篮子鱼内脏粗提液制备长心卡帕藻和细齿麒麟菜原生质体的初步研究
2014-06-26李俊鹏刘建国庞通李
李俊鹏 ,刘建国庞 通李 虎
(1.中国科学院 海洋研究所 生物工程中心,山东 青岛 266071;2.中国科学院大学,北京 100049)
卡帕藻属和麒麟菜属红藻,其富含红藻多糖卡拉胶,一直是生产卡拉胶的主要原料[1-3]。卡拉胶因其具有良好的凝胶性、保水性、增稠性和助悬性,在食品和化工中有着广泛的应用[4-6],研究表明,麒麟菜类海藻还具有工业生产乙醇的潜质[7-10],此类产卡拉胶红藻资源具有巨大开发价值。目前我国产卡拉胶红藻的养殖规模相对较小[2-3,11],这与麒麟菜在20~30℃下生长有关[1,12-14],导致养殖区域受到限制,另外,其长期营养繁殖造成种质退化,产量和质量均下降,敌害、病害频发[2]。因此,需要筛选和培育生长速率高、胶含量和质量好、抗病力强、耐低温的麒麟菜,应对日益增长的养殖需要。
尽管麒麟菜理论上具有无性生殖和有性生殖世代,但通常难以获得孢子和配子[2,15-16],通过有性生殖方式进行种苗繁育效率低,难度很大。运用细胞工程技术,进行麒麟菜的种苗繁育可以拓展拓宽新的途径。而原生质体的获得是进行细胞工程繁育,细胞融合等的重要前提,也为定向改良提供技术支撑。20世纪80年代,有关海藻原生质体研究有很多报道[17-22],但因诸多条件的限制,所制备原生质体效率都不高,大多未能用于海藻繁育的实际应用。近年来,Reedy[23]和 Yeong[24]分别制备出紫菜和江蓠的原生质体。Cheney[25]利用孢子制得原生质体,并通过细胞融合的方法获得了麒麟菜细胞融合体,获得了生长速率高的融合体植株。这为通过麒麟菜原生质体,进行遗传选育,获得目标株系提供借鉴。制备海藻原生质体,采用合适的酶是最关键的一环,Zablakis等[26]利用纤维素酶和卡拉胶酶混合酶液,酶解制备长心卡帕藻原生质体,但所用卡拉胶酶在很多国家均难以获得。Salvador等[27]通过鲍鱼内脏提取液获得长心卡帕藻原生质体,观察到细胞壁再生,但该方法在不同长心卡帕藻株系原生质体制备中的使用效果不一,差别很大。
本文采用热带海域广泛分布的杂食性篮子鱼为材料,从篮子鱼内脏提取制备酶液,用于麒麟菜原生质体的分离,为后期的麒麟菜组织培养及细胞工程育种打下基础。
1 材料与方法
1.1 实验材料
本实验采用 2种红藻,棕黑色长心卡帕藻(Kappaphycus alvarezii),红褐色细齿麒麟菜(Eucheuma denticulatum),上述海藻材料取自中国科学院海洋研究所海南陵水热带产卡拉胶海藻实验基地(18°27′N,110°5′E);篮子鱼(Siganus fuscessens)采于海南省陵水黎族自治县黎安港湾,置于网箱中暂养,每天用长心卡帕藻和细齿麒麟菜喂养,篮子鱼在饲喂 1周后,用于粗酶液的制备。
1.2 原生质体制备
