高等植物赤霉素生物合成及其信号转导途径
2014-03-21李强吴建明梁和黄杏丘立杭
李强吴建明梁和黄杏丘立杭
(1.广西大学农学院,南宁 530004;2. 中国农业科学院甘蔗研究中心 农业部广西甘蔗生物技术与遗传改良重点实验室,南宁 530007)
高等植物赤霉素生物合成及其信号转导途径
李强1,2吴建明2梁和1黄杏2丘立杭2
(1.广西大学农学院,南宁 530004;2. 中国农业科学院甘蔗研究中心 农业部广西甘蔗生物技术与遗传改良重点实验室,南宁 530007)
赤霉素是一种重要的植物激素,调节植物生长和发育的各个阶段,如促进种子萌发、茎杆伸长、叶片展开、花的发生及果实与种子的发育。综述了赤霉素合成、信号转导途径、与其他植物激素间的相互作用、对环境信号的响应以及DELLA泛素化降解过程的研究进展,这将有助于人们对赤霉素生理作用和分子调节机制的了解,有利于对赤霉素各方面的机理进行深入地研究。
赤霉素 信号转导 植物激素 调控机理
赤霉素(Gibberellins,GAs)属于一种四环双萜类植物激素,广泛分布于被子植物、裸子植物、蕨类植物、褐藻、绿藻、真菌和细菌中。目前已知有136种天然赤霉素[1],但在植物体内只有少数GA分子(如GA1、GA3、GA4和GA7等)具有生物活性,它们可调节和控制生物体生长发育的各个阶段,例如促进种子萌发、茎杆伸长、叶片展开、花的发生以及果实与种子的发育[2-6]。自20世纪60年代起,由于水稻sd1基因和小麦Rht1基因在育种中的大规模推广应用使世界主要粮食作物产量极大幅度地提高,这一历程即为众所周知的“绿色革命”。研究表明主要农作物的“绿色革命”都与赤霉素密切相关[7]。赤霉素参与调节植物生物发育中一个突出的特点是促使茎的伸长和植株增高,其效应包括:(1)GA能增加一些植物(如莲座天仙子)的细胞分裂,它能促进细胞分裂是因为缩短了细胞周期G1期和S期的时间。(2)GA可通过提高木葡聚糖内转糖基酶(XET)相关基因的转录水平,增加细胞壁组成成分木葡聚糖,促使细胞伸长。(3)GA促使微管的排列方向与生长着的细胞的长轴垂直。在缓慢生长的节间中,其居间分生组织以上的细胞内微管方向发生倾斜,因而有利于细胞伸长[8]。赤霉素合成途径
中的关键酶在拟南芥、水稻、南瓜、小麦、木薯和甘蔗等植物已经被克隆并进行表达及功能分析[9-15]。但是赤霉素生物合成过程极其复杂,参与的酶和影响其合成的因素很多,要深入了解GA信号传导途径尚需更深入的研究。
1 赤霉素的生物合成
赤霉素主要在高等植物的顶端幼嫩部位合成,如茎尖和根尖,以及生长中的种子和果实中。成熟的叶片也可以合成赤霉素,但很少输出。赤霉素在细胞中的合成部位是质体、内质网和细胞质基质等处[1,16,17]。科学家经过几十年的研究,现已基本确定高等植物赤霉素的生物合成途径(图1)。
