兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性
2013-12-09季子敬全先奎王传宽
季子敬,全先奎,王传宽
(东北林业大学生态研究中心,哈尔滨 150040)
兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性
季子敬,全先奎,王传宽*
(东北林业大学生态研究中心,哈尔滨 150040)
叶片易受环境因子影响,其形态解剖结构特征不但与叶片的生理功能密切相关,而且反映树木对环境变化的响应和适应。叶片结构的改变势必会改变树木的生理功能。同一树种长期生长在异质环境条件下,经过自然选择和适应,会在形态和生理特性等方面产生变异,形成特定的地理种群。另外,母体所经受的环境胁迫也会影响到其子代的生长、发育和生理等特征。因此,了解植物叶片形态结构对环境变化的响应与适应是探索植物对环境变化的响应和适应机制的基础。兴安落叶松(LarixgmeliniiRupr.)是我国北方森林的优势树种,主要分布在我国东北地区。为了区分叶片对气候变化的可塑性和适应性,采用同质园法比较测定了6个不同气候条件下的兴安落叶松种源的32年生树木的针叶解剖结构和光合生理相关因子,分析了针叶的解剖结构特征、光合能力(Pmax-a)、水分利用效率(WUE)之间的关系及其对气候变化的适应性。结果表明:表皮细胞厚度、叶肉细胞厚度、传输组织厚度、维管束厚度、内皮层厚度以及叶片总厚度均存在显著的种源间差异(Plt;0.05)。叶肉细胞厚度与Pmax-a、气孔导度和WUE之间均存在显著的正相关关系(Plt;0.05)。叶肉细胞厚度、表皮细胞厚度、叶片总厚度以及叶肉细胞厚度和表皮细胞厚度在叶片总厚度中所占比例均与种源地的干燥度指数(即年蒸发量与年降水量之比)呈正线性关系。这些结果说明:不同种源兴安落叶松针叶解剖结构因对种源原地气候条件的长期适应而产生显著的差异,从而引起其针叶光合作用、水分利用等生理功能发生相应的变化,从而有利于该树种在气候变化的情景下得以生存和繁衍。
气候变化;种源;叶片解剖;光合特性;水分利用效率
叶片在植物进化过程中作为可塑性较大并容易受环境因子影响的器官[1],其结构特征最能体现环境因子的影响或植物对环境的适应。叶片结构的改变势必会改变树木的生理功能[2]。例如,韩梅等[3]研究叶片解剖特征沿温度梯度变化时发现,不同功能型植物叶片各组织厚度对CO2浓度和温度升高的响应存在明显的种类差异。光照强度的差异会导致植物叶片的解剖结构发生变化,从而影响植物的生长发育;另外,干旱频发、水分短缺等也会严重影响植物的生长发育,使叶片的表皮细胞变薄,气孔数量和开度发生变化,以便适应干旱的环境条件[2]。因此,了解植物叶片形态结构对环境变化的响应与适应是探索植物对环境变化的响应适应机制的基础。