1.2.1 粗酶解液的制备
取长心卡帕藻和细齿麒麟菜喂养篮子鱼5~10只,饲喂 1周后取其胃和肝脏放入 4℃预冷的小烧杯中,按每克内脏加50 mmol/L pH为6.0的磷酸钠盐缓冲液20 mL,利用组织研磨器,充分研磨成浆。研磨后的匀浆分别用20、8和1 μm孔径的滤纸分级过滤。向过滤后的滤液中添加0.6 mol/L甘露醇和5 mmol/L氯化钙,冰浴保存备用。
1.2.2 材料准备
取生长状态良好的上述红藻,先用海水冲洗干净,再用消毒海水反复洗涤多次,再用无菌纸巾吸干并转入消毒液(100 mg氯霉素 + 200 mL无菌海水)中处理0.5~1.0 h。称取10 g经过上述消毒的藻段,置于多功能料理机(九阳 JYL-CO20)中并加入 100 mL无菌海水,连续搅拌2次,每次1 min,以打碎藻段,静止5 min后,弃去上清液(约100 mL),剩余底层的藻泥(约10 g)作为进一步酶解的材料,待用。
1.2.3 酶解与原生质体纯化
取制取的上述酶液和藻泥按设定的比例(详见1.2.4)加入10 mL的离心管中混匀,然后置于摇床(国华 ZD-85)上震荡酶解,震荡速度100 r/min,分别开展了不同酶解条件(详见 1.2.4)下的酶解实验,以每只上述离心管为一个单位酶反应体系。
酶解结束前,向酶解液中加入3 mL洗脱液,然后放回摇床继续酶解1 h,选用500 μm的筛绢过滤酶解液,将滤液转入离心管中,在相对离心力为218 g(湘仪 TGL-16M)的条件下离心5 min;吸取离心管中最上层的清液5 mL,并加入5 mL洗脱液(高压蒸汽灭菌过的含有0.6 mol/L甘露醇和5 mmol/L氯化钙PES培养基)混匀,在218 g离心力条件下离心5 min,弃上清,向离心管中加入1 mL洗脱液,然后转入1.5 mL的离心管中。
1.2.4 酶解条件梯度实验设计
实验设置3个酶解浓度组合。酶浓度Ⅰ,0.1 g藻泥加3 mL酶液;酶浓度Ⅱ,0.2 g藻泥加3 mL酶液;酶浓度Ⅲ,0.4 g藻泥加3 mL酶液。酶解温度25℃,酶解pH 6.0,酶液制备和酶解方法同前文所述,分别研究酶解(12,24,48 h)时,不同酶解浓度组合对其原生质体产量和产率的影响。
酶解温度实验设置3个梯度,分别为20,25和30℃,按照0.1 g藻泥加3 mL酶液的比例进行酶解,酶液制备和酶解方法如前文所述,酶解pH 6.0,期间利用摇床连续摇动,分别记录12,24和48 h时温度对原生质体产量和产率的作用效果。
酶解pH实验设置3个梯度,分别为6.0,7.0和8.0,酶浓度选取0.1 g藻泥加3 mL酶液的比例,酶解温度25℃,分别记录酶解12,24和48 h时,不同pH对原生质体产量和产率的作用效果。
1.2.5 原生质体制备效果鉴定
