不同生境中橘小实蝇种群动态及密度的差异
2013-12-20郑思宁
郑思宁
(1.浙江大学中国农村发展研究院,杭州 310058;
2.福建农林大学益虫研究所 联合国(中国)实蝇防控中心,福州 350028)
橘小实蝇Bactrocera dorsalis(Hendel),俗称果蛆、金苍蝇,又称东方果实蝇,属双翅目Diptera、实蝇科Tephritidae、寡毛实蝇亚科Dacinaet昆虫,是一种世界性检疫害虫。现已证明了橘小实蝇是由52个姐妹种组成的复合种。作为一种外来入侵害虫,橘小实蝇寄主范围广、危害大、防治困难[1-2],在国内主要分布于长江以南地区,严重为害多种水果和蔬菜[3-4]。
关于橘小实蝇的种群动态,目前国内开展了多方面的研究,但研究主要集中在单一寄主植物该虫的消长动态及气象因素对其种群动态的影响[5-13],缺乏对不同生境条件橘小实蝇种群动态的系统研究。本文通过大量的田间调查试验力求全面了解多生境状态下该虫的种群消长动态,为该虫的田间防治提供参考。
1 材料与方法
1.1 试验材料
甲基丁香酚(性诱剂,methyl eugenol,ME):江苏禾丰生化研究所生产,为纯化学品,含量≥99%。
诱捕器:佛山顺德容桂利合塑料五金厂生产(专利号:03361995.6),诱芯材质为木纤维,诱芯载量约为2g ME。
1.2 试验方法
试验于2008年5月—2009年1月在福州各监测点进行。试验方法为:每7d收集清点诱捕器内的橘小实蝇1次,并清理瓶内虫子。夏季每7d在诱芯上添加1次ME,冬季每14d加1次ME。诱集瓶长期挂在野外,为避免其因风吹日晒颜色变化而影响监测结果,每半年更换1次诱捕器。
1.2.1 不同寄主果园的橘小实蝇种群动态
监测点概况:监测点分别选择在福州市闽侯县武警副食品基地、消防总队副食品基地,闽侯县闽侯龙台山生态园。
闽侯武警副食品基地果园位于上街,园内地势平缓,果园混合种植有枇杷、芭乐、四季杨桃及芒果等果树品种,面积约2.5hm2;消防总队副食品基地与武警副食品基地相隔约1km,主要以葡萄、丝瓜和黄瓜等藤本植物为主,面积约3.4hm2;闽侯龙台山生态园与闽侯荆溪铁岭工业区相邻,种植的果树品种以柑橘类为主,兼种少量芭乐、杨梅等,面积近70hm2。每个果园设置3个诱捕器,每个诱捕器之间的距离约为50m。
1.2.2 果园与果园周边丛林橘小实蝇的种群动态
试验在闽侯武警副食品基地果园内及果园周边进行,诱捕器每隔约50m设置1个,共设6个监测诱捕器,其中果园内3个,果园南侧的小山丘上挂3个诱捕器。
1.2.3 果园内不同地理环境的橘小实蝇的种群动态
山谷生境与山脊生境的橘小实蝇种群动态试验在闽侯龙台山生态园进行(该监测点寄主植物为柑橘类植物),共设6个监测诱捕器,其中果园内3个。果园南侧的山脊上挂3个诱捕器,两侧的诱捕器每隔约50m设置1个。
沿河生境与非沿河生境橘小实蝇种群动态试验在福州市的白马河公园内进行(该监测点的寄主植物为芒果)。每隔约50m设置1个诱捕器,其中沿河阴湿处悬挂5个诱捕器,离河150m处设5个。
1.2.4 城区与郊区果园的橘小实蝇的种群动态
试验在福州市区、福州市与闽侯交接地区和闽侯地区的多个监测点进行(各监测点的寄主植物均以芒果为主)。第1个监测点位于福州市中心的乌山景区公园内,设5个诱捕器;第2个监测点位于福州市白马河公园内,共设5个诱捕器;第3个监测点位于福州市与闽侯交界的闽都大庄园内,共设5个诱捕器;第4个监测点位于闽侯马保的武警副食品基地果园内,共设5个诱捕器。其中,前两个监测点代表城区,第3个监测点代表城乡之交,第4个监测点代表乡村区域。
1.3 数据处理