同一树种长期生长在异质环境条件下,经过自然选择和适应,会在形态和生理特性等方面产生变异,形成特定的地理种群[4]。叶片解剖结构特征受树木生理状况和环境因子的共同限制,并随着环境条件的变化而呈现一定的变化规律。以往的研究表明,来源于不同自然环境的种群在叶片大小、形状等方面存在一定差异[5]。在同质园内,来自于高纬度/海拔种源树木具有较高的比叶重(Leaf Mass per Area, LMA),这可能是一种基因调控下的对种源原地环境适应的表现[6- 10];而LMA的种源差异往往是由叶片结构的改变而引起的,至于有哪些结构特征参数发生何种变化仍不清楚[6,8]。另外,母体所经受的环境胁迫也会影响到其子代的生长、发育和生理等特征[11],但这种母体效应的影响程度和机理还有待深入研究。这些问题的回答,对于认识树种叶片解剖结构对气候变化的响应和适应特征具有重要意义。
叶片的解剖结构与植物的光合能力密切相关。通常,叶片厚度的增加会使其光合速率升高[12]。以往对于阔叶植物的研究表明:叶肉细胞增厚,栅栏组织加厚,细胞层次增多,可使叶片光合能力增强[13- 14]。但对于针叶树叶片结构与光合的关系报道甚少。马立祥[15]等认为不同地理种群的两针松(即Pinusdensiflora、Pinussylvestrisvar.sylvestriformisandPinussylvestrisvar.mongolica)的光合能力存在差异,但对于处在不同地理条件下的同一种群针叶的解剖特性是否存在差异、以及这种差异是植物对环境变化的驯化还是遗传调控下的适应等问题还不太清楚。
兴安落叶松(LarixgmeliniiRupr.)是我国北方森林的优势树种,主要分布在我国东北地区,但日益加剧的气候变化可能会改变其现有的分布区。例如,据张新时[16]预测,如果气温增加2—4 ℃、年降水量增加20%的条件下,兴安落叶松可能北移,取而代之的是温带草原和针阔混交林。然而,类似的模型预测尚未考虑树木的形态、解剖结构、生理功能等对气候变化的潜在响应和适应机制。为此,本研究采用同质园法比较测定了6个不同气候条件下的兴安落叶松种源的32年生树木的叶片解剖结构和光合生理相关因子,旨在比较分析不同种源针叶解剖结构变化与其光合生理特征的关系,为预测气候变化对北方森林生态系统的影响提供理论基础。
1 研究材料与方法
1.1 实验地自然概况
本实验所用的同质种植园位于东北林业大学帽儿山森林生态站(127 ° 30′—34 ′E, 45 ° 20 ′—25 ′N)。在兴安落叶松国内的分布范围内,在以表征能量差异为主的纬度梯度上选取代表性种源6个(表1),于1980年秋采种,1981年育苗,1983年春造林,1997年和2001年间伐两次,株行距现为4.5 m × 2.5 m。同质园内土壤、地形、气候条件一致,平均海拔300 m,平均坡度10—15 °,土壤为暗棕壤,年平均降雨量629 mm,年均蒸发量864 mm,年平均温度3.1 ℃,1月和7月份平均温分别为-18.5 ℃和22 ℃,无霜期120—140 d(5—9月份)[17]。