根据赤霉素在细胞内不同的合成部位,其途径大致分为3个阶段:第1阶段在质体中进行:由牻牛儿牻牛儿基焦磷酸(Geranylgeranyl diphosphate,GGDP)为赤霉素生物合成的前体,经古巴焦磷酸合成酶(Ent-copalyl diphosphate synthase,CPS)和内根-贝壳杉烯合成酶(Ent-kaurene synthase,KS)催化下环化为赤霉素的前身内根-贝壳杉烯(Entkaurene);第2阶段在内质网中进行:内根-贝壳杉烯的C-19的甲基在内根-贝壳杉烯氧化酶(Ent-kaurene oxidase,KO)催化下不断被氧化,分别形成内根-贝壳杉烯醇(Ent-kaurenol)、内根-贝壳杉烯醛(Entkaurenal)和内根-贝壳杉烯酸(Ent-kaurenoic acid,KA)。KA在内根-贝壳杉烯酸氧化酶(Ent-kaurenoic acid oxidase,KAO)的催化作用下,在C-7α位上进行3步脱氢氧化反应,逐步形成内根-7α-羟基贝壳杉烯酸和GA12-醛,它是GA的最初产物,进一步转化成GA12,而在GA13氧化酶的作用下还可转变为GA53;第3阶段在细胞质基质中进行:由内质网合成的GA12和GA53运输至细胞质基质中,在其C20处经GA20氧化酶、GA3氧化酶和GA2氧化酶进行一系列氧化作用下转变为其他种类GAs。
2 赤霉素生物合成途径的调控
赤霉素的生物合成与代谢是多种酶参与的多步骤酶促反应过程,植物细胞通过对这些基因表达的精确调控来调节赤霉素合成与代谢速率[19]。GA作为调控植物生长发育的重要激素之一,它的合成在时间和空间上都受到严格的控制,许多因素都直接或间接参与GA合成调控。在植物体内GA可以通过前馈或反馈来调节GA合成途径的许多关键酶,控制赤霉素代谢[7]。赤霉素生物合成调节主要发生在转导途径后期,催化GA12转变为GA4相关的酶有GA20-oxidase和GA3-oxidase。GA2-oxidase能将有生物活性赤霉素催化为无活性,拟南芥有5个GA2-oxidase能钝化C19赤霉素的生物活性[20]。
到目前为止,科学家们对植物GA生物合成和代谢途经的调控领域研究相对成熟,主要集中于逆境、发育阶段、光、温度、活性GAs水平和其他激素对基因表达调节的研究(图2)。
2.1 逆境对GA生物合成的调节
非生物胁迫可改变植物GA的代谢途径。DDF1基因的超量表达导致拟南芥活性GA4含量减少和植株矮化,由DDF1基因编码一个AP2与逆境应答紧密相关的干旱应答元件结合转录因子(DREBs)和DDF1在高盐胁迫条件下表达强烈。在高盐条件下,转基因植物DDF1的超量表达和GA缺陷型ga1-3突变体的存活率较高,而施用活性GA处理的存活率降低[18]。通过检测盐胁迫GA相关突变体发现,DELLA蛋白在GA缺陷型ga1突变体中积累,在GA不敏感型gai-1突变体中趋于稳定水平。但在低盐胁迫诱导生长条件下DELLA功能缺失型突变体的存活率降低,不能正常响应对盐胁迫诱导生长的抑制。