1)原生质体去壁程度: 用蒸馏水处理待检样品,在显徽镜下观察细胞大小的变化,细胞膨胀均匀,破碎后无细胞壁残余,表明所制备的原生质体良好;若细胞单方向外涨至破碎,且胀碎后存在一定的细胞壁残余,表明所制备的原生质体尚有细胞壁存在,原生质体制备不完全;若细胞不发生膨胀,细胞体积没有变化,表明细胞壁依然完整,原生质体制备不成功。
2)原生质体完整性检测: 向制备的原生质体中加入 0.5%伊文思蓝染液染色,在显微镜下观察原生质体色泽,并利用细胞计数板计数,统计获得原生质体数目,其中细胞深色的为原生质体已经破损,颜色较浅的为完好的原生质体。
1.2.6 原生质体产量产率统计
原生质体产量监测用显微镜计数法,先以血球计数板计算出每毫升酶解液中原生质体数量,然后折算单位酶反应体系所获得的原生质体的总数目;原生质体产率为每克藻泥获得原生质体的数目,即原生质体产量除以实验所用藻泥质量。
2 结果
2.1 两种海藻原生质体产量和产率对比
对比2种藻原生质体的制备效果(图1),得出:1)细齿麒麟菜比长心卡帕藻的原生质体制备效果更好,前者制备的原生质体产量和产率都明显高于后者,统计分析效果显著(P<0.01)。2)酶解时间延长有利于原生质体制备。其中,在0.1 g藻泥加3 mL酶液处理组,48 h酶解时间内,长心卡帕藻原生质体产量从 1.2 ×103cell上升到 1.45 ×103cell(图 1.A),原生质体产率从 1.20 ×104cell/g 增加到 1.45×104cell/g(图1.C);而细齿麒麟菜原生质体产量从 1.35×103cell到1.75×103cell(图 1.A),其产率从 1.35×104cell/g 增加到1.75 ×104cell/g(图1)。3)实验中海藻用量对酶解效果也产生明显影响,在等量酶浓度下,藻量越多,原生质体产量越高(图1.A、B),而其产率却越低(图1.C、D)。其中,0.1 g的长心卡帕藻和细齿麒麟菜原生质体产量分别只有 1.45 ×103cell和 1.75×103cell(图1.A)内,而0.4 g的长心卡帕藻和细齿麒麟菜原生质体产量分别达到3.83 ×103cell和4.15 ×103cell(图1.B);相反,0.1 g的长心卡帕藻和细齿麒麟菜原生质体产率分别在 1.45 ×104cell/g 和 1.75×104cell/g (图 1.C)范围内,而0.4 g的长心卡帕藻和细齿麒麟菜原生质体产率只有0.96 ×104cell/g 和 1.04×104cell/g(图 1.D)。
就获得最大原生质体产量而言,可选择的酶解条件: 25℃和pH 6.0,以及0.4 g藻泥加3 mL酶液,其酶解时间为48 h(图1.B)。在此酶解条件下,细齿麒麟菜原生质体最大产量为4.15×103cell,而长心卡帕藻的原生质体最大产量可达到 3.8×103cell(图1.B)。为得到最高原生质体产率,可选择的酶解条件:25℃和pH 6.0,以及0.1 g藻泥加3 mL酶液,其酶解时间为48 h(图1.C)。在此酶解条件下,细齿麒麟菜原生质体最高产率达到1.75×104cell/g,而长心卡帕藻的原生质体产率最高1.45×104cell/g(图1.C)。