表1 兴安落叶松种源原地地理气象条件及生长现状
表中的a、b、c表示HSD检验显著性差异组别(α=0.05); MAT:mean annual air temperature, MAP:mean annual precipitation, MAE:mean annual evaporation, DBH:diameter at breast height, and LMA:leaf mass area
1.2 光合能力及相关因子的测定
在同质园内,每个种源选取3株标准木,搭建触及冠层(14 m左右)的脚手架,以便活体测量针叶的光合作用及其相关因子。每株标准木选取冠层上部向阳方的当年生枝新生叶3簇,于2011年8月中旬,选择无雨天气于6:30—12:00时段,采用Li- 6400便携式CO2/H2O红外分析仪(Li-Cor, NB, USA)活体测定针叶气体交换过程以及相应的环境因子。测定时采用的叶室环境控制为:温度25 ℃,流速500 μmol/s,CO2浓度400 μmol CO2/mol。测定前采用饱和光强对叶片进行充分光诱导。在测定光响应曲线时,光量子通量密度梯度为: 2000、1500、1200、800、400、200、150、100、50、0 μmol 光子量·m-2·s-1;测定输出参数包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(gs)等。用于光合测定的针叶面积采用扫描图像处理获得[18];随后将叶片在65 ℃下烘至恒重称重(精度0.0001),获取其比叶重(LMA,g/m2):
LMA=叶片干重/叶面积
(1)
1.3 叶片解剖方法
在同质园内各种源每株标准木上选取与测定光合能力叶片位置相近、发育程度一致的健康针叶各10簇,用FAA固定液(酒精∶福尔马林∶冰醋酸=90∶5∶5)固定。样品经系列酒精脱水后,二甲苯透明、浸蜡、包埋,然后用旋转式薄片切片机(Rotary Microtome, Leitz, Germany)制片。在每个针叶中部取10个横向切片,切片厚度为8—10 μm。切片脱蜡后,用番红-固绿对染,即用1%的番红水溶液染色12 h, 再用0.5%固绿染色,然后用阿拉伯树胶封片,用ZEISS Imager A1型光学摄影显微镜观察摄影;之后用Axio Vision Release 4.5 SP1测量叶片横切面照片的叶片厚度(LD)、上表皮细胞厚度(LUPD)、下表皮细胞厚度(LBPD)、近轴面叶肉细胞厚度(LUCD)、远轴面叶肉细胞厚度(LBCD)、维管束厚度(LVD)、内皮层厚度(LED)和传输组织厚度( LTD);每个测量指标重复测量3次。表皮细胞厚度(LPD)和叶肉细胞厚度(LCD)分别由下式获得:
LPD=LUPD + LBPD
(2)
LCD=LUCD + LBCD
(3)
1.4 数据分析
光合作用的光响应曲线采用Prado等[19]的非线性拟合方程:
Pn=Pmax-a(1-e-k(PAR-P))
(4)
式中,Pmax-a为单位叶面积最大净光合速率(μmol CO2·m-2·s-1),k是常数,Pn是实际测定的净光合速率,PAR是光合有效辐射,P是光补偿点。
叶片尺度的水分利用效率(WUE, μmol CO2/mmol H2O )由下式[20]获得:
WUE=Pn/Tr
(5)
式中,Tr为蒸腾速率。
干燥度指数(AI)由下式[21]获得:
AI=MAE/MAP
(6)
式中,MAE为平均年蒸发量;MAP为平均年降水量。
数据分析和差异性检验采用SPSS 19.0(SPSS Inc., Chicago 2001)统计软件分析;所有图采用Sigma Plot 9.0(Systat corporation, USA)绘制。
2 结果
2.1 叶片解剖结构的种源差异
兴安落叶松叶片厚度(LD)及各解剖结构特征厚度均存在显著种源差异(图1,Plt;0.05)。LD的大小顺序为:中央站((392.8±9.8) μm,平均值±标准误)gt; 根河(383.0±7.5) μmgt; 鹤北(375.3±8.5) μmgt; 三站(373.7±3.7) μmgt; 塔河(362.5±5.3) μmgt; 乌伊岭(354.3±6.8) μm;来自寒冷干燥地区的种源具有较高的LD。比较叶片各解剖结构特征参数的大小时发现,来自中央站种源的叶片均具有较高的厚度,而来自低纬度乌伊岭种源叶片则均具有较小的厚度,其它种源因解剖结构特征参数而具有不同的种源差异。

图1 六个种源兴安落叶松的叶片结构特征参数Fig.1 Parameters of leaf anatomy of the six provenancesLD:叶片厚度Leaf thickness,LUPD:上表皮细胞厚度Leaf adaxial epidermis cells thickness,LBPD:下表皮细胞厚度Leaf abaxial epidermis cells thickness,LUCD:近轴面叶肉细胞厚度Leaf adaxial mesophyll cells thickness,LBCD:远轴面细胞厚度Leaf abaxial mesophyll cells thickness,LED:内皮层厚度Leaf endoderm cells thickness,LTD:传输组织厚度Leaf transporting tissue thickness,LVD:维管束厚度Leaf vascular bundle tissue thickness
叶片各解剖结构特征厚度在叶片总厚度中所占的比例差异明显(图2),其中叶肉细胞厚度(LCD)所占比例最大,其平均值波动在(38.4%—43.3%)之间;而表皮细胞厚度(LPD)所占比例为最小,其平均值波动范围为(5.1%—6.1%);维管束厚度(LVD)波动在(16.15%—19.0%)之间;传输组织厚度(LTD)波动范围为(12.95%—18.2%);而内皮层厚度(LED)则在(11.5%—15.3%)之间波动。