张新蕊[22]在生长素和赤霉素参与调节低磷胁迫下玉米根系形态改变的研究中发现,低磷胁迫下,赤霉素受体的基因GID1a和GIDlb表达显著上调,说明赤霉素信号转导途径处于较活跃状态。对玉米进行GA3处理或低磷和GA3处理结果表明,在赤霉素合成过程中抑制控制早期步骤基因的表达,导致玉米根系GA20ox1和GA20ox的表达丰度显著下降,却促进GA2ox1和GA2ox2的表达。其分析GA3处理对GA20ox1、GA20ox2、GA2ox1和GA2ox2表达的影响,得出GA3处理拮抗低磷胁迫产生的效应。
2.2 光对GA生物合成的调节
光调节控制植物一生的生长和发育过程,以适应外界环境的变化。自然环境下,植物需要经过一定时间适宜的光周期诱导才能开花。经过诱导后的植物,内源性GA含量升高。光周期通过影响GA
水平和GA代谢调节植物的生长发育[1]。隐花色素能通过介导蓝光调节GA代谢和代谢基因,影响GA水平和抑制下胚轴伸长,控制部分去黄化表型。拟南芥野生型对照或生长在黑暗或连续的蓝光下的
cry1cry2(cryptochrome1/cryptochrome2)突变体幼苗,其体内的GA4含量没有变化,这表明隐花色素也可以调节GA响应性和触发细胞或者特定组织生物活性GA水平的变化[23]。

图1 高等植物赤霉素生物合成途径[18]
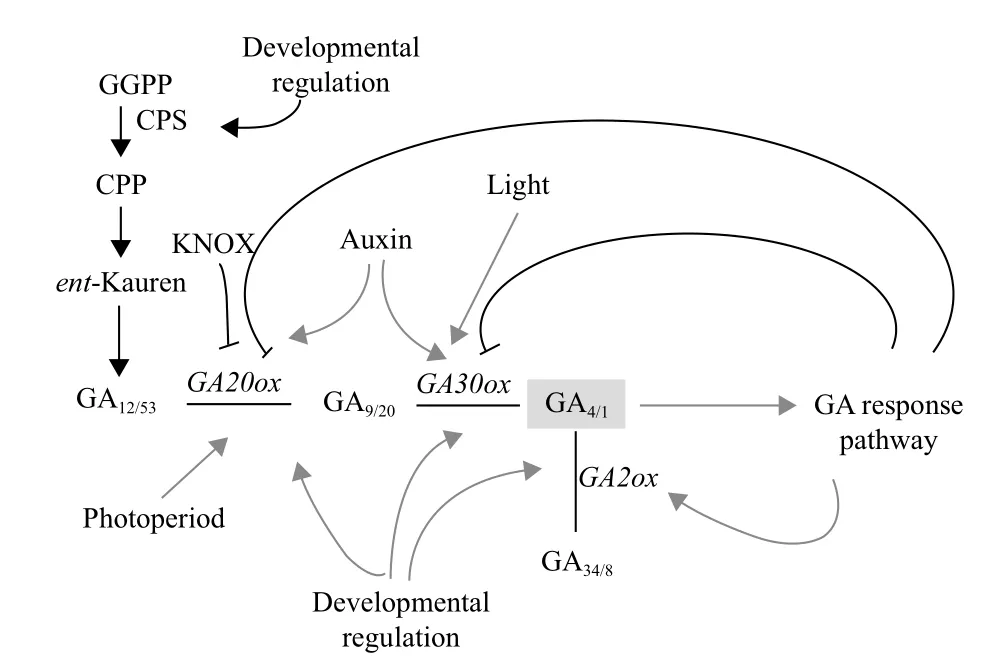
图2 赤霉素合成和代谢酶基因的调控[21]
莴苣和拟南芥种子萌发取决于红光照射,从而提高GA3ox的表达,导致活性GA含量增加。豌豆幼苗的去黄化改变GA20ox和GA3ox基因的表达,诱导GA1含量急速减少。绿色植物茎的伸长也受到不同的光辐照特性的影响。低辐光照能促进豌豆和油菜茎的伸长和活性GA含量的增加。光周期通过长日植物(如天仙子、金光菊和黑眼菊)和木本植物(如杨柳和杂交白杨)GA生物合成的不同阶段调节活性GA的含量,主要是GA20ox和GA3ox基因的转录水平。光通过光敏色素改变GA生物合成和代谢及GA响应,调节需光植物茎的伸长[1,24]。
GA缺乏时,位于核内的DELLA蛋白累积在较高水平,并与光敏色素作用因子PIF3相互作用,阻止PIF3与靶基因启动子结合和调节基因表达,从而消除PIF3介导光调节下胚轴伸长。GA充足时,GA受体GID1蛋白与核内DELLA蛋白相互作用得到提高,引发DELLA蛋白的泛素化和酶体介导降解,从而释放出PIF3[25]。
2.3 温度对GA生物合成的调节
