短截杧果结果母枝的果糖含量动态变化
2014-02-28汪开华洪明伟李雪娇杨立菊
彭 磊,汪开华,杜 飞,洪明伟,李雪娇,杨立菊,周 玲,王 莹,周 雯
(1.云南农业大学园林园艺学院,云南昆明 650201;2.华坪县农业局,云南华坪674800;3.云南农业大学龙润普洱茶学院,云南昆明 650201)
短截杧果结果母枝的果糖含量动态变化
彭 磊1,汪开华2,杜 飞1,洪明伟1,李雪娇1,杨立菊2,周 玲3,王 莹1,周 雯1
(1.云南农业大学园林园艺学院,云南昆明 650201;2.华坪县农业局,云南华坪674800;3.云南农业大学龙润普洱茶学院,云南昆明 650201)
测定杧果花芽分化期间结果母枝各器官内果糖含量,探索果糖在杧果花芽分化中的作用。利用杧果花枝短截后剪口芽在当年还能再次花芽分化并开花结果的现象,定期测定剪口叶、附近韧皮部及芽内果糖含量。剪口叶内果糖含量于2月20日达到最高(58.66µg·g-1),附近韧皮部及芽于3月12日达到最高,分别为67.60和61.20µg·g-1;叶片于3月2日降至最低(34.93µg·g-1),韧皮部于2月25日降至最低(28.44µg·g-1),剪口芽内果糖含量于短截当日最低(23.96µg·g-1);各检测部位平均含量分别为43.67, 48.76和49.39µg·g-1。剪口叶、附近韧皮部及芽内果糖存在从剪口叶流向附近韧皮部,再到剪口芽的运输通道现象,表明杧果花芽分化期间碳水化合物主要以果糖的形式进行运输,果糖是杧果花芽分化期间碳水化合物的主要运输形式和能量物质。短截后,剪口芽芽眼中心颜色于2月20日出现蜡黄迹象,25日向芽体四周扩散;3月2-7日,芽体逐渐膨大,蜡黄色加深、芽眼呈半透明状;3月7日后绿色逐渐增加,半透明状消失。
杧果;花芽分化;结果母枝;短截;果糖
果树花芽分化研究历来是果树生理学的重要研究内容,也是果树发育生物学的热点之一,因与生产密切相关,所以倍受人们重视[1]。杧果(Mangife raindicaL.)是云南省热区退耕还林中主要的经济林树种之一。由于每年都有倒春寒发生,杧果花穗受害程度随倒春寒发生程度不同有很大差异,发生严重的年份,几乎绝收。在一次生产过程中,课题组偶然发现短截杧果花枝后,剪口下1~3芽没抽梢而开花,倒春寒和花期主要病害白粉病危害高峰过后进入盛花期[2]。此开花特性的发现,为杧果花期回避自然灾害和减少农药用量、促进环境保护提供了一种新的生产方法。为杧果花芽分化过程中各种内含物的种类、含量测定及其功能研究提供了极为便利的方式和研究条件[3-7]。许多报道表明,糖是一个非常关键的因子[8-9]。由于蔗糖是主要的光合产物,是非还原糖,在筛管中占汁液干重的90%,在绿色细胞中含量较高,其理化性质适宜长距离运输[10-11]。故许多学者认为,蔗糖是碳水化合物的主要运输形式[12-14]。陈清西等[15]用K C l 03诱导龙眼树花芽分化,证明龙眼成花的过程中,淀粉含量由高水平逐渐下降、可溶性总糖和果糖含量逐渐上升,但杧果花芽分化各阶段中果糖的作用尚不清楚。果树花芽分化具有长期性和相对集中性的特点[16]。研究应用开花的杧果结果母枝短截后,剪口芽还能在当年春季再次进行花芽分化并开花结果的特性,选用云南省主栽品种3年杧为试验材料进行短截处理。采集的样品属3年杧花芽分化进程中的何时期,确定果糖含量与花芽分化某阶段的对应关系。
1 材料与方法
1.1 材料
试验在元江农场(海拔585 m)进行。选用14年生、长势及树体营养基本一致的品种3年杧(M.i ndi c a L.)植株为试验材料。
1.2 方法
2010年2月15日,对正在开花的结果母枝进行短截,单株为1次重复,重复5次。以后每5 d随机选取植株上、中、下部剪口芽、附近韧皮部、叶片进行采样,直至花芽开始膨大、准备萌发时止。设不短截植株为对照(采样时才进行短截,取剪口芽、附近韧皮部、叶片)。各重复中采过样的枝用红油漆进行标记,避免重复采样。采下的材料放入自封袋,封好后置入冰壶带回实验室检测。
用蒽酮比色法[17]测定果糖含量。
1.3 数据处理
用Excel对原始数据进行初步整理,用DPS(D unc a n法)进行差异显著性分析。
2 结果与分析
2.1 剪口芽花芽分化期间外观性状变化