试验数据用SPSS 16.0统计分析软件进行处理。试验2.1.1用两向分组单个观察值资料的方差分析:不同处理看作A因素,不同日期看作B因素(日期间的F测验不必进行,因为试验目的不是研究日期效应)。试验2.1.2、2.2、2.3 和 2.4 用 paired-samples T-test(配对 t测验法)检验不同生境橘小实蝇种群数量间的显著性,用相关性分析考察种群动态之间的关系[14]。
2 结果与分析
2.1 不同果园橘小实蝇的种群动态
2.1.1 单一果园橘小实蝇的种群动态
各单一果园橘小实蝇种群消长动态见图1所示。
2008年5月到2008年12月,芭乐园和杨桃园橘小实蝇种群动态监测试验在闽侯武警副食品基地进行。试验结果表明:芭乐园橘小实蝇的种群数量在7、8月份开始缓慢上升,9月初到10月中旬其种群数量迅速上升,最高诱捕数为9月23日—9月30日的968.33头/瓶。10月下旬随着芭乐成熟期的结束,种群数量逐渐下降。可见,9月至10月是芭乐园里该虫的主要发生时期。
杨桃园橘小实蝇的种群数量从8月份开始缓慢上升,9月上旬迅速上升,峰值的诱捕数分别为9月9日—9月16日的950.20头/瓶和10月4日—10月11日的661.67头/瓶。11月份,杨桃成熟期过后,其种群数量开始逐渐下降。可见,9至10月是杨桃里该虫发生的主要时期。
葡萄属多年生藤本植物,也是橘小实蝇的嗜好寄主植物之一。从图1可看出,葡萄园的橘小实蝇种群数量在7月上旬开始缓慢上升,8月中旬橘小实蝇的种群数量迅速上升,峰值的诱捕数为9月23日—9月30日的780.33头/瓶。11月后种群数量开始逐渐下降。说明8、9、10月这3个月是葡萄园橘小实蝇发生的主要时期。
丝瓜和黄瓜等蔬菜也是橘小实蝇的寄主植物之一。由图可知,丝瓜园橘小实蝇的种群数量在7月下旬开始缓慢上升,8月份迅速上升,8月底到9月出现第1个高峰期,其峰值分别为9月9日—9月16日和9月23日—9月30日的164.33头/瓶和192.67头/瓶。第2个高峰期出现在10月下旬,峰值在10月28日的162.33头/瓶。11月份后其种群数量开始逐渐下降。可见,8、9、10月是丝瓜园里该虫发生的主要时期。黄瓜园橘小实蝇的种群数量在8月中旬开始缓慢上升,9月中旬其种群数量迅速上升,最高诱捕数为9月23日—9月30日的304.67头/瓶。到10月中下旬又呈现第2个峰值,最高诱捕数出现在10月14日—10月21日的301.00头/瓶。11月份,随着黄瓜采收期的结束,种群数量逐渐下降。可知,9至10月是黄瓜园里该虫发生的主要时期。
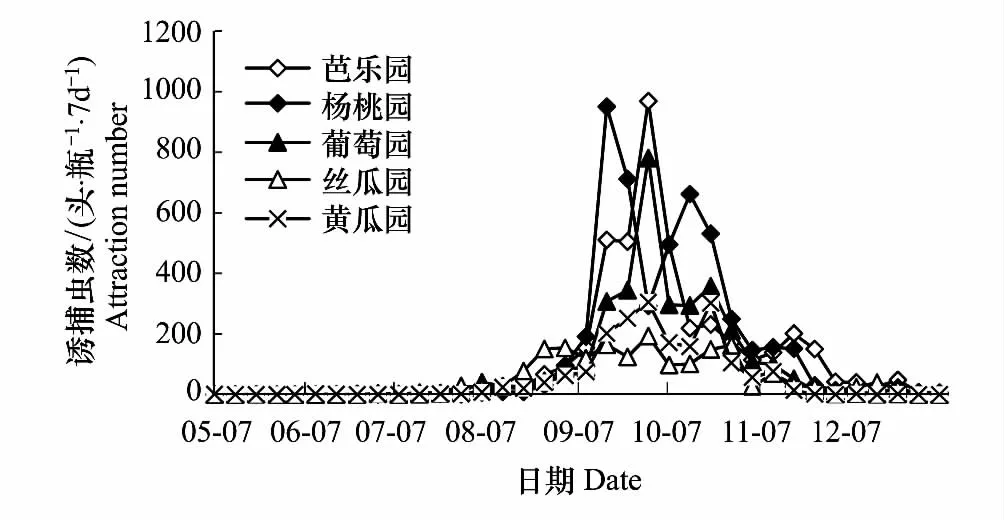
图1 不同果园橘小实蝇种群动态Fig.1 The population dynamics of different orchards
从5个果园橘小实蝇种群密度的显著性比较看(表1),芭乐园橘小实蝇种群密度与葡萄园、丝瓜园和黄瓜园存在显著差异,与丝瓜园、黄瓜园存在极显著的差异。而杨桃园与丝瓜园、黄瓜园的橘小实蝇种群数量存在极显著差异。可见,芭乐、杨桃等多年生瓜果是橘小实蝇的嗜好寄主,而该虫对丝瓜、黄瓜等葫芦科1年生藤本植物的喜好程度不强,甚至不如葡萄等多年生藤本植物(葡萄园与丝瓜和黄瓜园的种群数量存在5%显著水平的差异)。

表1 不同果园橘小实蝇种群显著性比较*Table 1 Comparison of the population dynamics of B.dorsalis among different orchards
2.1.2 单一果园与混合果园橘小实蝇种群动态比较
单一果园与混合果园橘小实蝇种群动态比较试验在龙台山生态园进行,比较果园类型选择柑橘园、柚子园以及柑橘和柚子的混合果园(图2)。