图2 叶片各解剖结构特征参数占叶片总厚度的比例 Fig.2 The proportion of the parameters of leaf anatomy to the total leaf thickness
2.2 叶片解剖结构与生理功能之间的关系
LD、LCD和LTD均与最大净光合速率(Pmax-a)、气孔导度(gs)及水分利用效率(WUE)之间存在显著的正相关关系(图3;Plt;0.05;LTD与gs除外)。总体而言,中央站和根河种源叶片具有较高的Pmax-a、WUE和gs,而乌伊岭和塔河种源的各测定生理特征参数值较低。且种源间LCD与各测定生理特征间相关性要高于LD和LTD。种源间Pmax-a、gs及WUE波动范围分别为(9.60±0.86)—(16.61±0.91) μmol CO2·m-2·s-1,(0.20±0.002)—(0.30±0.009) mmol CO2·m-2·s-1和(2.06±0.08)—(3.83±0.05) μmol CO2·mmol-1H2O。
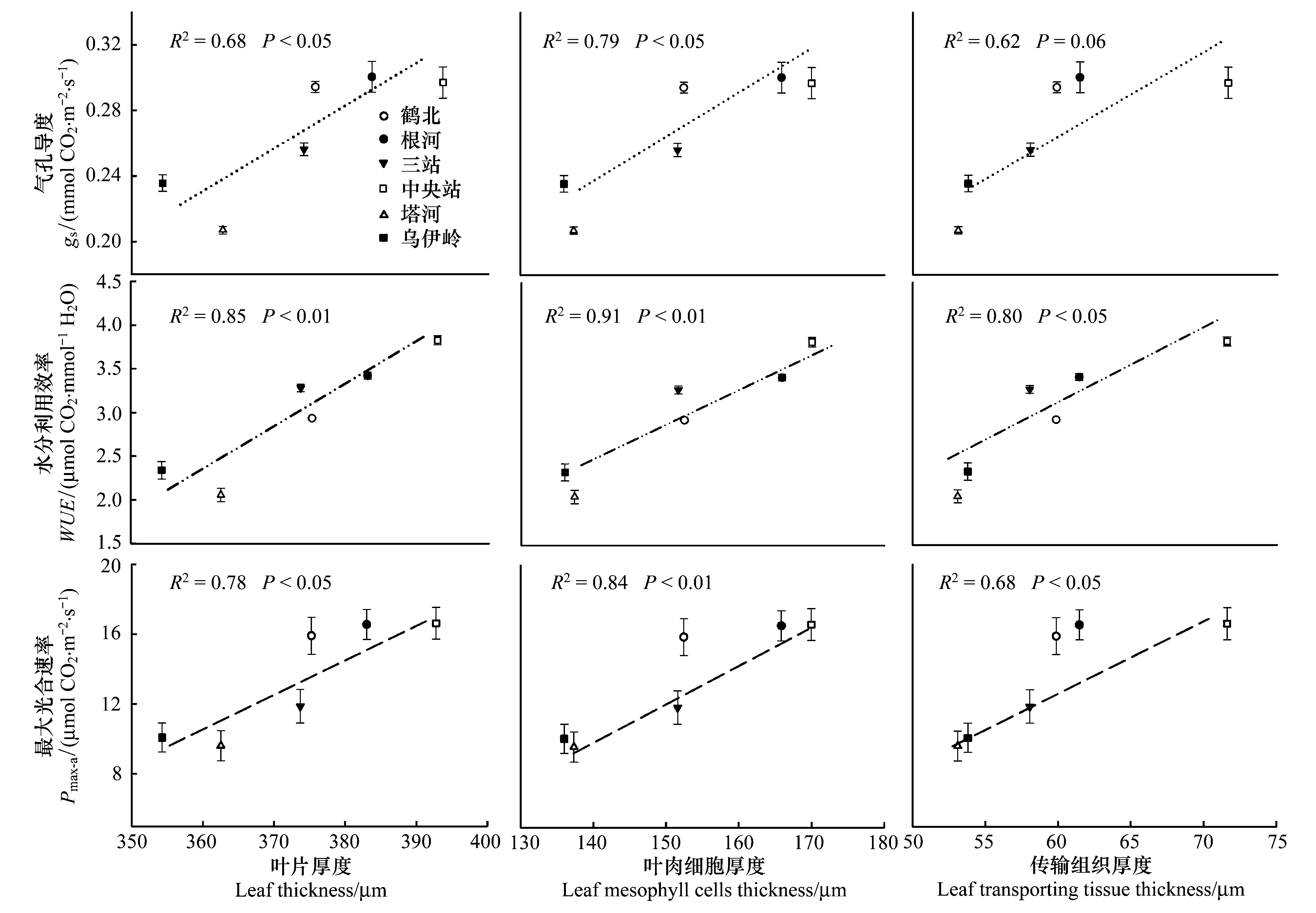
图3 叶片解剖结构特征参数和最大净光合速率(Pmax-a)、水分利用效率(WUE)、气孔导度(gs)之间的关系(n=6)Fig.3 Relationships between parameters of leaf anatomy and Pmax-a、WUE and gs(n=6)
2.3 叶片解剖结构与种源地干燥度指数之间的关系
LD、LCD和LPD均与种源原地干燥度指数(AI)之间呈线性相关,其中AI和LCD之间的相关性最紧密(表1,图4)。随着种源原地的干燥程度增加,LD、LCD和LPD均明显增大,且LCD和LPD在LD中所占比例也随AI的增加而显著增大(图4)。
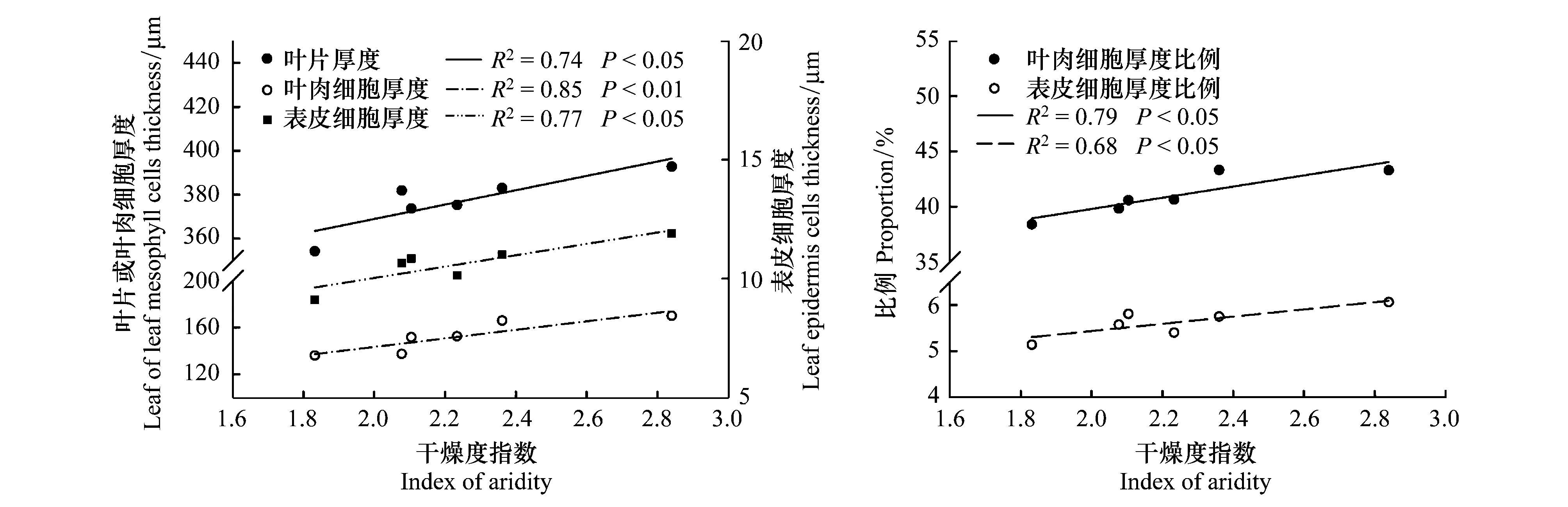
图4 叶片解剖结构特征参数与种源原地干燥度指数之间的关系(n=6)Fig.4 Relationships between parameters of leaf anatomy and the aridity index at the original sites (n=6)
3 讨论
6个种源兴安落叶松针叶的解剖结构特征参数差异显著(图1),因而导致了针叶总厚度(LD)和LMA的种源间差异(图1,表1)。在各解剖结构特征参数中,叶肉细胞最为突出,其厚度约占LD的40%左右,且与LD呈现相似的种源间差异。因为6种源兴安落叶松已经在环境条件一致的同质园内生长了32a,所以可以认为这种解剖结构上的变化是树木对种源原地气候条件长期适应的结果。进一步通过与原地气候因子进行回归分析发现,LD、LCD、LPD以及LCD和LPD占LD的比例均与种源原地的干燥度指数(AI)呈线性关系(图4)。虽然其它解剖特征参数与AI相关没有达到显著性水平,但总体趋势不变,即来自寒冷干燥的种源具有较厚的叶片以及较大的各解剖结构特征参数。杨传平等[4]对该6个种源兴安落叶松的前期研究也发现,其材积、树高、胸径等等形态指标也存在显著的种源间差异。