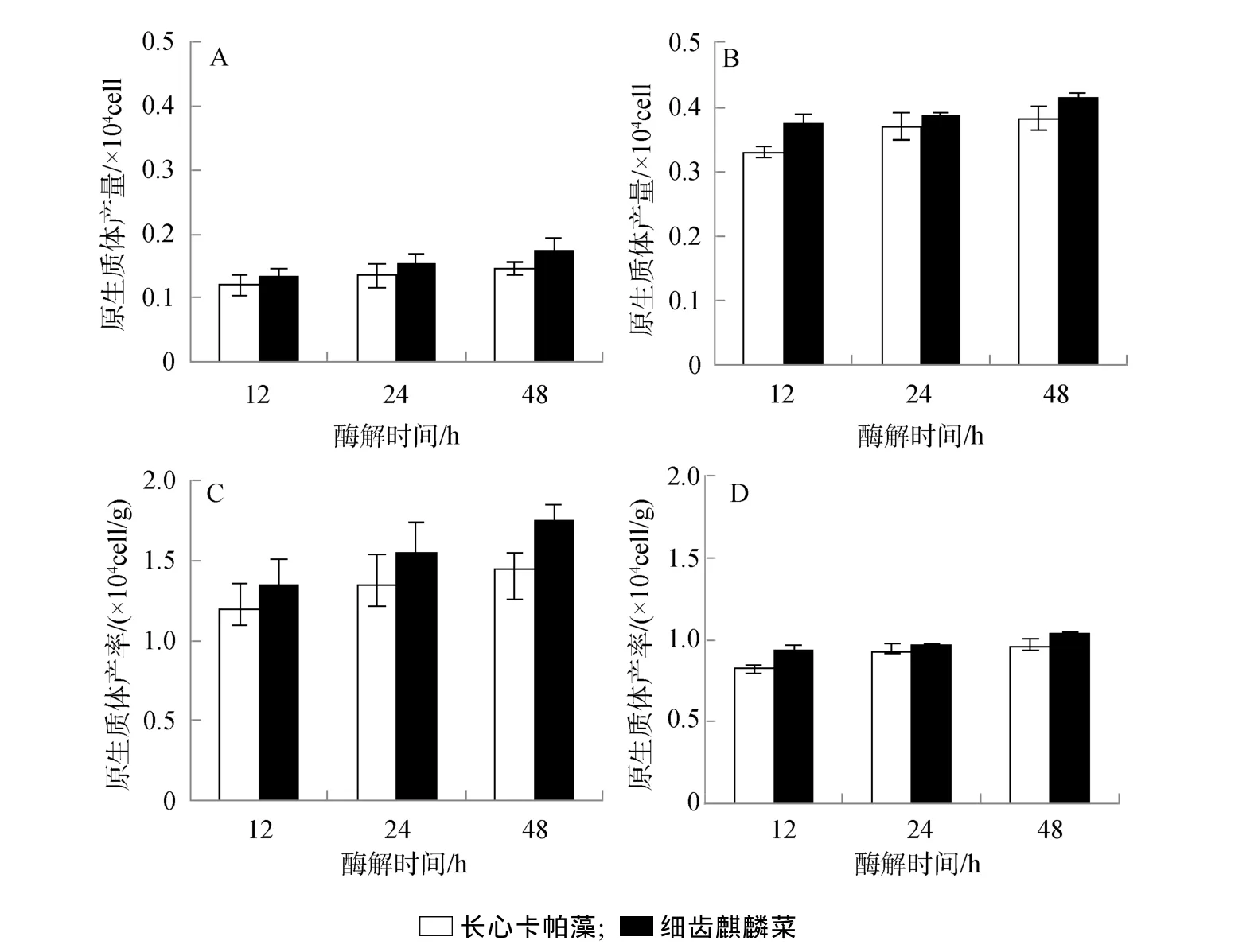
图1 长心卡帕藻与细齿麒麟菜原生质体产量和产率对比Fig.1 The comparation between total protoplast yields and the relative protoplast yields of E.denticulatum and K.alvarezii
2.2 藻泥与酶浓度配比对原生质体制备的影响
不同藻泥与酶浓度配比对长心卡帕藻和细齿麒麟菜原生质体制备效果如图2。结果表明: 2种海藻的原生质体制备效果与藻泥和酶浓度配比存在直接关系。在实验酶浓度恒定和藻泥0.1~0.4 g范围内,藻泥越多,所制备出的长心卡帕藻和细齿麒麟菜原生质体产量越多(图 2.A、B),即 0.4 g > 0.2 g > 0.1 g 藻泥酶解处理组;但原生质体产率与藻泥用量呈负相关性,所用的藻泥越高,所制备的长心卡帕藻或细齿麒麟菜原生质体产率越低(图2.C、D),即0.4 g <0.2 g < 0.1 g藻泥酶解处理组。上述结果与图1结果一致,也进一步证实在等量酶浓度下,藻量越多,原生质体产量越高。
在实验酶解时间范围内,2种海藻原生质体与酶解时间存在一定相关性,随着酶解时间延长,2种海藻原生质体呈增加趋势,但其产量和产率增加幅度略有不同(图2)。
2.3 pH对原生质体制备的影响
2种红藻原生质体制备与酶解酸碱度变化关系如图3所示。从中可以看出,pH对2种海藻原生质体制备存在一定程度的影响;总体而言,在弱碱性(pH8.0)条件下,无论长心卡帕藻还是细齿麒麟菜原生质体产量(图3.A、B),还是其产率(图3.C、D)均低于中性(pH7.0)和弱酸性(pH6.0)的效果(P<0.01);在中性(pH7.0)和弱酸性(pH6.0)条件下,2种海藻原生质体产量和产率基本一致,其中相对而言以弱酸性条件下酶解制备效果略高一点;随着酶解时间延长,在3个实验pH范围内,原生质体产量和产率均呈明显上升趋势。
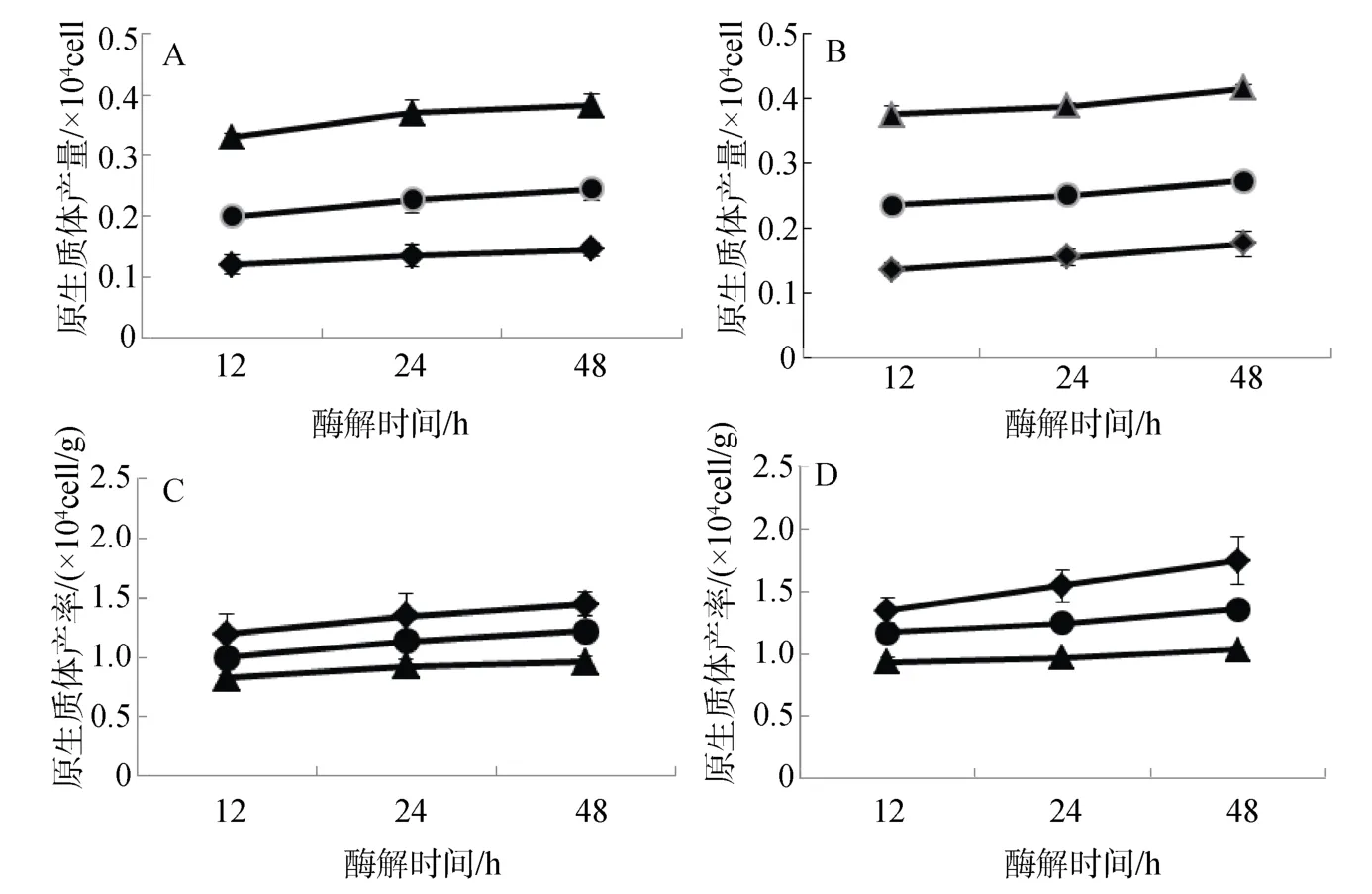
图2 藻泥和酶浓度配比对长心卡帕藻(A、C)和细齿麒麟菜(B、D)原生质体制备的影响Fig.2 Effect of dosage of enzyme extract and algal pellets on protoplast isolation of K.alvarezii and E.denticulatum
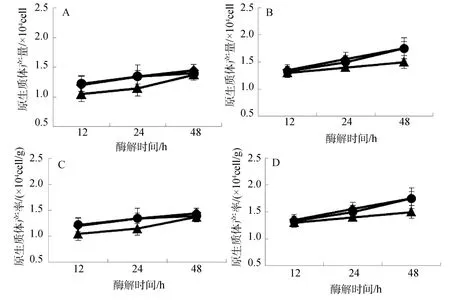