从图可看出,柑橘园橘小实蝇的种群数量在8月中下旬开始缓慢上升,9月中旬其种群数量迅速上升。之后出现了3个峰值,分别在9月下旬(9月22日—9月29日的142.29头/瓶)、10月下旬(10月20日—10月27日的138.33头/瓶)和11月中旬(11月10日—11月17日的95.20头/瓶)。11月下旬后随着气温下降,橘小实蝇发生量迅速下降。说明9月下旬到11月中旬是该虫发生的主要时期。
柚子园橘小实蝇的种群数量在8月中下旬开始缓慢上升,9月中旬其种群数量迅速上升。高峰期主要集中在10月中上旬到11月中旬。高峰期出现了3个峰值,分别出现在10月中旬(10月6日—10月13日的121.33头/瓶)、10月下旬(10月20日—10月27日的159.20头/瓶)和11月中旬(11月10日—11月17日的197.67头/瓶)。11月下旬后由于柚子的陆续采收以及气温的下降,橘小实蝇发生量开始震荡下降。可见,10月上旬到11月中旬是柚子园该虫的主要发生期。
另外,从监测结果可明显地看出混合果园和单一果园的橘小实蝇的发生规律有着明显的区别:首先,混合果园的总体虫量要远远大于单一果园:混合果园总诱虫量为2511.10头/瓶,单一柑橘园和单一柚子园的总诱虫量分别为1167.67头/瓶和1371.67头/瓶,前者是后者的两倍。paired-samples T-test的结果表明,混合果园与后二者的差异均达到极显著水平(P值均为0.000,小于0.01,差异达到极显著水平)。其次,在种群动态变化方面,混合果园中橘小实蝇种群数量在整个主要发生期波动不大,虫口密度始终维持在高位;而单一果园则波动较大,单一柑橘园和柚子园在监测期内分别呈现3次比较大的波动,出现了3次峰值。这是因为混合果园的各种果成熟期各异,寄主种类较多,有利于种群数量稳定地维持在比较高的水平。

图2 柑橘、柚子及混合果园橘小实蝇种群动态Fig.2 The population dynamics of B.dorsalis in citrus,grapefuit and mixture orchard
2.2 果园与果园周边丛林橘小实蝇的种群动态
图3给出了2008年7月—2009年1月闽侯武警副食品基地果园与周边丛林的橘小实蝇种群消长动态。由图可以直观地看出,8月份开始到10月7日,果园内各种水果成熟转色,田间橘小实蝇种群数量迅速上升,这时果园内部的种群数量大于果园周边的种群数量。10月7日后,随着园内水果成熟期的结束,田间种群数量迅速下降,这时果园周边的种群数量大于果园内。
分别对图3的10月7日前和10月7日后的数据(不包含10月7日的数据)进行t测验和及相关性分析发现:
2008年10月7日前果园内和果园周边丛林2种不同的生境平均每7d诱捕到橘小实蝇数分别为216.95头/瓶和133.02头/瓶。前者是后者的1.3倍,差异达到显著水平(P=0.043<0.05)。此外,两种生境橘小实蝇种群数量增减趋势基本一致,相关系数达0.893,决定系数为0.797。可见,两者关系密切,这是由于橘小实蝇迁飞能力强决定的。
2008年10月7日后果园内和果园周边丛林2种不同的生境平均每7d诱捕到橘小实蝇数分别为127.95头/瓶和304.56头/瓶。后者是前者的2.4倍,差异达到极显著水平(P=0.001<0.01)。该时期两种生境橘小实蝇种群数量增减趋势同样基本一致,相关系数达0.964,决定系数为0.930,二者均包含了对方90%以上的信息,二者关系极为密切。
由以上分析可知,果园内和果园周边丛林橘小实蝇种群数量动态是一致的,但在10月7日前前者大于后者,且差异达到显著。在10月7日后后者大于前者,差异达到极显著。发生这种变化的原因可能与水果采收期结束有关。