叶片的形态、解剖结构和生长发育是受遗传和环境条件(如:地理位置、生境条件和气候特性等)共同调控的[22- 23],其形态和结构特征的改变能够反映树木对环境变化的响应和适应。Glloway[11]对风铃草(Campanulaamericana)的研究显示,母体对后代如何适应当地环境条件有暗示作用,即生长在与母体相同环境下的后代植株比其他环境下的后代表现更为出色。刘全宏[24]对太白冷杉(Larixchinensis)的研究报道,随着年降水量的增加,叶片厚度、传输组织、维管束厚度和内皮层厚度呈减小或变薄的趋势。在干燥条件下,叶片往往通过增加叶片厚度尤其是表皮厚度防止水分的过度蒸发,进而保证植物的正常代谢。Huner[25]等的研究表明,寒冷地区叶片具有较厚的表皮细胞,这正是植物御寒的一种表现。
结构是功能的基础,叶片结构特征的变化将会影响树木的光合、呼吸、蒸腾等生理功能。研究结果显示,Pmax-a和WUE均与LD、LCD和LTD呈显著的正线性关系(图3),与Oguchi等[26]的研究结果一致。也有研究报道,来自寒冷高纬度的种源具有较强的光合能力,且与叶片的LMA显著相关[6- 8,27]。叶片及其叶肉组织增厚,叶绿体含量也相应地升高(数据未显示),为光合作用提供了更多的场所,进而提高了光合能力;同时,也提高了植物的水分利用效率[28](图3),从而增强了树木抵御干旱等环境胁迫的能力。
总之,兴安落叶松长期适应当地的环境条件,使得其针叶的形态特征、解剖结构特征和生理功能上产生具有遗传能力的变异,是其在严酷的北方森林生境内得以生存繁衍的生态对策之一。
4 结论
本研究利用同质园实验有效地区分了兴安落叶松针叶解剖结构和光合生理对气候变化响应的可塑性和适应性。针叶的解剖结构特征参数存在显著的种源间差异,而且叶肉细胞厚度与针叶的光合能力和水分利用效率之间均存在显著的正相关关系。叶肉细胞厚度、表皮细胞厚度、叶片总厚度以及叶肉细胞厚度和表皮细胞厚度在叶片总厚度中所占比例均与种源原地的干燥度指数呈正线性关系。这表明兴安落叶松长期适应当地的环境条件,在针叶的形态特征、解剖结构特征和生理功能上产生具有遗传能力的变异,从而有利于该树种在气候变化的情景下得以生存和繁衍。
致谢: 感谢黑龙江帽儿山森林生态系统国家野外科学观测研究站和林木遗传育种国家重点实验室(东北林业大学)提供的野外基础支持。
[1] Wang X L, Wang J. Plant Morphology and Environment. Lanzhou: Lanzhou Publication, 1989, 105- 138.