从短截至花芽开始膨大,剪口芽外观性状有较明显变化。2月15日短截时,剪口芽镶嵌于叶腋内呈扁平状,芽眼呈绿色;芽体最外层垂直方向的左右两侧各有一鳞片包被,两鳞片非闭合,类似半闭的眼睛。2月20日芽眼出现蜡黄迹象。2月25日,芽体稍微向上凸起,此时可见最外层鳞片下,芽体水平方向的上下两侧也各有一鳞片包被,芽眼中心颜色趋向于蜡黄。3月2日至7日,芽体逐渐膨大,且呈蜡黄、半透明状,透明性较3月2日有较大增加,3月7日后此现象逐渐消失而绿色增加,至12日花芽进入松散及花穗伸长期。
2.2 剪口叶果糖含量
图1显示,短截后5 d内,剪口叶内果糖含量升至试验期内最高值,从38.87升至58.66µg·g-1,当芽眼出现蜡黄迹象时,呈缓慢下降趋势,2月25日后大幅降至试验期内最低值;芽体逐渐膨大且呈蜡黄、半透明状后(3月2日),叶内果糖含量呈水平趋势。5 d内对照剪口叶内果糖含量急剧升至试验期内最高值,从35.93升至67.92µg·g-1,随后大幅降至期内最低值,3月2日后缓升直至12日。多重比较表明,短截处理与C K的各检测部位果糖含量差异均不显著。
2.3 剪口附近韧皮部果糖含量
图2显示,短截后韧皮部内果糖含量波动较大,2月20日升至49.96µg·g-1,芽眼出现蜡黄迹象时,于25日又降至期内最低值(28.44µg·g-1),后持续上升,于3月12日达期内最高值(67.60µg·g-1)。C K也于短截后5 d内大幅上升,并达期内最高值(53.72µg·g-1),之后持续下降,3月7日后呈回升趋势。多重比较表明,与C K相比,短截处理韧皮部内果糖含量于3月7日在1%水平上差异极显著,2月25日和3月12日,在5%水平上差异显著。
2.4 剪口芽果糖含量
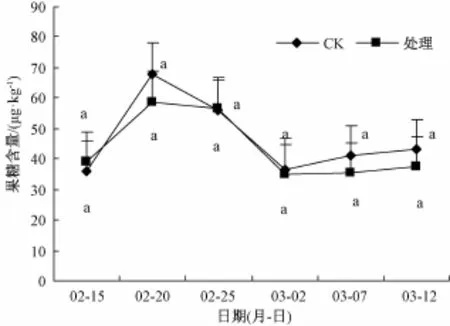
图1 叶片果糖含量变化
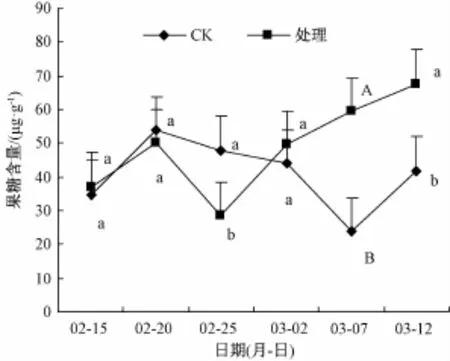
图2 韧皮部果糖含量

图3 剪口芽果糖含量
图3 显示,短截处理后至3月2日,当芽体逐渐膨大且呈蜡黄、半透明状时,剪口芽内果糖含量呈现持续上升趋势,从23.96升至61.12µg·g-1。
至3月7日略微下降,2月12日达期内最高值(61.20µg·g-1)。从2月15日到25日,C K芽体稍微向上凸起,芽眼中心颜色趋向于蜡黄时,芽内果糖含量从31.08升至70.20µg·g-1。3月2日又大幅降至41.52µg·g-1,后持续升至期内最高值(75.24µg·g-1)。多重比较结果表明,与C K相比,短截处理剪口芽内果糖含量于2月25日,3月2日和12日,在1%水平上差异达极显著。
3 小结与讨论
李桂芬[18]对紫花杧结果母枝顶芽切片观察表明,花芽未分化时芽体瘦小,外有褐色苞片包裹,当顶芽的基部膨大,苞片顶尖稍微松开,已进入花芽分化前期,之后进入花序分化期,外观上看顶芽的基部继续膨大,苞片顶尖松开,出现花序原始体突起。此试验中,3月2日芽体开始膨大,表明此前属花芽分化前期;短截后第15日,剪口芽开始进入花芽分化期。
2月20日,短截处理的叶内果糖含量达最高值,之后持续下降,到花芽分化结束时含量一直处于较低水平;2月20-25日,剪口附近韧皮部内果糖含量大幅下降,可能向芽输送而引起含量下降,随后由于叶内果糖向芽输送途经韧皮部,于2月25日后其含量持续上升;剪口芽内果糖含量则一直呈持续上升趋势,花芽分化结束时达试验期内最高值。因此,花芽分化临界期前后,剪口芽附近的组织可能向此时的生长中心(剪口芽)输送果糖。对照的叶于2月20至3月2日、韧皮部2月20日至3月7日呈下降趋势,芽内果糖含量2月25至3月2日也呈大幅下降趋势,表明对照的叶、韧皮部和芽内果糖主要输向其他组织或器官。
试验结果显示,短截杧果结果母枝后,存在果糖从剪口叶流向附近韧皮部,再到剪口芽的运输通道现象,表明此期杧果的碳水化合物主要以果糖的形式进行运输,并且是花芽分化的主要能量物质。由此推测是否存在每年仅开1次花的果树,其碳水化合物主要以蔗糖的形式进行运输,而具有1年多次开花潜能的果树,其碳水化合物主要以果糖形式进行运输,或花芽分化期间转为以果糖形式进行运输,或近距离运输以果糖形式运输为主。然而陈清西等[15]研究虽然证明龙眼成花过程中果糖含量逐渐上升,但至今尚无龙眼在一年内进行二次开花的报道,因此,具有二次开花潜能的果树碳水化合物主要以果糖的形式进行运输的假设还需进一步探索。