2.3 果园内不同地理环境橘小实蝇的种群动态
已有的研究表明,光照和温湿度对橘小实蝇活动有很大的影响[15-18]。为此,本研究通过对同一果园内山谷与山脊、沿河与非沿河生境橘小实蝇种群动态的监测,考察同一果园不同地理环境对橘小实蝇种群动态的影响。
果园内不同地理环境橘小实蝇种群动态监测试验于2008年7月—2009年1月分别在龙台山生态园和白马河果园进行,监测结果见图4。由图可以直观地看出,山谷生境的橘小实蝇种群数量要大于山脊生境,沿河生境的橘小实蝇种群数量要大于非沿河生境沿河生境。

图3 果园与果园周边丛林橘小实蝇种群动态比较Fig.3 Comparison of the population dynamics of B.dorsalis between orchard and its surroundings jungle

图4 果园内不同地理环境橘小实蝇种群动态比较Fig.4 Comparison of the population dynamics of B.dorsalis between different geographical environment in orchard
配对t测验及相关性分析的结果进一步揭示了以上结论:山谷和山脊2种不同的生境平均每7d诱捕到橘小实蝇数分别为31.24头/瓶和53.90头/瓶。后者是前者的1.7倍,差异达到极显著水平(P=0.002<0.01)。且二者的种群数量增减趋势基本一致,相关系数为0.917,决定系数为0.841,二者均包含了对方84%以上的信息。可见,二者关系极为密切。同样,沿河生境与非沿河生境每7d诱捕到橘小实蝇数分别为529.23头/瓶和198.67头/瓶,后者是前者的2.7倍。t测验及相关性分析的结果表明,二者差异达到极显著水平(P=0.000<0.01),二者的增减趋势同样保持基本一致,相关系数达0.926,决定系数为0.858,二者均包含了对方84%以上的信息。
2.4 城区与郊区果园的橘小实蝇的种群动态
几个监测点的2008年5月—2008年12月的橘小实蝇种群动态见图5。
由图可知,城市两个监测点的橘小实蝇种群数量和消长趋势均比较接近:种群数量在6月前很低,一般每个诱捕器7d只诱捕到1—2头,6月其种群数量仍然维持在较低水平,7月初其种群数量迅速上升,高峰期出现在8月中旬到10月底。有2个明显的峰值,分别于9月4日和9月18日,白马河公园和乌山两个监测点在这两个观察日分别诱捕到橘小实蝇2031.15、2329.00头/瓶和2433.70、2292.00 头/瓶。进入10 月份种群数量开始迅速下降,到11月份随着气温的下降种群数量明显下降,进入12月,每7d每诱捕器仅诱捕到3—8头橘小实蝇。市区的两个监测点8个月诱虫总数分别为:16404.50 头/瓶和16381.00 头/瓶。
城市与乡村交界地域橘小实蝇的种群在7月前数量很低,一般每个诱捕器7d只诱捕到1—2头,7月其种群数量仍然很低,一般每个诱捕器每7d只能诱捕到十几头。8月份随着芒果进入成熟转色期,橘小实蝇种群数量出现第1个高峰,峰值为8月27日观测的756.33头/瓶。9月中旬过后由于各种水果先后成熟,该虫种群数量迅速上升,进入全年发生高峰,最高峰值出现在10月1日观测的2517.33头/瓶。进入11月份,由于寄主的消失和气温的下降,种群数量迅速下降,12月以后,每个诱捕器每7天只能诱捕到十几头。总之,其种群数量的高峰期在8月初到10月底。城市与乡村交界地域的监测点8个月诱虫总数为11271.00头/瓶。