植物种子的春化作用可调节内源GA的水平。经过低温处理,拟南芥种子GA生物合成基因,如AtGA3ox1和AtGA20ox2,能够通过适当地调节表达,导致体内GA水平升高,打破休眠,促进种子萌发。李波[26]在杜鹃花芽形态分化初期低温GA对花期的影响结果显示,随着低温处理时间的延长,其内源激素GA变化呈现先上升后下降的趋势。在拟南芥中,低温处理可提高AtGA20ox1和AtGA20ox2的表达,进而提高活性GA的水平。通过GC-MS分析表明,种子在4℃下的活性GA水平明显高于22℃下的活性[1]。
2.4 生长阶段对GA生物合成的调节
GA生物合成途径早期是由单拷贝或小基因家族调控的。古巴焦磷酸合成酶(CPS)、内根-贝壳杉烯合成酶(KS)和氧化酶(KO)以及内根-贝壳杉烯酸氧化酶(KAO)主要促进GA早期的生物合成,多个CytP450单加氧酶催化内根-贝壳杉烯转化为GA12。
CPS是GA生物合成途径早期的主要限速酶之一。Silverstone等[27,28]研究发现在拟南芥各器官AtCPS转录水平极低,而当AtCPS过表达时,内源性内根-贝壳杉烯水平大增,这说明CPS可能限制拟南芥内根-贝壳杉烯的合成。在向日葵生长和发育阶段GA生物合成早期,HaCPS1L、HaKS2L、HaKS3L基因受到高度调控表达。并且这3个基因的mRNA水平在快速生长的组织中极为丰富。HaKS2L的表达水平比HaCPS1L和HaKS3L较低。在种子发育过程中,胚快速生长期,HaCPS1L和HaKS3L基因的转录水平较高。而且这3个基因不受赤霉素活性的反馈调节[29]。HaKAO1在向日葵各个器官都有表达,而HaKAO2主要在根部表达[30]。
2.5 活性GA水平对GA生物合成的调节
通过外施活性GA可对植物内源活性GA起到调节作用。杨艳华等[31]对几个水稻品种施用活性GA3的研究表明,GA3能够促进水稻地上部分伸长生长,抑制根伸长生长;GA3诱导GA20ox2和GA3ox2基因表达,使内源性GA含量增加。吴建明等[15]采用实时荧光定量PCR研究施用活性GA诱导甘蔗GA20ox的表达情况,结果表明未经过GA处理的甘蔗GA20ox表达量最高,GA处理后GA20ox表达量持续下降,在6 h达到最低值,12 h后有所回升,但低于未处理的,到48 h又呈现明显下降的情况。这说明外施高浓度的活性GA抑制植物内源活性GA生物合成,导致内源活性GA的含量下降。
2.6 其他激素对GA生物合成的调节
植物生长激素(如IAA)至少在一定程度上通过上调GA生物合成基因(如GA3ox和GA20ox)和下调GA分解代谢基因GA2ox的表达,导致生物活性GA1的水平升高,从而影响GA的新陈代谢,调节植物的生长[32]。
Wolbang和Ross等[33,34]采用14C同位素示踪法在去除烟草顶芽IAA促进GA生物合成的研究发现,在去除烟草顶芽切口以下的节间组织中,内源性IAA、GA20和GA1的水平下降,GA生物合成途径:由GA19转变为GA20,再转变为活性GA1的途径降低,并出现由GA20转变为GA29,再转变为GA29代谢物的失活途径;施用IAA后,GA1和GA20的含量增加,其GA20的水平是完整植株的3倍,但GA生物合成途径由GA20转变为GA1不能恢复到完整植株的水平。这表明IAA能够促进GA的生物合成,具有协同作用。
