平疣桑椹石磺精子结构观察
2014-03-25钱静沈和定王成暖
钱静, 沈和定, 王成暖
(上海海洋大学 水产与生命学院,上海 201306)
精子是生物发生发育的基础,是受精生物学中的重要研究内容。研究精子形态结构有助于受精机制的深入研究,促进人工繁殖工作的顺利开展。此外,由于动物精子形态结构具有种的特异性,可在一定程度上为研究物种间亲缘关系和系统分类提供参考依据。精子头部顶体的大小与形态,以及亚顶体腔的有无及其形态结构等均可作为种间鉴别的依据。精核的形态,大小以及横径与纵径的比例,核前窝与核后窝的有无及其结构差异,精子中段线粒体数量及排列方式也是研究系统演化的重要依据,比较外形相似种的精子超微结构之间的差异可为软体动物的系统分类工作提供参考资料[1]。迄今,国内外学者对腹足类软体动物精子超微结构已有较多研究[2-5]。
石磺科贝类隶属软体动物门(Mollusca)、腹足纲(Gastropoda)、肺螺亚纲(Pulmonata)、缩眼目(Systellommatophora)、石磺科(Onchidiidae)。广泛分布于世界各地,主要生活于沿海潮间带高潮区。目前国内有关石磺科贝类的研究主要集中于形态分类,内部解剖、人工繁殖及胚胎发育等方面[6-10]。对于石磺科贝类精子结构的研究较为匮乏,仅应雪萍等[11]研究了瘤背石磺(Onchidiumstruma)的精子超微结构,其余种类石磺精子结构至今未见报道。本文研究了平疣桑椹石磺(Platevindexmortoni)精子超微结构,为其繁殖生物学及相关基础研究积累资料;同时与瘤背石磺精子形态进行了比较,为石磺科分类工作提供参考。
1 材料和方法
1.1 实验材料
平疣桑椹石磺于2013年5—6月采自广东湛江,样品采集后立即带回实验室暂养。挑选活力好,成熟度高的个体置冰块上进行活体解剖,迅速分离出生殖腺。
1.2 实验方法
1.2.1 光镜标本制作
解剖分离出两性腺和支囊,切成1 mm3的小块,放入Bouin氏液中常温固定24 h。酒精梯度脱水,二甲苯透明,石蜡包埋(56~58℃),Leica RM2016型切片机连续切片,切片厚度6 μm,苏木素和伊红双重染色,中性树胶封藏。另外取少量新鲜的成熟两性腺和支囊,直接在载玻片上涂片,染色,Nikon-50i电光源显微镜下观察、拍照。
1.2.2 透射电镜标本制作
将迅速分离出的两性腺和支囊切成0.5 mm见方的小块,用 0.1 mol/L、pH值7.4的磷酸缓冲液配制的3%戊二醛-1.5%多聚甲醛前固定, 4℃保存备用。固定样品经磷酸缓冲液充分漂洗,再用1%锇酸后固定2 h,酒精-丙酮系列脱水,Epon812环氧树脂浸透包埋,LKB-8800V超薄切片机切片, 切片厚度70 nm,醋酸铀-柠檬酸铅双重染色,日立H-7700型透射电镜下观察并拍照。
2 结果
2.1 光镜的观察
平疣桑椹石磺属雌雄同体贝类,两性腺滤泡既可产生精子也可产生卵子,属于滤泡混合型,精卵发育不同时(图1-1)。支囊中充满精子,H.E染色后,精子头部细胞核区蓝色(图1-2)。两性腺中的精子成簇分布,头部为球形(图1-3),精子束输送至支囊后自动散开,头部变成梭形或直线形(图1-4)。
2.2 透射电镜的观察
平疣桑椹石磺精子为典型的进化型精子,包括头部、中段和末段。
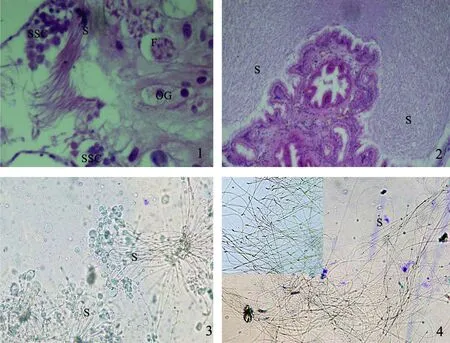
图1 平疣桑椹石磺精子发育的形态结构
1.两性腺切片,示腺泡、次级精母细胞、成熟精子与卵原细胞 ×200;2.支囊中精子形态 ×400;3.两性腺中精子束,示球形头部 ×400;4.支囊中精子散开,头部呈梭形 ×400。SSC—次级精母细胞;S—精子;F—腺泡;OG—卵原细胞。
2.2.1 头部
头部长约4.5 μm,纵切面形态似辣椒,由顶体(acrosome)和精核(nucleus)构成。
顶体位于头部最前端,约为0.7 μm,似奶嘴状,电子密度小且均匀。顶体基部内凹形成圆弧形,与核前端相连,几乎看不到间隙,外有质膜包被(图2-1)。