[2] Li F L, Bao W K. Responses of the morphological and anatomical structure of the plant leaf to environmental change. Chinese Bulletin of Botany, 2005, 22(Supplement): 118- 127.
[3] Han M, Ji C J, Zuo W Y, He J S. Interactive effects of elevated CO2and temperature on the leaf anatomical characteristics of eleven species. Acta Ecologica Sinica, 2006, 26(2): 326- 333.
[4] Yang C P, Jiang J, Tang S S, Li J Y, Wang H R. The provenance test of 21-year oldLarixgmeliniiat Maoershan area. Journal of Northeast Forestry University, 2002, 30(6): 1- 5.
[5] Saxe H, Cannell M G R, Johnsen Ø, Ryan M G, Vourlitis G. Tree and forest functioning in response to global warming. New Phytologist, 2001, 149(3): 369- 400.
[6] Cordell S, Goldstein G, Mueller-Dombois D, Webb D, Vitousek P M. Physiological and morphological variation inMetrosiderospolymorpha, a dominant Hawaiian tree species, along an altitudinal gradient: the role of phenotypic plasticity. Oecologia, 1998, 113(2): 188- 196.
[7] Bresson C C, Vitasse Y, Kremer A, Delzon S. To what extent is altitudinal variation of functional traits driven by genetic adaptation in European oak and beech? Tree Physiology, 2011, 31(11): 1164- 1174.
[8] Oleksyn J, Modrzynski J, Tjoelker M G, Reich P B, Karolewski P. Growth and physiology ofPiceaabiespopulations from elevational transects: common garden evidence for altitudinal ecotypes and cold adaptation. Functional Ecology, 1998, 12(4): 573- 590.
[9] Oleksyn J, Tjoelker M G, Reich P B. Adaptation to changing environment in Scots pine populations across a latitudinal gradient. Silva Fennica, 1998, 32(2): 129- 140.
[10] Robson T M, Sánchez-Gómez D, Cano F J, Aranda I. Variation in functional leaf traits among beech provenances during a Spanish summer reflects the differences in their origin. Tree Genetics amp; Genomes, 2012, 8(5): 1111- 1121.
[11] Galloway L F. Parental environmental effects on life history in the herbaceous plantCampanulaamericana. Ecology, 2001, 82(10): 2781- 2789.
[12] Oguchi R, Hikosaka K, Hirose T. Leaf anatomy as a constraint for photosynthetic acclimation: differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees. Plant, Cell and Environment, 2005, 28(7): 916- 927.
[13] Hanba Y T H. Kogami H, Terashima I. The effect of growth irradiance on leaf anatomy and photosynthesis inAcerspecies differing in light demand. Plant, Cell and Environment, 2002, 25(8): 1021- 1030.
[14] Wang X L. The development of plant ecological anatomy. Chinese Bulletin of Botany, 1993, 10, 1- 10.
[15] Ma L X, Wang X W, Mao Z J, Liu L X, Sun T. Variations in photosynthetic capacity and growth characters for 2-needled pine from diverse geographic populations. Bulletin of Botanical Research, 2010, 30(6): 680- 684.
