可口革囊星虫体腔中卵细胞大小组成的周年变化
2014-03-23龙玲利曾海祥丁理法竺俊全
刘 懂, 龙玲利, 曾海祥, 丁理法, 竺俊全
(1. 宁波大学 教育部应用海洋生物技术重点实验室, 浙江 宁波 315211;2. 温岭市水产技术推广站, 浙江 温岭 317500)
可口革囊星虫(Phascolomaesculenta)隶属于星虫动物门、革囊星虫纲、革囊星虫目、革囊星虫科,自然分布于中国东南沿海[1]。由于其风味独特,食用、营养及药用价值较高,增养殖发展前景看好,育苗与养殖及其生物学研究倍受重视,已见人工繁殖与养殖技术[2-3]、生殖与发育[4-8]、消化系统结构[9]、生化与分子[10-11]等方面的研究报道。该种动物的性腺非常细小,以致肉眼难辨;雌性个体的卵原细胞在卵巢中发育到初级卵母细胞早期阶段落入体腔中,悬浮在体腔液中生长发育[12];雄性个体的精原细胞在精巢中发育到精细胞早期以细胞团形式脱离精巢入体腔液中继续发育[6]。虽然,陈慧等[12]研究过福建宁德海区潮间带可口革囊星虫的生殖周期,但因地理分布的不同,浙江沿海可口革囊星虫的生殖周期与福建宁德的存在差异。体腔中卵细胞大小组成的年变化能反映星虫的生殖周期,因此,本研究在浙江温岭沿海每月采样,显微观察了性成熟可口革囊星虫体腔液中卵细胞大小组成的周年变化,旨在了解其生殖周期,确定繁殖期,为其苗种繁育及养殖提供指导。
1 材料和方法
1.1 材料
可口革囊星虫取自浙江温岭沿海潮间带,一周年中每月中旬采样1次,每次采集100多条,体质量1.5~2.5 g/条。每次采得的样品均带回宁波大学实验室暂养,供实验用。
1.2 方法
1.2.1 体腔中生殖细胞的观察和测量
从每次采得的样品中随机挑选40条进行解剖,用吸管吸取体腔液,用10倍过滤海水稀释后涂片,在显微镜下观察生殖细胞的特征,区分雌雄;用目微尺测量雌体卵细胞大小,每条虫随机测30个卵细胞的直径,当卵由圆形变为椭圆形以后,测其长径。
取部分雌性个体的体腔液,静置沉淀后获得体腔细胞(内含卵细胞),将体腔细胞用Bouin 氏液固定,常规石蜡切片,HE染色,显微观测与拍照。
1.2.2 数据处理
应用Excel对卵细胞数据进行分析并作图,以显示卵细胞组成的年变化情况。
2 结果
2.1 雌雄鉴别及性比