精核与顶体后端紧密相连,长约3.8 μm,宽约0.8 μm至1.2 μm,染色质浓缩且均匀,电子密度较高,透射电镜下观察未发现核泡的存在。细胞核前端平整,未形成核前窝(front nuclear pocket ),但核后端内凹形成核后窝( posterior nuclear pocket )(图2-2)。精子头部与质膜间往往有少量的细胞质,质膜(plasma membrane)呈波纹状(图2-3)。
2.2.2 中段
平疣桑椹石磺精子的中段紧接于核的后端,结构较为复杂。包括线粒体、中心粒、质膜、类晶体物质和糖原质螺旋体等。
核后窝内包含一对相互垂直的中心粒,与多数腹足纲贝类相似,近端中心粒(proximal centriole)长轴与精子长轴垂直,靠近细胞核;远端中心粒(distal centriole)长轴与精子长轴方向一致(图2-4)。通过精子前中段横切面可清晰观察到一圈由圆形颗粒构成的类晶体层包裹着轴丝(axoneme) ,图2-5。类晶体层外层为基质层,纵切面观基质层较厚(图2-6)。
糖原质螺旋体(glycogen helix)内含许多颗粒物质,电子密度较高(图2-7、8)。纵切面观,糖原质螺旋体呈细长囊泡状,过精子前中段横切面呈两个圆形,其中一个糖原质螺旋体随中段往后逐渐变小,剩下的一个糖原质螺旋体过中后段横切面呈半圆形或弯月形(图2-9、10)。
过精子中段纵切面可清晰看见线粒体演化成线粒体鞘,螺旋状包绕轴丝(图2-11、12)。
2.2.3 末段
平疣桑椹石磺精子末段结构较为简单,无糖原质螺旋体和线粒体,横切面呈圆形,能清晰看到内部轴丝呈典型的“9+2”结构(图2-13)。
3 讨论
3.1 腹足纲贝类精子超微结构的比较
腹足纲贝类精子分为原始型和进化型两种。原始型精子头部呈锥形,中段较短,有4~5个线粒体,无线粒体鞘(chondriosomal mantle)结构;进化型精子头部呈线形,中段加长,线粒体演化成线粒体鞘结构包围中心粒排列[12]。通过比较9种腹足纲贝类的精子超微结构,发现不同科贝类精子形态结构差异较大,如表1所示。
原始腹足类的角蝾螺(Turbocornutus)顶体呈啤酒瓶状[13];塔形马蹄螺(Trochuspyramis)顶体呈圆锥形[4];鲍类顶体呈圆锥状或弹头状;而新腹足目的香螺(Npatunedecumingi)[12]则无顶体结构。可见,不同种贝类精子的顶体发达程度及形态结构均存在较大差异。
原始腹足目角蝾螺和塔形马蹄螺精核呈长柱形,均有明显的核前窝与核后窝结构[4, 13];皱纹盘鲍(Haliotisdiscushannai)的精子核呈长柱状,核前窝凹陷较深呈V字型,但核后端凹陷较浅[14]。肺螺类的瘤背石磺精子[11]与本文研究的平疣桑椹石磺精子均有明显核后窝却无核前窝。可见精核除了形态结构差异外,核前窝与核后窝的有无也是作为种间区别的重要依据。
原始腹足目的角蝾螺[13]、塔形马蹄螺[4]与盘鲍(Haliotisdiscus)精子中段有5个线粒体;皱纹盘鲍与杂色鲍(Haliotisdiversicolorsupertexta)精子中段有5~6个线粒体[14]。新腹足目的疣荔枝螺(Purpuraclavigera)的精子中段线粒体均螺旋化,肺螺亚纲的瘤背石磺精子与本文研究的平疣桑葚石磺精子中段线粒体均为5个,螺旋化形成线粒体鞘包绕在轴丝外。

表1 9种腹足纲贝类精子超微结构比较
3.2 同科不同种贝类精子结构差异比较
对比发现同科不同种贝类的精子形态差异不是很大,但在体尺参数、顶体形态、精核形态、线粒体数目等方面存在或大或小的差异。例如,柯才焕等[14]研究的3种鲍精子的超微结构发现鲍类精子形态的基本结构是相似的,但顶体和核的尺寸及外形上有着或大或小的差异。盘鲍与皱纹盘鲍的精子在外形较为相似,却明显长于杂色鲍。盘鲍精子顶体长度大于皱纹盘鲍大于杂色鲍,杂色鲍的顶体呈圆锥状,顶端较尖,而皱纹盘鲍和盘鲍的顶体呈圆柱状,顶端较钝。盘鲍顶体下腔的内凹较深,杂色鲍次之,皱纹盘鲍最浅。

图3 瘤背石磺与平疣桑椹石磺头部纵切示意图
1.瘤背石磺精子头部纵切;2.图右为平疣桑椹石磺精子头部纵切。A—顶体;N—精核;PM—质膜;PC—近端中心粒;DC—远端中心粒;Nv—核泡。