图3 pH对长心卡帕藻(A、C)和细齿麒麟菜(B、D)原生质体制备的影响Fig.3 Effect of pH on protoplast isolation of K.alvarezii and E.denticulatum
2.4 温度对原生质体制备的影响
长心卡帕藻和细齿麒麟菜原生质体制备与酶解温度变化关系如图4所示。结果表明: 酶解温度对原生质体制备存在明显影响,其中在25℃酶解条件下,两种海藻的原生质体产量一般都显著高于20℃和 30℃时的产量(图 4.A、B,P<0.01);两种海藻原生质体产率也随酶解温度变化,呈现与其原生质体产量相类似的结果(图 4.C、D);同时,在三个酶解温度梯度下,随着酶解时间的延长,原生质体产量和产率均呈线性上升趋势,并且以细齿麒麟菜在25℃酶解条件下原生质体产量和产率增幅最快(图4.B、D)。
2.5 原生质体制备效果观察
长心卡帕藻和细齿麒麟菜匀浆后的藻渣,经过篮子鱼内脏研磨提取液酶解,2种海藻细胞壁都可被有效酶解,依据酶解程度不同,制备出的原生质体有完全游离的,也有部分原生质体尚残存细胞壁(图5.B、D),游离原生质体经依文思蓝染色,可检查其细胞完整性,其中结构完整的原生质体不能吸收依文思蓝,不被染色(图 5.E),而染色后原生质体深色的表明其结构已经受损,为不完整的破损原生质体(图5.F)。
在不同酶解条件下,所获得原生质体数目占藻细胞总数目比例依然较低,尚不足细胞总数的1%(图6)。在实验酶浓度恒定和0.1~0.4 g藻泥范围内,藻泥越多,长心卡帕藻和细齿麒麟菜原生质体比例越低(图6.A);在本实验pH梯度范围内,pH 8.0时长心卡帕藻和细齿麒麟菜原生质体比例都显著低于 pH 6.0~7.0的酶解效果(P<0.01),pH 6.0~7.0的酶解条件下,2种藻的原生质体比例无显著差异(图6.B);25℃时2种藻的原生质体比例显著高于20℃和30℃时(图6.C);在48 h酶解时间内,长心卡帕藻原生质体随时间延长逐渐升高(P<0.01),而细齿麒麟菜在48 h时的原生质体制备效率较比24 h略有下降(P>0.05)。
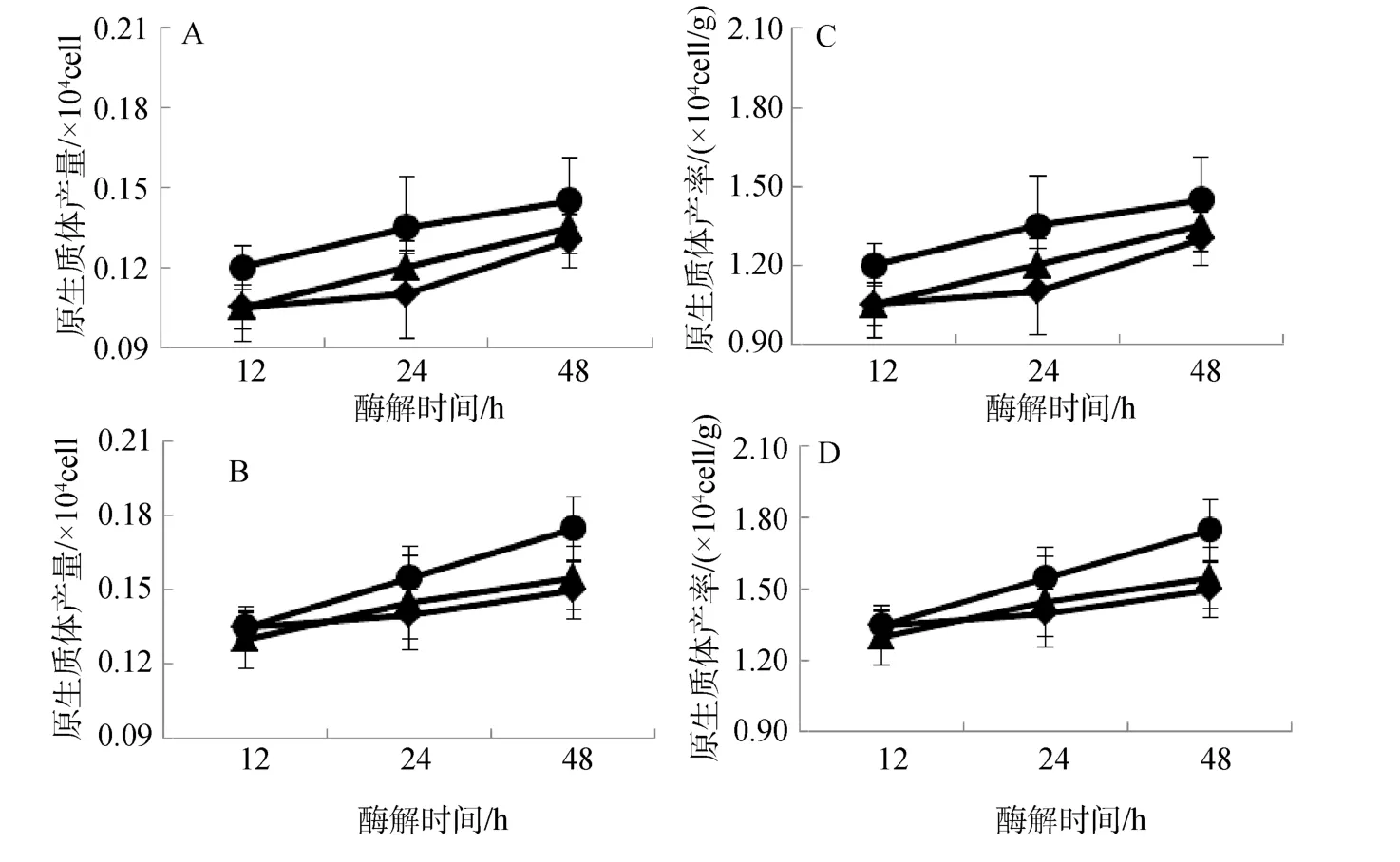
图4 温度对长心卡帕藻(A、C)和细齿麒麟菜(B、D)原生质体制备的影响Fig.4 Effect of temperature on protoplast isolation of K.alvarezii(A and C)and E.denticulatum(B and D)
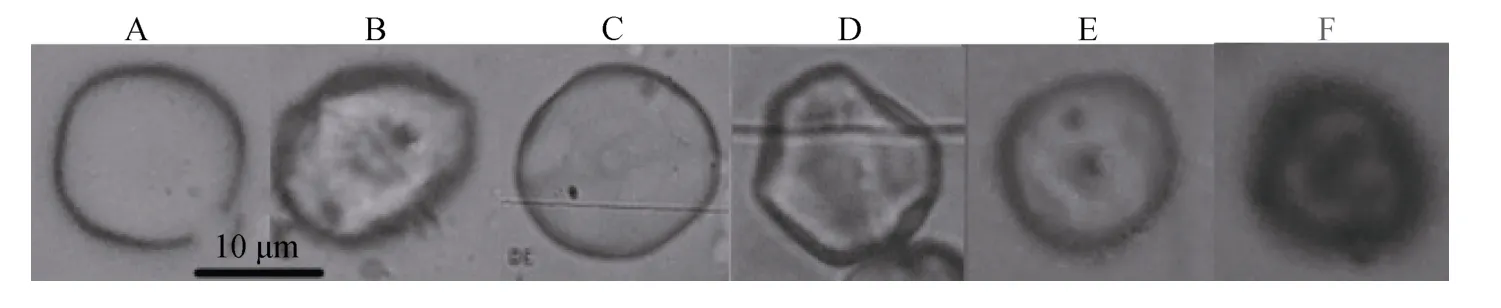
图5 长心卡帕藻 (A、B)和细齿麒麟菜 (C、D)酶解制备原生质体效果Fig.5 Protoplast of K.alvarezii (A and B)and E.denticulatum (C and D)
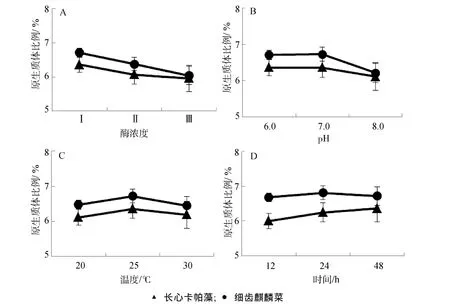