虽然短截杧果结果母枝后没对叶、附近韧皮部内的果糖进行标记,但根据检测结果可以推定花芽分化临界期前后,剪口芽附近的组织向剪口芽输送果糖,以满足花芽分化所需的营养和能量物质。剪口芽花芽分化前期和分化期,致使芽眼呈现蜡黄、半透明状现象的因素或机制尚不清楚。
[1] 陈杰忠,赵红业.芒果花芽分化研究进展[J].中国南方果树,1999,28(2):34-35.
[2] 彭磊,唐发贵,王信保,等.杧果花枝回缩后剪口芽再花研究[J].中国南方果树,2006,35(3):35-36.
[3] 彭磊,高小俊,龙雯虹,等.短截后芒果花芽分化期间A B A含量的变化[J].云南农业大学学报:自然科学版, 2011,26(3):434-436.
[4] 彭磊,周玲,王莹,等.杧果短截结果母枝玉米素核苷含量的动态变化[J].南方农业学报,2013,44(1):39-42.
[5] 彭磊,周玲,汪开华,等.杧果短截结果母枝蔗糖含量动态变化[J].中国农学通报,2013,29(7):117-120.
[6] 彭磊,周玲,王莹,等.杧果短截结果母枝葡萄糖含量动态变化[J].江西农业学报,2012,24(11):5-7.
[7] 彭磊,周玲,高小俊,等.杧果短截结果母枝G A3含量动态变化[J].农业研究与应用,2012(6):5-8.
[8] 王广鹏,孔德军,刘庆香.营养成分调控果树花芽分化研究进展[J].云南农业大学学报,2009,24(6):908-912.
[9] 李天红,黄卫东,孟昭清.苹果花芽孕育机理的探讨[J].植物生理学报,1996,22(3):251-257.
[10] 白宝璋,叶尚红,王玉国.植物生理学[M].北京:中国农业科技出版社,1996:134-136.
[11] Budan T, FaustM.Flowerbud initiation in appletrees:internalcontroland differentiation [J].HorticulturalReviews,1982, 4: 174 -203.
[12] apex of straw berryunder flowerbud inductive conditions[J].Acta Horticulturae, 2003, 626: 305 -308.
[13]apex of strawberr yunder flowerbud inductiveconditions[J].ActaHorticulturae, 2003, 626: 305 -308.
[14]carbohydrate contents in shoottips, leaves and roots ofstrawberry( Fragaria xAnanassa Duch.) duringflower-buddifferentiation [J].ScientiaHorticultural, 2007, 113 ( 3 ) :255 -260.
[15] 陈清西,李松刚.K C l 03诱导龙眼成花及其叶片碳水化合物与蛋白质的变化[J].福建农林大学学报:自然科学版,2004,33(2):182-185.
[16] 河北农业大学.果树栽培学总论[M].2版.北京:中国农业出版社,1980:62-63.
[17] 中国科学院上海植物生理研究所.现代植物生理学试验指导[M].北京:科学出版社,1999.
[18] 李桂芬.芒果花期调控及花芽分化的研究[D].南宁:广西大学,2005.
(责任编辑:张瑞麟)
S 667.7文献标志码:A文章编号:0528-9017(2014)05-0689-03
2013-12-18
云南省应用基础研究计划项目(2007C 0058M);云南省教育厅科学研究基金(2010Y 338)
彭 磊(1969-),云南元江人,副教授,硕士研究生,主要从事热带果树研究工作。E-mail:ppl l 250@y na u.e du.c n。
文献著录格式:彭磊,汪开华,杜飞,等.短截杧果结果母枝的果糖含量动态变化[J].浙江农业科学,2014(5):689-691.