GA与ABA、GA与PP333通常表现为拮抗作用。
GA能够打破种子休眠,促进种子萌发,主要是GA刺激诱导糊粉层α-淀粉酶合成引起。而ABA能通过诱导生成一种脱落酸诱导蛋白激酶(PKABA)阻断GA信号传递,抑制转录因子GAMyb的形成,从而抑制GA的生物合成和种子萌发,促进种子休眠[35]。杨艳华等[31]对几个水稻品种施用10 mg/L ABA的研究表明,ABA处理严重抑制种子萌发和地上部伸长生长;ABA抑制GA20ox2和GA3ox2基因表达,使植物减少体内活性GA的含量。
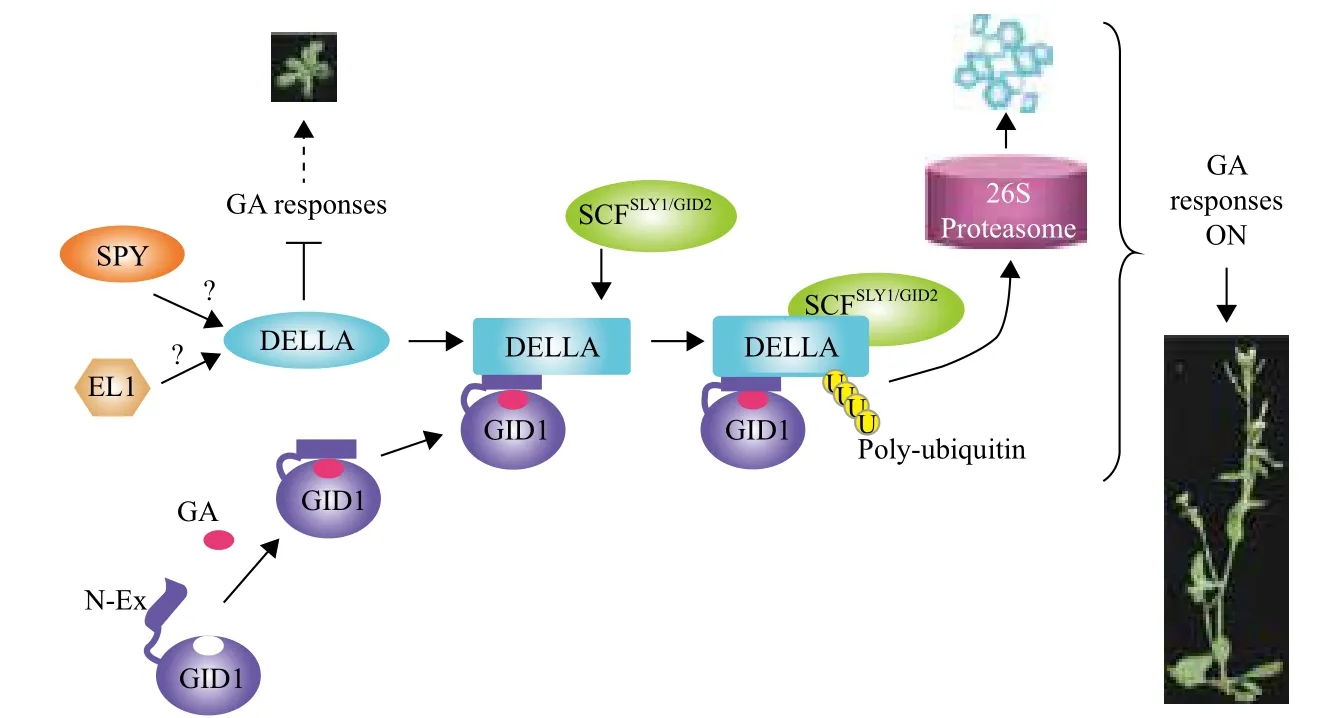
图3 GA信号转导模式[39]
刘芳等[36]在采用外源GA3和PP333对甜樱桃新梢生长及赤霉素代谢关键基因表达的影响试验结果表明,GA3可明显促进新梢生长,GA3处理14 d后可使其内源GA3含量高于对照的1.2倍,随后迅速下降,抑制GA20ox和GA3ox基因的转录表达;而PP333能显著抑制新梢的生长,PP333处理后内源GA3含量呈持续下降趋势,提高GA20ox和GA3ox基因的转录水平。PP333对水稻叶片衰老过程中游离GA4含量的影响中发现,PP333处理并不改变叶片内源游离GA4含量变化的进程,但降低了游离GA4的含量水平。这说明PP333对叶片内GA4的生成有抑制作用[37]。
3 赤霉素信号转导模式
GA信号转导通路中的组分主要包括受体、起到关键调控作用的DELLA蛋白以及介导DELLA蛋白降解的其他调控因子。GA信号转导途径已经基本被确定,根据这一模型(图3)可知,当GA水平较低时,受体GID1不和GA结合,阻遏蛋白DELLA处于高水平,并与靶基因结合,抑制靶基因的转录,进而抑制植物的生长。翻译后修饰,如糖基化和磷酸化也可能影响DELLA的活性。SPINDLY(SPY)是一个含有34个氨基酸重复序列的氧连N-乙酰氨葡萄糖[O-linked GlcNAc(O-GlcNAc)]转移酶(OGT),主要起阻遏GA信号作用。在动物体内,O-GlcNAc定位于核和细胞质内,SPY可通过O-GluNAc修饰激活DELLA,而DELLA的磷酸化是以SPY同靶蛋白质的Ser/Thr位点的O-磷酸化竞争,去除O-磷酸
化而实现O-GluNAc修饰;SPY也能将GluNAc(N-乙酞氨基葡萄糖)转移到含有Ser/Thr的靶蛋白上并对Ser/Thr基团进行糖基化而起到修饰作用。而水稻FLOWERING1(EL1)基因编码的酪蛋白激酶,可使DELLA磷酸化活性保持稳定,因此它也具有阻遏GA信号的功能[7,38-41]。