乡村果园橘小实蝇的种群数量在7月前很低,一般每个诱捕器7d只诱捕到1—2头,8月份其种群数量迅速上升,马宝监测点在9月29日、10月底以及11月17日出现了3个峰值,分别为579.71、500.43和170.38头/瓶。12月份后,种群数量迅速下降,诱捕虫数仅为每瓶十几头。可见,乡村橘小实蝇群数量高峰发生在8月到11月底,8个月的诱虫总数仅为4407.60头/瓶。
从福州市几个监测点情况来看,在总体虫量上,城区>城乡之交区域>乡村区域。城市橘小实蝇种群是城乡之交的1.5倍,是乡村区域的5倍甚至10倍以上,三者之间的差异均达到极显著水平(P=0.000<0.01)。这可能与城区一般没有采取橘小实蝇的防治措施有关。从发生时间来看,城区7月份种群数量开始上升,城乡之交在8月份种群数量开始上升,而乡村果园则要到9月份该虫种群数量才开始上升。这可能是由于城区气温要高于乡村所致。
3 讨论与结论

图5 城市与乡村果园的种群动态的比较Fig.5 The relations of the population dynamics of B.dorsalis between city and country
橘小实蝇的种群动态和寄主植物的成熟期关系密切,寄主植物的转色成熟期成为各果园橘小实蝇发生的高峰期,这与已有的研究结果基本一致[5-6]。另外,研究结果显示芭乐园和杨桃园的种群数量最高,而丝瓜和黄瓜园中的橘小实蝇种群数量要明显低于其他果园,种群数量存在显著的差异,充分说明橘小实蝇对葫芦科等1年生藤本植物的喜好不如石榴科等多年生瓜果类植物,这可能与橘小实蝇对不同水果气味的趋向性不同有关[19-20],具体原因还需通过室内试验予以进一步揭示。此外,本研究还发现混合果园的橘小实蝇种群发生期要明显长于单一果园,且发生量更大。因此在实际生产中一定要保持果园、瓜园中瓜果品种的单一性,尽量避免各类瓜果混合栽培。不仅如此,研究结果还表明,在水果的转色、成熟期,果园内种群数量高于果园周边丛林的种群数量。当水果的转色、成熟期过后,果园内部的种群数量小于果园周边的种群数量。这可能与果园周边丛林的植物种类繁多,多杂果,且丛林灌木密集,湿度较大有关。引起果园与周边种群数量变化的原因还有待于进一步探讨。
山谷与山脊生境的橘小实蝇种群有极显著差异,山谷生境的种群数量明显高于山脊生境。这可能与山谷生境较山脊生境阴凉、小气候较稳定、昼夜温差小,适合该虫的生存与活动有关[17-18]。由于山谷生境和山脊生境在种群数量方面存在显著的正相关,所以山谷生境橘小实蝇种群的增长会引起山脊生境其种群的快速增长;同样,沿河生境相对湿度较高、小气候较稳定,因此,沿河生境的种群要显著高于非沿河生境。为此,田间小气候在害虫的测报防治方面应予以充分考虑。
关于城区和乡村橘小实蝇种群消长动态的差异,目前还缺乏相关的研究报道。本文对此进行了初步的探讨。结果表明,城区橘小实蝇的发生期早于乡村,发生高峰期略早于乡村。主要原因在于在南方许多行道树多为芒果树,是橘小实蝇的嗜好寄主。此外,城区没有对该虫进行有效的防治措施且平均气温要高于乡村,为该虫提供很好的越冬场所,政府和检疫部门对此应予以重视。
本文系统研究了不同生境条件下橘小实蝇的种群消长动态,但由于受到试验条件的限制,本研究仅对各生境条件橘小实蝇种群动态进行了初步探讨,试验存在取样点数量不足的问题。如城区与郊区果园的橘小实蝇的种群动态的研究由于取样点的不足无法精确反映城乡橘小实蝇种群数量的时空动态变化特征。各生境橘小实蝇种群的时空动态变化特征有待深入研究。
[1]Steiner L F.Field evaluation of oriental fruit fly insecticides in Hawaii.Journal of Economic Entomology,1957,50:16-24.