对比瘤背石磺精子结构与本文研究的平疣桑椹石磺精子结构发现两种精子中段和末段均相似,但头部差异较大(图3)。瘤背石磺顶体0.8 μm,与核前端有间隙,精核4.2 μm,前端较窄约0.29 μm,后端较宽约0.96 μm;而平疣桑葚石磺顶体0.7 μm,与核前端无间隙,精核长约3.8 μm,前后端直径相差不大,前端稍窄,宽度约为0.8 μm,后端宽约1.2 μm。瘤背石磺精子顶体呈典型奶嘴状,顶体囊内电子密度稍低于顶体构架体;平疣桑椹石磺精子顶体奶嘴状形态不明显,顶体囊与顶体构架体内电子密度无明显差异。两种石磺精子间最明显的区别是瘤背石磺精核存在一些大小不等的核泡,而平疣桑椹石磺精核电子密度高,未发现核泡的存在。有学者推测精核中的囊泡可能携带了由线粒体释放出的能量,由核区向顶部运动,起到能量运输或传递的作用[1]。关于核泡的来源及其意义仍待进一步研究。
参考文献:
[1]焦海峰,尤仲杰,竺俊全. 软体动物精子的超微结构及其分类学意义[J]. 宁波大学学报:理工版, 2005, 17(4): 414-418.
[2]Jörger K M, He B M, Neusser T P, et al. Sex in the beach: spermatophores, dermal insemination and 3D sperm ultrastructure of the aphallic mesopsammic Pontohedyle milaschewitchii (Acochlidia, Opisthobranchia, Gastropoda)[J]. Marine Biology, 2009, 156(6): 1159-1170.
[3]Schilthuizen M, van Heuven B. Dextral and sinistral Amphidromus inversus (Gastropoda: Pulmonata: Camaenidae) produce dextral sperm[J]. Zoomorphology, 2011, 130(4): 283-287.
[4]吴洪流,蒲利云,冯永勤,等. 塔形马蹄螺精子发生的超微结构研究[J]. 热带生物学报, 2010, 1(1): 21-26.
[5]Cuezzo M G. Ultrastructure of spermatozoa in Orthalicidae (Mollusca, Gastropoda, Stylommatophora) and its systematic implications[J]. Zoomorphology, 2011, 130(3): 147-166.
[6]吴旭峰,沈和定,吴文健,等. 中国华东沿海 4 种石磺形态学比较[J]. 动物学杂志, 2010, 45(6): 92-100.
[7]金春华,郑忠明,华建权,等. 瘤背石磺人工繁育关键技术的研究[J]. 水产科学, 2010, 29(10): 597-600.
[8]陈美英,李建榜,黄克蚕,等. 瘤背石磺的生殖系统结构特点及生殖周期研究[J]. 四川动物,2010(3): 426-431.
[9]沈永龙,黄金田,戈贤平,等. 几种重要环境因子对瘤背石磺人工养殖成活率的影响[J]. 南方水产科学,2012, 8(6): 57-64.
[10]沈永龙,黄金田,戈贤平,等. 不同孵化方式, 培育密度和附着基对瘤背石磺人工繁育的影响[J]. 海洋科学, 2013, 37(3): 109.
[11]应雪萍,孙胜安,陈美英. 瘤背石磺的精子结构 3[J]. 动物学报, 2008, 54(2): 373-382.
[12]侯 林,高 岩,邹向阳,等. 香螺精子发生及精子超微结构[J]. 动物学报, 2006, 52(4): 746-754.
[13]竺俊全,杨万喜. 角蝾螺精子的超微结构[J]. 浙江大学学报:理学版, 2006, 32(6): 685-690.
[14]柯才焕,周时强,田 越,等. 三种鲍精子超微结构的比较[J]. 海洋学报, 2003, 25(3): 138-142.
[15]竺俊全,杨万喜,焦海峰. 疣荔枝螺精子的超微结构[J]. 水产学报,2006, 29(5): 643-648.