GA受体GID1的N端具有一个N-Ex的灵活结构,当GA处于较高水平时,GID1可感知GA信号并与之结合,引发GID1的C端结构域的构象改变,从而使N端的N-Ex结构将GA封在GID1的结合位点上,形成GA-GID1复合体。进一步促进了与DELLA相互作用结合,形成较为稳定的GA-GID1-DELLA复合体。在由Skp1、cullin、F-box蛋白质和RING-H2域组成的E3连接酶复合物—SCF复合物的参与下,通过在靶蛋白上添加一个多聚泛素链,由此诱发通过26S蛋白酶复合体途径降解DELLA,从而激活GA活性[39,42]。
4 结语
随着生物技术的高速发展,人们对GA生物合成与代谢,以及信号转导途径有了更深入的了解,阐明了GA合成过程中一些关键基因的调节机制,信号转导途径中相关组分(GA、GID1和DELLA等)间的相互作用。但DELLA蛋白功能的分子机制方面还有许多问题有待解决。而在植物生长发育各个阶段,GA与其他植物激素的水平是如何维持相对平衡的;在GA信号转导途径中,GA受体GID1的结构与功能存在什么样的联系;它们是如何进行识别与结合的;DELLA蛋白通过泛素化降解后,其代谢物的去向如何;这些问题都需要进一步深入地研究才能够解答。SPY和EL1可能是GA信号传导途径中的反向作用因子,但SPY和EL1与DELLA蛋白的相互作用以及修饰反应的遗传调控、分子生物学机制也有待进一步研究[8]。随着GA合成与信号转导分子机制的深入研究,将有助于GA广泛应用于农业生产,加快种质创新与良种繁育。
[1]潘瑞炽. 植物生理学[M].第6版. 北京:高等教育出版社, 2008:167-280.
[2]左圆圆, 周杨杨, 李小平, 等. 植物激素-赤霉素(GA)细胞信号转导机制[J]. 淮北师范大学学报:自然科学版, 2011, 32(2):49-54.
[3]王彦波, 鲜开梅, 张永华, 等. 赤霉素的应用研究进展[J].北方园艺, 2007(6):74-75.
[4]李保珠, 赵翔, 安国勇, 等. 赤霉素的研究进展[J].中国农学通报, 2011, 27(1):1-5.
[5]王洪梅, 周显昌, 周志军, 等. 赤霉素促进针叶树开花结实技术的研究进展[J].林业科技, 2011, 36(3):11-15.
[6]Eriksson S, Bohlenius H, Moritz T, et al. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation[J]. Plant Cell, 2006, 18(9):2172-2181.
[7]黄先忠, 蒋才富, 廖立力, 等. 赤霉素作用机理的分子基础与调控模式研究进展[J].植物学通报, 2006, 23(5):499-510.
[8]谈心, 马欣荣. 赤霉素生物合成途径及其相关研究进展[J].应用与环境生物学报, 2008, 14(4):571-577.