图6 不同酶解条件下原生质体占总细胞数的比例Fig.6 The proportion of protoplast number to total free cell
3 讨论
对不同制备温度(20~30℃)的试验,25℃酶解所制备的原生质体产量显著高于20℃和30℃酶解效果(图3),这可能与篮子鱼内脏研磨提取液在25℃时有更高的酶活性有关。基于此,我们在后续实验中,可在22~28℃范围内,优化酶解温度。
在上述各实验组的48 h酶解条件下,无论长心卡帕藻还是细齿麒麟菜的原生质体,其产量均随酶解时间的延长而增加,至少在 48 h酶解条件下,篮子鱼内脏粗提液仍具有酶的活性,延长酶解时间是否有效,有待后续深入研究。在3个酶解pH梯度实验(图4)中,pH 6.0时长心卡帕藻和细齿麒麟菜有更多原生质体的产出,间接证明其在pH 6.0时酶活性更好。
在藻泥与酶浓度配比实验中,酶解体系中长心卡帕藻或细齿麒麟菜藻泥含量越高,所制备的原生质体产量越高,而其产率却随藻泥浓度增加而降低(图 2);酶液用量较藻泥比率越高,原生质体产率越高(图2)。基于饲喂篮子鱼和从内脏中研磨提取酶没有制备藻泥方便,因此,可采用适当增大藻泥质量并延长酶解时间,以有效提高原生质体产量。
在藻泥与酶浓度配比实验中,酶解体系中长心卡帕藻或细齿麒麟菜藻泥含量越高,所制备的原生质体占总细胞数目的比例越低(图6)。这与原生质体产量随藻泥与酶浓度配比变化则相反,而与产率变化一致。由此可见,藻泥含量增大导致获得的游离细胞总数目增多和原生质体数目增多,但其并没有提高原生质体数目占总细胞数目的比例。在原生质体制备的温度梯度(20~30℃)中,25℃酶解所制备的原生质体数目占总细胞数目的比例显著高于 20℃和30℃酶解效果(图 6,P<0.01),其与原生质体产量随温度变化规律相一致。2种藻原生质体占总细胞数目的比例随pH变化规律与产量变化规律相似,pH为8.0时原生质体产量和原生质体所占的比例都低于pH为6.0和7.0时(图3;图6)。其可能是因为篮子鱼内脏研磨提取液中的酶组分在弱碱性的环境中酶活力较低。2种藻原生质体产量都随酶解时间延长而增加,其中长心卡帕藻原生质体占总细胞数目的比例随时间变化规律与产量结果一致,但细齿麒麟菜在酶解 24 h以后,原生质体占总细胞数目的比例不再增加(图 6.D),而仅仅是总的游离细胞数目增加导致原生质体产量的提高。
Salvador等[27]以鲍鱼内脏提取液获得长心卡帕藻原生质体,并在25℃和pH 6.1条件下,同样按0.1 g海藻材料加3 mL酶液的比例酶解48 h时,获得最高酶解效率,其最大原生质体产率为 8.2×103cell/g,这和我们的结果大致相同,所得提取液的效率(1.45×104Cell/g)的56%。Sahu等[28]将不同长心卡帕藻进行嫁接得到融合体植株,其生长速率更高,并且所产卡拉胶的黏度和强度都有提高。Cheney等[25]曾利用孢子制得红藻原生质体,并通过细胞融合的方法获得了江蓠细胞融合体,进而获得了生长速率更高的融合体植株。