可口革囊星虫雌雄异体,外形上难以区别雌雄,只有解剖观察才能鉴别。非繁殖季节,雌体的体腔液中含有卵细胞(图1:a),雄体体腔液中含有精细胞团(图1:b)。繁殖季节,雌性个体的体腔液由于含有大量的接近成熟的卵细胞而呈青灰色,雄性个体体腔液因为有大量的精细胞团而呈乳白色。精卵发育成熟后由体腔进入肾管储存,因而雌体的肾管呈青灰色或青褐色(图2:a),雄体肾管呈乳白色(图2:b)。从3月到翌年2月,我们共解剖、显微观察了480条可口革囊星虫,其中雌体250条,雄体210条,无法判定性别的为20条,雌雄比接近1∶1。

图1 可口革囊星虫体腔液中的卵细胞(♀)及精子细胞团(♂)Fig 1 Oocytes(♀) and spermatid masses(♂) in the coelomic fluid of P.esculenta
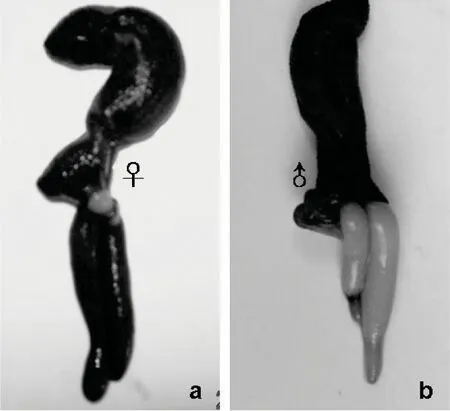
图2 可口革囊星虫肾管充满卵子(♀)及精子(♂)Fig 2 Oocytes(♀) and sperm(♀) filling the nephridia of P.esculenta
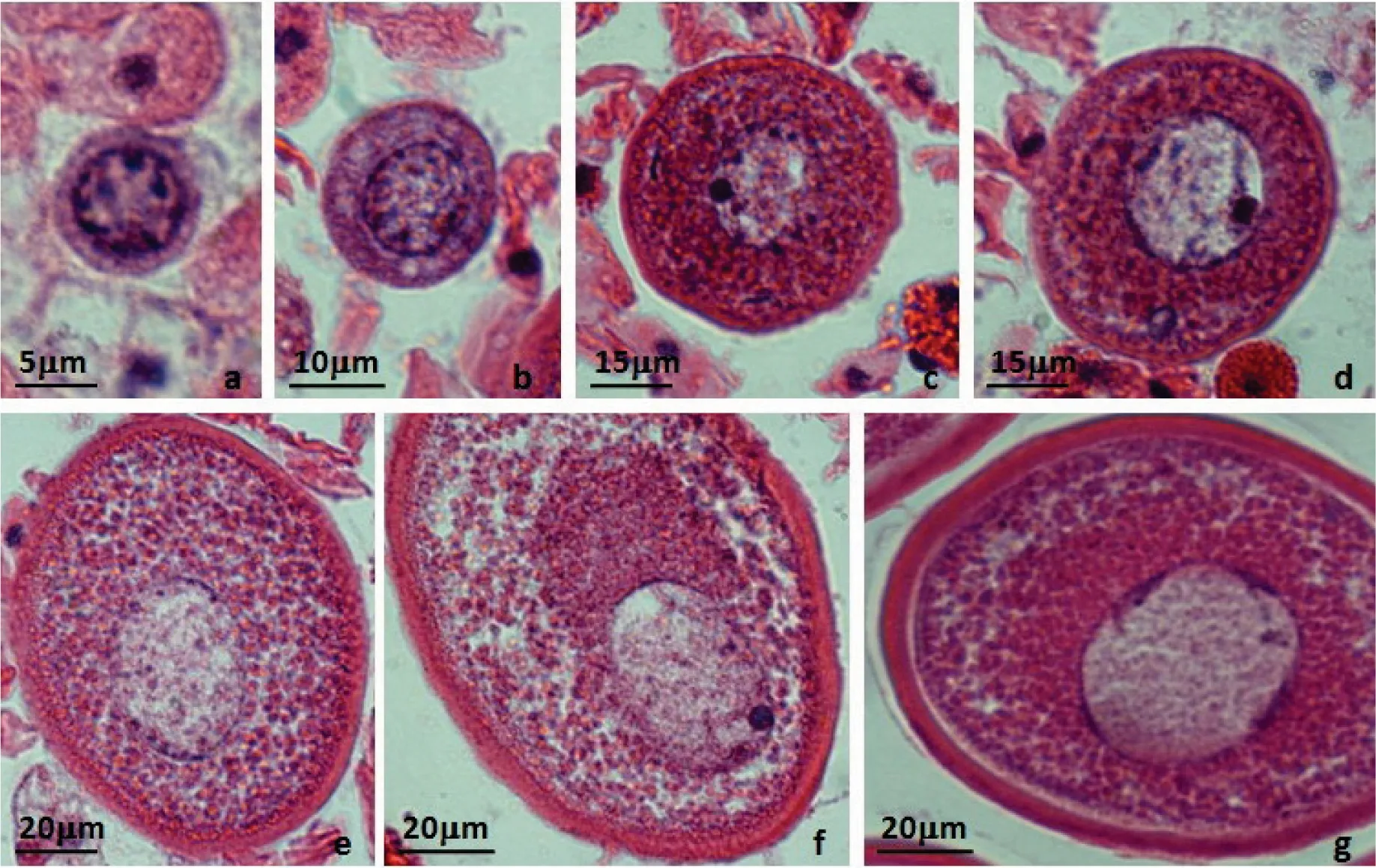
图3 可口革囊星虫体腔中卵细胞的形态特征
a—小型卵细胞,卵径约10 μm;b—小型卵细胞,卵径约20 μm;c—中型卵细胞,卵径约50 μm;d—中型卵细胞,卵径(长径)约70 μm;e—大型卵细胞,卵径(长径)约110 μm;f—大型卵细胞,卵径(长径)约130 μm;g—大型卵细胞,卵径(长径)约140 μm。
2.2 体腔中卵细胞的形态特征
可口革囊星虫雌性成体体腔中存在不同大小的卵细胞,卵径小于60 μm的一般呈圆形,大于60 μm的一般呈卵圆形或椭圆形。最小的卵细胞卵径约10 μm,细胞质少,核大而圆(图3:a);卵径约20 μm的小型卵细胞胞质明显增加,核质比下降(图3:b);卵径约50~70 μm的中型卵细胞质膜外的卵黄膜明显,核呈圆形,核仁明显,卵质中卵黄较多(图3:c,d);卵径约110~130 μm的大型卵细胞卵黄膜厚达6~7 μm,核呈椭圆形,卵质中卵黄丰富(图3:e,f);卵径约140 μm以上的大型卵细胞达生长成熟,体积不再增大,卵黄膜厚达7~9 μm,核呈椭圆形,卵质中充满卵黄(图3:g)。
2.3 体腔中卵细胞大小组成的周年变化