[9]Rieu I, Ruiz-Rivero O, Fernandez-Garcia N, et al. The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle[J]. The Plant Journal, 2008, 53(3):488-504.
[10]Grennan AK. Gibberellin metabolism enzymes in rice[J]. Plant Physiology, 2006, 141(2):524-526.
[11]Lange T, Kappler J, Fischer A, et al. Gibberellin biosynthesis in developing pumpkin seedlings[J]. Plant Physiology, 2005, 139(1):213-223.
[12]Appleford NEJ, Evans DJ, Lenton JR, et al. Function and transcript analysis of gibberellin-biosynthetic enzymes in wheat[J]. Planta, 2006, 223(3):568-582.
[13]Bomke I, Rojas MC, Gong F, et al. Isolation and Characterization of the gibberellin biosynthetic gene cluster in Sphaceloma manihoticola[J]. Applied and Environmental Microbiology, 2008, 74(17):5325-5339.
[14]吴建明, 李杨瑞, 王爱勤, 等. 甘蔗GA20氧化酶基因片段克隆及序列分析[J].热带作物学报, 2009, 30(6):817-821.
[15]吴建明, 李杨瑞, 王爱勤, 等. 赤霉素诱导甘蔗GA20-Oxidase基因实时荧光定量PCR分析[J].分子植物育种, 2009, 7(5):922-927.
[16]岳川, 曾建明, 曹红利, 等. 高等植物赤霉素代谢及其信号转导通路[J].植物生理学报, 2012, 48(2):118-128.
[17]吴建明, 李杨瑞, 董伟清, 等. 赤霉素的生物合成及其在甘蔗上的应用研究[J]. 作物杂志, 2007, (6):23-30.
[18]Yamaguchi S. Gibberellin metabolism and its regulation[J]. Annual Review of Plant Biology, 2008, 59:225-251.
[19]张迎迎, 何祖华. 高等植物赤霉素的代谢与信号转导[J].植物生理学通讯, 2010, 46(7):623-630.
[20]Middleton AM, Úbeda-Tomás S, Griffiths J, et al. Mathematical modeling elucidates the role of transcriptional feedback in gibberellin signaling[J]. Proc Natl Acacl Sci USA, 2012, 109(19):7571-7576.
[21]Hedden P, Phillips AL. Gibberellin metabolism:new insights revealed by the genes[J]. Trends in Plant Science, 2000, 5(12):523-530.
[22]张新蕊.生长素和赤霉素参与调节低磷胁迫下玉米根系形态的改变[D]. 济南:山东大学, 2011.
[23]Zhao XY, Yu XH, Foo E, et al. A study of gibberellin homeostasis and cryptochrome-mediated blue light inhibition of hypocotyl elongation[J]. Plant Physiology, 2007, 145(1):106-118.
[24]García-Martinez JL, Gil J. Light regulation of gibberellin biosynthesis and mode of action[J]. Journal of Plant Growth Regulation, 2001, 20(4):354-368.
[25]Feng SH, Martinez C, Gusmaroli G, et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins[J]. Nature, 2008, 451(7177):475-479.
[26]李波. 生长调节剂与光温处理对西洋杜鹃花期及内源激素的调控作用[D].杭州:浙江大学, 2012.
[27]Silverstone AL, Chang CW, Krol E, et al. Developmental regulation of the gibberellin biosynthetic genes GA1in Arabidopsis thaliana[J]. Plant J, 1997, 12(1):9-19.
[28]Yamaguchi S. Gibberellin biosynthesis in Arabidopsis[J]. Phytochemistry Reviews, 2006, 5(1):39-47.
[29]Pugliesi C, Fambrini M, Salvini M. Molecular cloning and expression profile analysis of three sunflower(Helianthus annuus)diterpene synthase genes[J]. Biochem Genet, 2011, 49(1-2):46-62.