本研究篮子鱼内脏粗提液酶解海藻藻泥的方法,成功制得了长心卡帕藻和细齿麒麟菜原生质体。同时,实验测定的了几种不同酶解条件对酶解反应的影响,建议按0.4 g藻泥中添加3 mL酶液的浓度配比,在25℃,pH 6.0的酶解条件下酶解48 h以更有效地获得海藻原生质体。
[1]Ohno M,Largo D B,Ikumoto T.Growth-Rate,carrageenan yield and gel properties of cultured kappa-carrageenan producing red alga Kappaphycusalvarezii (doty)doty in the subtropical waters of shikoku,Japan[J].Journal of Applied Phycology,1994,6(1): 1-5.
[2]Ask E I,Azanza R V.Advances in cultivation technology of commercial eucheumatoid species: a review with suggestions for future research[J].Aquaculture,2002,206(3-4): 257-277.
[3]Bindu M S ,Levine I A.The commercial red seaweed Kappaphycus alvarezii-an overview on farming and environment[J].Journal of Applied Phycology,2011,23(4): 789-796.
[4]Tye R J.Industrial and non-fooduses for carrageenan[J].Carbohydrate Polymers,1989,10(4): 259-280.
[5]Tobacman J K,Wallace R B,Zimmerman M B.Consumption of carrageenan and other water-soluble polymers used as food additives and incidence of mammary carcinoma[J].Medical Hypotheses,2001,56(5): 589-598.
[6]Pedersen J K.Carrageenan - functional properties and application in food[J].Cereal Science Today,1974,19(10): 471-476.
