, 2005,25(4):727-732.
[66] 王国安, 韩家懋, 刘东生. 中国北方黄土区C3草本植物碳同位素组成研究[J]. 中国科学, 2003, 33: 550-556.
[67] O′Leary M H. Carbon isotopes in photosynthesis[J]. Bioscience, 1988, 38(5): 328-336.
[68] Osório J, Osório M L, Chaves M M, et al. Effects of water deficits on13C discrimination and transpiration efficiency ofEucalyptusglobulusclones[J]. Functional Plant Biology, 1998, 25(6): 645-653.
[69] Schulze E D, Williams R J, Farquhar G D, et al. Carbon and nitrogen isotope discrimination and nitrogen nutrition of trees along a rainfall gradient in northern Australia[J]. Functional Plant Biology, 1998, 25(4): 413-425.
[70] Bowling D R, McDowell N G, Bond B J, et al.13C content of ecosystem respiration is linked to precipitation and vapor pressure deficit[J]. Oecologia, 2002, 131(1): 113-124.
[71] Marshall J D, Zhang J. Carbon isotope discrimination and water-use efficiency in native plants of the north-central Rockies[J]. Ecology, 75(7):1994,1887-1895.
[72] Körner C, Farquhar G D, Roksandic Z. A global survey of carbon isotope discrimination in plants from high altitude[J]. Oecologia, 1988, 74(4): 623-632.
[73] 陈拓,杨梅学, 冯虎元, 等. 青藏高原北部植物叶片碳同位素组成的空间特征[J]. 冰川冻土, 2003, 25(1): 83-87.
[74] Burgess S S O, Adams M A, Turner N C, et al. Characterisation of hydrogen isotope profiles in an agroforestry system: implications for tracing water sources of trees[J]. Agricultural Water Management, 2000, 45(3): 229-241.
(责任编辑 张 坤)
Application of Stable Isotope Techniques in the Study of Plant Water Sources and Use Efficiency
WU Jun-en1,2, LIU Wen-jie1, ZHU Chun-jing1,2
(1.Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences,Menglun Yunnan 666303, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
This paper described the application of stable hydrogen, carbon and oxygen isotopes techniques in tracing sources of plant water and water use efficiency. Since the process that plant roots uptake water is non-fractionating, plant xylem water has the same composition of stable hydrogen and oxygen isotopes as soil water utilized by the plant. By analyzing the composition of hydrogen and oxygen isotopes of plant xylem water and potential sources of water, we could quantitatively determine the contributions of the sources to plants. For the study of water use efficiency, there were many researches showed that the composition of stable carbon isotopes of leaves can be an indirect indicator of plant water use efficiency. Photosynthesis is one of the most significant processes of carbon isotope fractionation in nature. So, stable carbon isotopes techniques had become the best way to study the long-term water use efficiency of plant leaves presently. As a safe tracer material, stable isotopes technique will help people to understand the plant’s ecophysiological process. Anyway, with the continuous improvement of techniques, stable isotope techniques will be widely used in many areas of ecology.
stable isotope;plant; water sources;water use efficiency
2014-01-20
国家自然科学基金项目(41271051、31170447)资助;云南省应用基础研究重点项目(2013FA022)资助。
刘文杰(1969—),男,博士,研究员。研究方向:生态水文。Email:lwg@xtbg.ac.cn。
10.3969/j.issn.2095-1914.2014.05.020
S718.45
:A
:2095-1914(2014)05-0103-08
第1作者:吴骏恩(1990—),男,硕士生。研究方向:生态水文。Email:wujunen@xtbg.ac.cn。