[2]Chen P,Ye H.Advances in Taxonomy of Bactrocera dorsalis complex.Entomological Knowledge,2007,44(1):41-47.
[3]Zhan K R,Zhao SX,Zhu S F,Zhou W C,Wang N W.Study on Viability of Bactrocera doraslis in China.Journal of South China Agricultural University,2006,27(4):21-25.
[4]Fan JA.Studies on the assessment of potential establishment of oriental fruit(Bactrocera dorsalis)in China with synthetic judgement on Fuzzy Mathematics.Plant Quarantine,1998,12(2):76-80.
[5]Yuan SY,Kong Q,Xiao C,Chen B,Li Z Y,Zhu C Y.Study on the population fluctuation of Bactrocera dorsalis Hendel in Honghe megranate orchard.Journal of Mountain Agriculture and Biology,2005,24(3):217-220.
[6]Xu J L,Han SC,Ou J F,Huang H,Wu H.A preliminary report on the integrated control of Bactrocera dorsalis in Carambola orchard.Natural Enemies of Insects,2003,25(3):105-112.
[7]Zhu C G,Zhang S F,Xia X N.The population dynamics and the correlation with environmental factors of Bactrocera dorsalis(Diptera:Tephritidae)Occurring in the green spaces.Acta Phytophylacica Sinica,2008,35(5):389-393.
[8]Ye H,Liu J H.Population dynamics of Bactrocera dorsalis(Diptera:Tephritidae)in Xishuangbanna of Southern Yunna.Chinese Journal of Applied Ecology,2005,16(7):1330-1334.
[9]Liu JH,Ye H.Population Dynamics of Bactrocera dorsalis(Diptera:Tephritidae)in Yuanjiang Dry-hot Valley,Yunnan with an analysis of the related factors.Acta Entomologica Sinica,2005,48(5):706-711.
[10]Chen P,Ye H,Liu J H.Population dynamics of Bactrocera dorsalis(Diptera:Tephritidae)along with analysis on the factors influencing the population in Ruili,Yunnan Province.Acta Ecologica Sinica,2006,26(9):2801-2809.
[11]Chen P,Ye H.Population dynamics of Bactrocera dorsalis(Diptera:Tephritidae)in Liuku,Yunnan with an analysis of the influencing factors.Acta Entomologica Sinica,2007,50(1):38-45.
[12]LüX,Han S C,Xu J L,Huang H,Wu H,Ou J F,Sun L.Population dynamics of Bactrocera dorsalis(Hendel)in Guangzhou,Guangdong province,with analysis of the climate factors.Acta Ecologica Sinica,2008,28(4):1850-1856.
[13]Liu L,Yu L.Population Dynamics of Bactrocera dorsalis and Its Impact Factors.Guizhou Agriculture Sciences,2010,38(8):100-102.
[14]Gai JY.The Methods of Test Statistics.Beijing:China Agriculture Press,2000:86-88,118-124,227-231.