以横轴为卵细胞的直径(长径),纵轴为卵细胞的百分比,显示体腔中各月卵细胞大小组成如图4。3—4月,大部分卵细胞卵径分布在30~90 μm之间,50~60 μm处形成明显波峰,100 μm以上的卵细胞很少。5月,卵径30~90 μm的卵细胞仍占大部分,100 μm以上的卵细胞明显增多。6月,卵细胞卵径主要分布在70~130 μm之间,其中110~120 μm的卵细胞最多,130 μm以上的卵细胞很少。7月,大部分卵细胞卵径在100 μm以上,其中120~130 μm的卵细胞占比例最高,130~150 μm的卵细胞明显增多,约占1/5。8月,卵细胞卵径均在110 μm以上,其中130~150 μm的卵细胞约占2/3。9月,卵细胞卵径仍多数在110 μm以上,其中110~130 μm的卵细胞占大部分,130~150 μm的卵细胞约占1/3。10月,以110~140 μm的卵细胞占多,40 μm以下的卵细胞较少,130~150 μm的卵细胞约占30%,但这些大卵细胞多数已经开始进入退化期。11月至翌年2月,随着大型卵细胞的逐渐退化,小型及中型卵细胞逐渐增多;1~2月在40~50 μm处形成波峰,并仍可见少量未退化的大型卵细胞。
以横轴为月份、纵轴为卵细胞月平均卵径,显示卵细胞平均大小的年变化情况如图5。卵细胞月平均卵径在3—8月逐渐增大,8月最大,8月以后逐渐减小,其中3—5月及6—8月增大较慢,5—6月增大较快,8—12月减小较快,12—2月减小较慢。

图4可口革囊星虫体腔卵细胞大小组成的年变化
Fig 4 Monthly change of oocyte size in the coelom ofP.esculenta
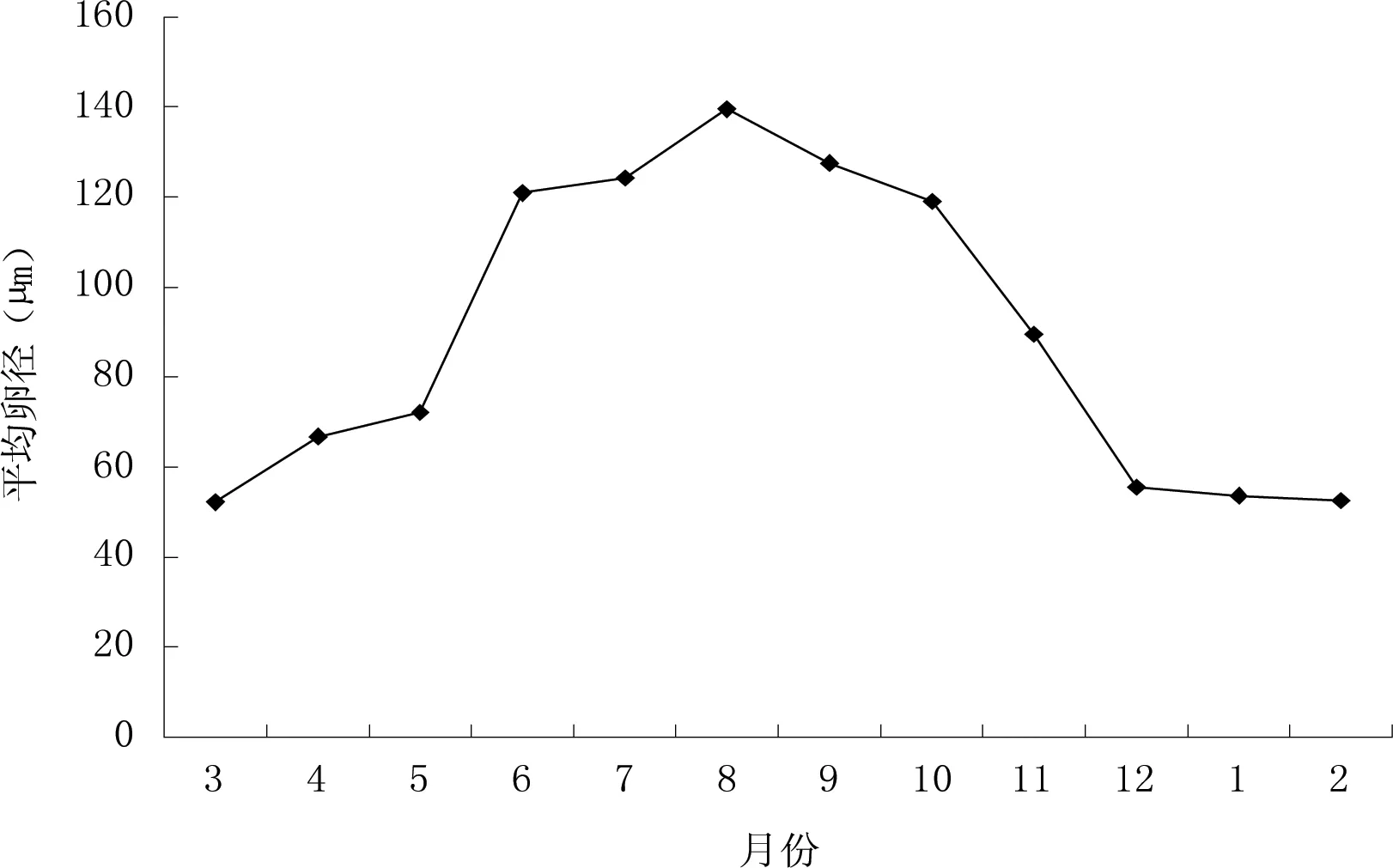
图5 可口革囊星虫体腔中卵细胞平均大小的年变化
3 讨论
星虫类动物体腔中的卵细胞大小组成随季节变化而发生周期性变化。Gonse研究了Golfingiavulgaris星虫的生殖周期,发现体腔中小型卵细胞全年存在,并于产卵期以后出现几个月生长停滞;中型卵细胞在产卵季节和产卵后会消失[13]。郭学武[14]对方格星虫生殖周期的研究也得到了和Gonse相同的结果。Rice[15]研究发现Golfingiapugettensis星虫的小型卵细胞常年存在,产卵期间出现生长停滞,产卵后重新生长;繁殖季节,中型卵细胞基本消失,只存在大型卵细胞和小型卵细胞。