[30]Fambrini M, Mariotti L, Parlanti S, et al. The extreme dwarf phenotype of the GA-sensitive mutant of sunflower, dwarf2, is generated by a deletion in the ent-kaurenoic acid oxidase1(HaKAO1)gene sequence[J]. Plant Mol Biol, 2011, 75(4-5):431-450.
[31]杨艳华, 张亚东, 朱镇, 等. 赤霉素(GA3)和脱落酸(ABA)对不同水稻品种生长和生理特性及GA20ox2、GA3ox2基因表达的影响[J].中国水稻科学, 2010, 24(4):433-437.
[32]Reid JB, Davidson SE, Ross JJ. Auxin acts independently of DELLA proteins in regulating gibberellin levels[J]. Landes Bioscience Plant Signaling & Behavior, 2011, 6(3):406-408.
[33]Wolbang CM, Ross JJ. Auxin promotes gibberellin biosynthesis in decapitated tobacco plants[J]. Planta, 2001, 214(1):153-157.
[34]Ross JJ, O’Neill DP, Wolbang CM, et al. Auxin-gibberellin interactions and their role in plant growth[J]. Journal of Plant Growth Regulation, 2002, 20(4):346-353.
[35]石海燕, 郭靖, 周颖, 等. 赤霉素和脱落酸在植物生长发育中相互关系的研究进展[J].华中师范大学研究生学报, 2007, 14(1):138-142.
[36]刘芳, 袁华招, 沈欣杰, 等. 外源GA3和PP333对甜樱桃新梢生长及赤霉素代谢关键基因表达的影响[J].核农学报, 2013, 27(3):272-278.
[37]俞炳呆, 严景华. PP333对水稻叶片衰老过程中内源GA4和ABA含量的调节作用[J].南京农业大学学报, 1995, 18(1):101-103.
[38]肖景华. 番茄GA20-氧化酶和GA2-氧化酶基因的克隆与功能分析[D].武汉:华中农业大学, 2006.
[39]Sun TP. Gibberellin-GID1-DELLA:a pivotal regulatory module for plant growth and development[J]. Plant Physiology, 2010, 154(2):567-570.
[40]王荣, 张根发.赤霉素信号转导的中间组分研究[J].生物学杂志, 2007, 24(2):5-8.
[41]Wang YJ, Deng DX. Molecular basis and evolutionary pattern of GA-GID1-DELLA regulatory module[J]. Mol Genet Genomics, 2014, 289(1):1-9.
[42]Hirano K, Ueguchi-Tanaka M, Matsuoka M. GID1-mediated gibberellin signaling in plants[J]. Trends in Plant Science, 2008, 13(4):192-199.
(责任编辑 狄艳红)
Gibberellins Biosynthesis and Signaling Transduction Pathway in Higher Plant
Li Qiang1,2Wu Jianming2Liang He1Huang Xing2Qiu Lihang2
(1. Agricultural College of Guangxi University,Nanning 530004;2. Key Laboratory of Sugarcane Biotechnology and Genetic Improvement(Guangxi),Ministry of Agriculture,P. R. China,Sugarcane Research Center,Chinese Academy of Agricultural Sciences,Nanning 530007)
The hormone gibberellins(GA)plays an essential role in many aspects of plant growth and development, such as promoting seed germination, stem elongation, occurrence of flower, fruit and seed development. This review elaborated the GA biosynthesis and signaling transduction pathway, as well as interactions with other hormones, response to the environment signals, and the recent advances in understanding of degradation of DELLA proteins by polyubiquitination of the DELLAs via the E3 ubiquitin-ligase SCFSLY1/GID2in the 26S proteasome. This helps not only people understanding of gibberellin physiology and its molecular regulating mechanism, but also to every aspect of gibberellin mechanism are studied deeply.
Gibberellins Signal transduction Plant hormones Regulation mechanism
2014-03-17
国家自然科学基金项目(31360312),国家科技支撑计划子课题(2012BAD40B04-3)
李强,男,硕士研究生,研究方向:作物生理与调控;E-mail:274228904@163.com
吴建明,男,博士,副研究员,研究方向:甘蔗栽培及分子生物学;E-mail:wujianming2004@126.com