[15]Alyokhin A V,Mille C,Messing R H,Duna J J.Selection of pupation habitats by oriental fruit fly larvae in the laboratory.Journal of Insect Behavior,2001,14(1):57-67.
[16]Fletcher B S.Life History Strategies of Tephritid fruit flie//Robinson A S,Hooper G,eds.World Crop Pests,Vo1.3(B).Amsterdam:Elsevier,1989:195-219.
[17]Liu J H,Ye H.Effects of light,temperature and humidity on the flight activities of the oriental fruit fly,Bactrocera dorsalis.Entomological Knowledge,2006,43(2):211-214.
[18]Huang J C,Zheng S N,Ye G L,Cheng J H.The effect of different seasons and meteorological factors on the daily flight activity rhythm of Bactrocera dorsalis(Hendel).Chinese Journal of Tropical Crops,2011,32(10):1926-1930.
[19]Wang B,Huan Y,Huang JC,Cheng J H.Bioassay of botanical attractants to Bactrocera dorsalis.Chinese Journal of Applied Entomology,2012,49(6):1547-1552.
[20]Shi W,Zhang Z Y,Ye H.Taxis responses of three oriental fruit fly(Bac rocera dorsalis Hendel)populations to the odors of three host fruits.Journal of Yunnan University:Natural Sciences,2003,25(1):77-80.
参考文献:
[2]陈鹏,叶辉.橘小实蝇复合体分类学研究进展.昆虫知识,2007,44(1):41-47.
[3]詹开瑞,赵士熙,朱水芳,周卫川,王念武.橘小实蝇在中国的适生性研究.华南农业大学学报,2006,27(4):21-25.
[4]范京安.用模糊综合评判法探讨桔小实蝇在中国的适生分布.植物检疫,1998,12(2):76-80.
[5]袁盛勇,孔琼,肖春,陈斌,李正跃,朱春燕.红河石榴园橘小实蝇种群动态规律.山地农业生物学报,2005,24(3):217-220.
[6]徐洁莲,韩诗畴,欧剑峰,黄鸿,吴华.杨桃园桔小实蝇综合防治初报.昆虫天敌,2003,25(3):105-112.
[7]朱春刚,张守锋,夏希纳.园林绿地中桔小实蝇种群消长与环境因子的相关性.植物保护学报,2008,35(5):389-393.
[8]叶辉,刘建宏.云南西双版纳桔小实蝇种群动态.应用生态学报,2005,16(7):1330-1334.
[9]刘建宏,叶辉.云南元江干热河谷桔小实蝇种群动态及其影响因子分析.昆虫学报,2005,48(5):706-711.
[10]陈鹏,叶辉,刘建宏.云南瑞丽桔小实蝇成虫种群数量变动及其影响因子分析.生态学报,2006,26(9):2801-2809.
[11]陈鹏,叶辉.云南六库桔小实蝇成虫种群数量变动及其影响因子分析.昆虫学报,2007,50(1):38-45.
[12]吕欣,韩诗畴,徐洁莲,黄鸿,吴华,欧剑峰,孙力.广州桔小实蝇(Bactrocera dorsalis(Hende1))发生动态及气象因子.生态学报,2008,28(4):1850-1856.
[13]刘莉,余丽.云南镇康地区桔小实蝇种群动态及其影响因子.贵州农业科学,2010,38(8):100-102.
[14]盖钧镒.试验统计方法.北京:中国农业出版社,2000:86-88,118-124,227-231.
[17]刘建宏,叶辉.光照、温度和湿度对橘小实蝇飞翔活动的影响.昆虫知识,2006,43(2):211-214.
[18]黄居昌,郑思宁,叶光禄,陈家骅.不同季节与气象因素对橘小实蝇日飞翔活动节律的影响.热带作物学报,2011,32(20):1926-1930.
[19]王波,韩英,黄居昌,陈家骅.橘小实蝇植物源引诱活性物质的生物测定.应用昆虫学报,2012,49(6):1547-1552.
[20]施伟,张智英,叶辉.桔小实蝇对寄主水果气味的趋向反应测试.云南大学学报:自然科学版,2003,25(1):77-80.