Towle[16]对美国加州蒙特利海湾的Phascolosomaagassizii星虫进行了研究,发现小卵细胞在7月出现,随着季节的推移,卵细胞不断增大,数量不断增加,翌年3月体腔中全部为大型卵细胞。我们的研究表明,可口革囊星虫的小型卵细胞一般在10月出现,此后小卵细胞逐渐长大,到次年5—6月迅速生长,7—9月大多数成为大型卵细胞。
星虫的繁殖季节因种类及地理分布的不同存在差异。Rice[15]研究了阿根廷圣胡安沿海的3种星虫的生殖周期,发现Phascolosomaagassizii星虫的繁殖季节在6月中旬到9月上旬,Themistepyroides星虫的繁殖季节在3月至8月,Golfingiapugettensis星虫的繁殖季节在10月至翌年1月。Towle[16]研究发现美国加州海岸的Phascolosomaagassizii星虫繁殖季节在3—5月。郭学武等[14]研究认为方格星虫厦门种群的产卵季节为5—9月,兰国宝[17]和王庆恒[18]分别对方格星虫北海和湛江种群的研究也得到类似结论。陈慧等[12]报道的福建宁德沿海的可口革囊星虫繁殖季节为5~9月,其中7—8月为繁殖盛期。本研究的浙江温岭的可口革囊星虫在7~9月期间,各月均有部分卵细胞卵径大于140 μm。根据顾晓英等[4]对可口革囊星虫卵子发生的组织学观察结果,卵径达140~145 μm的为生长成熟的卵细胞,在外界条件适宜的情况下,这些卵细胞能短时间内生理成熟,并通过肾管产出体外。根据竺俊全等[6]对可口革囊星虫精子发生的研究结果,4—5月雄性个体体腔中精细胞团逐渐增多;6—9月精细胞团几乎充满体腔,在适宜的外环境因子诱导下精细胞能短时间内发育为成熟精子,并经肾管排出体外;10—11月残留的精细胞团逐渐退化;12—3月体腔中只留下少量精细胞团。作者育苗生产试验发现,7—9月人工催产能获得成功。因此,推断可口革囊星虫在浙江温岭的繁殖期主要在7—9月。
一般认为星虫的繁殖盛期多在夏季,这可能是由于夏季的水温较高,有利于星虫的胚胎和幼体发育。Field[19]认为夏季海水中浮游生物丰富,可以为浮游幼虫提供丰富的饵料,提高幼体的存活率。
参考文献:
[1]李风鲁,周 红, 等.中国沿海星虫动物门名录[J].青岛海洋大学学报, 1992, 22(2):72-87.