[7]Khambhaty Y,Mody K,Gandhi M R,et al.Kappaphycus alvarezii as a source of bioethanol[J].Bioresource Technology,2012,103(1): 180-185.
[8]Meinita M D N,Hong Y K,Jeong G T.Comparison of sulfuric and hydrochloric acids as catalysts in hydrolysis of Kappaphycus alvarezii (cottonii)[J].Bioprocess and Biosystems Engineering,2012,35(1-2): 123-128.
[9]Kraan S.Mass-cultivation of carbohydrate rich macroalgae,a possible solution for sustainable biofuel productionl[J].Mitigation and Adaptation Strategies for Global Change,2010,18(1): 27-46.
[10]庞通,刘建国,林伟,等.藻类生物燃料乙醇制备的研究进展[J].渔业现代化,2012,39(5): 63-71.
[11]刘建国,庞通,王莉,等.导致热带产卡拉胶海藻大规模死亡原因分析与藻株抗病差异性比较[J].海洋与湖沼,2009,40(2): 235-241.
[12]Mairh O P,Soehtun U,Ohno M.Culture of Eucheuma striatum (rhodopyta,solieriaceae)in subtropical waters of shikoku,Japan[J].Botanica Marina,1986,29(2): 185-191.
[13]Glenn E P,Doty M S.Photosynthesis and respiration of the tropical red seaweeds,Eucheuma striatum (tambalang and elkhorn Varieties)and E.denticulatum[J].Aquatic Botany,1981,10(4): 353-364.
[14]AzanzaCorrales R,Aliaza T T,Montano E.Recruitment of Eucheuma and Kappaphycus on a farm in tawi-tawi,Philippines[J].Hydrobiologia,1996,327: 235-244.
[15]Dawes C J.Ecological-studies of floridian Eucheuma(rhodophyta,gigartinales).IV.seasonal biochemical variations and reproduction[J].Bulletin of Marine Science,1982,32(1): 237-242.
[16]Azanzacorrales R,Mamauag S S,Alfiler E,et al.Reproduction in Eucheuma-Denticulatum (burman)collins and hervey and Kappaphycus-alvarezii (doty)doty farmed in danajon reef,Philippines[J].Aquaculture,1992,103(1): 29-34.
[17]Araki T,Hayakawa M,Tamaru Y,et al.Isolation and regeneration of haploid protoplasts from Bangia-atropurpurea (rhodophyta)with marine bacterial enzymes[J].Journal of Phycology,1994,30(6): 1040-1046.
[18]Chen C S,Shyu J F.Regeneration of protoplasts from the brown alga,Endarachne-binghamiae (phaeophyta,punctariales,scytosiphonaceae)[J].Botanical Bulletin of Academia Sinica,1994,35(3): 189-193.
[19]Fujimura T,Kawai T,Kajiwara T,et al.Protoplast Isolation in the marine brown alga Dictyopteris prolifera(dictyotales)[J].Plant Cell Reports,1995,14(9): 571-574.
[20]Gross W.Preparation of protoplasts from the carrageenophyte Gigartina-corymbifera [J].Journal of Microb-iological Methods[J]1990,12(3-4): 217-223.
[21]Linz K W,Kohler K.Isolation of protoplasts from the coccal green-alga Eremosphaera-viridis debary for patch-clamp measurements[J].Botanica Acta,1993,106(6): 469-472.
[22]Mejjad M,Loiseauxdegoer S ,Ducreux G.Protoplast lsolation,development,and regeneration in different strains of Pilayellalittoralis (L)kjellm (Phaeophyceae)[J].Protoplasma,1992,169(1-2): 42-48.
[23]Reddy C R K,Gupta M K,Mantri V A,et al.Seaweed protoplasts: status,biotechnological perspectives and needs[J].Journal of Applied Phycology,2007,20(5): 619-632.
[24]Yeong H-Y,Khalid N,Phang S-M.Protoplast isolation and regeneration from Gracilaria changii (Gracilariales,Rhodophyta)[J].Journal of Applied Phycology,2008,20(5): 641-651.
[25]Cheney D,Rudolph B,Wang L Z,et al.New Developments in Marine Biotechnology [M].New York:Springer-Verlag New York Inc,1998: 101-104.
[26]Zablackis E,Vreeland V,Kloareg B.Isolation of protoplasts from Kappaphycus alvarezii var tambalang(rhodophyta)and secretion of iota-carrageenan fragments by cultured-cells[J].Journal of Experimental Botany,1993.44(266): 1515-1522.
[27]Salvador R C,Serrano A E.Isolation of protoplasts from tissue fragments of Philippine cultivars of Kappaphycus alvarezii (Solieriaceae,Rhodophyta)[J].Journal of Applied Phycology,2005.17(1): 15-22.
[28]Sahu N,Ganesan M,Eswaran K.Inter- and intrageneric grafting in seaweeds in the Indian coasts[J].Current Science,2010.99(2): 235-239.