[2]吴洪喜,陈 琛,曾国权,等.可口革囊星虫人工繁殖试验[J].海洋科学,2010,34(3):21-25.
[3]徐敏娴,丁理法,周敏华.可口革囊星虫人工繁育及增养殖技术的研究[J].水产养殖,2011,32(11):39-41.
[4]顾晓英,竺俊全,许式见,等.可口革囊星虫(Phascolosomaesculenta)卵子发生的组织学研究[J].海洋与湖沼,2009, 40(3): 283-288.
[5]金春华,竺俊全,许式见,等.可口革囊星虫(Phascolosomaesculenta)胚胎及幼虫发育研究[J].海洋与湖沼,2011,42(1): 94-100.
[6]竺俊全,王 武,许式见,等.可口革囊星虫精子发生及精子结构[J].动物学报,2007,53(4):733-741.
[7]竺俊全,王 武,应雪萍,等.可口革囊星虫受精过程及早期卵裂的细胞学变化[J].动物学报,2008,54(2): 290-298.
[8]竺俊全,王 伟,丁理发.可口革囊星虫(Phascolosomaesculenta)卵黄合成期卵母细胞发育及卵黄发生与卵膜形成的超微结构[J].海洋与湖沼,2012,43(4): 870-876.
[9]雷世勇,卢明明,丁理法,等.可口革囊星虫消化道的形态结构特征[J].生物学杂志,2013,30(2):33-36(39).
[10]雷世勇,丁理发,黄福勇,等.可口革囊星虫不同组织同工酶的比较[J].生物学杂志,2013,30(4):11-14.
[11]Su X R, Du L L, Li Y Y, et al. Cloning and expression of HSP70 gene of sipunculaPhascolosomaesculenta[J]. Fish & Shellfish Immunology, 2010,28(3): 461-466.
[12]陈 慧, 林国文,陈 武,等.可口革囊星虫生殖周期的观察[J].海洋渔业,2009,31(2):139-145.
[13]Gonse P. Lovogenese chezPhascolosomavulgaraII. Recherches biometriques sur les ovocytes[J]. Acta Zool, 1956, 37:225-233.
[14]郭学武,李复雪.光裸星虫生殖周期的研究[J].热带海洋,1993,12(2):69-76.
[15]Rice M E. Gametogenesis in three species of Sipuncula:Phascolosomaagassizii,GolfingiapugettensisandThemistepyroides[J].Cellule, 1974, 70(2):295-313.
[16]Towel A, Giese A C. The annual reproductive cycle of the sipunculidPhascolosomaagassizii[J]. Phsiol Zool, 1967, 40:229-237.
[17]兰国宝,阎 冰.方格星虫繁殖生物学研究[J].水产学报,2002,6(26):503-509.
[18]王庆恒,杜小东,黄洪艳.湛江地区光裸星虫的生殖细胞发育和生殖周期[J].湛江海洋大学学报,2005,25(1):5-9.
[19]Field W G. The structure, development, food relations, reproduction and life history of the squidLoligoopalescensBerry[J]. Fisheries Bull, 1965, 131:1-108.