[16] Zhang X S. A vegetation-climate classification system for global change studies in China. Quaternary Sciences, 1993, (2): 157- 169.
[17] Sang Y R, Wang C K, Huo H. Inter-specific and seasonal variations in photosynthetic capacity and water use efficiency of five temperate tree species in northeastern China. Scandinavian Journal of Forest Research, 2011, 26(1): 21- 29.
[18] Xiao Q, Ye W J, Zhu Z, Chen Y, Zheng H L. A simple non-destructive method to measure leaf area using digital camera and Photoshop software. Chinese Journal of Ecology, 2005, 24(6): 711- 714.
[19] Prado C H B A, De Moraes J A P V. Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field conditions. Photosynthetica, 1997, 33(1): 103- 112.
[20] Wang J L, Yu G R, Fang Q X, Jiang D F, Qi H, Wang Q F. Responses of water use efficiency of nine plant species to light and CO2and it′s modeling. Acta Ecologica Sinica, 2008, 28(2): 525- 533.
[21] Meng M, Ni J, Zhang Z G. Aridity index and its applications in geo-ecological study. Acta Phytoecologica Sinica, 2004, 28(6): 853- 861.
[22] Xu D C, Lü F D, Li B, Jiang Y. Comparison of chlorophyll fluorescence characteristics and leaf characters of pecan in different site conditions. Nonwood Forest Research, 2005, 23(4): 17- 20.
[23] Peng S B, Kriegdr D R, Girma F S. Leaf photosynthetic rate is correlated with biomass and grain production in grain sorghum lines. Photosynthesis Research, 1991, 28(1): 1- 7.
[24] Liu Q H, Wang X A, Tian X H, Xiao Y P. Morphological and anatomical characteristics of leaf ofLarixchinensisand their relationship to environmental factors in Taibaishan Mountain. Acta Botanica Boreali-Occidentalia Sinica, 2001, 21(5): 885- 893.
[25] Huner N P A, Palta J P, Li P H, Carter J V. Anatomical changes in leaves of Puma Rye in response to growth at cold-hardening temperatures. Botanical Gazette, 1981, 142(1): 55- 62.
[26] Oguchi R, Hikosaka K, Hirose T. Does the photosynthetic light-acclimation need change in leaf anatomy? Plant, Cell and Environment, 2003, 26(4): 505- 512.
[27] Scheepens J, Frei E S, Stöcklin J. Genotypic and environmental variation in specific leaf area in a widespread Alpine plant after transplantation to different altitudes. Oecologia, 2010, 164(1): 141- 150.
[28] Masle J, Farquhar G D. Effects of soil strength on the relation of water-use efficiency and growth to carbon isotope discrimination in wheat seedlings. Plant Physiology, 1988, 86(1): 32- 38.
参考文献:
[1] 王勋陵, 王静. 植物形态结构与环境. 兰州: 兰州大学出版社, 1989: 105- 138.
[2] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 2005, 22(增刊): 118- 127.
[3] 韩梅, 吉成均, 左闻韵, 贺金生. CO2浓度和温度升高对11种植物叶片解剖特征的影响. 生态学报, 2006, 26(2): 326- 333.
[4] 杨传平, 姜静, 唐盛松, 李景云, 王会仁. 帽儿山地区21年生兴安落叶松种源试验. 东北林业大学学报, 2002, 30(6): 1- 5.
[14] 王勋陵. 植物生态解剖学研究进展. 植物学通报, 1993, 10: 1- 10.
[15] 马立祥, 王秀伟, 毛子军, 刘林馨, 孙涛. 不同地理种群两针松光合和生长特性的差异. 植物研究, 2010, 30(6): 680- 684.
[16] 张新时. 研究全球变化的植被——气候分类系统. 第四纪研究, 1993, (2): 157- 169.
[18] 肖强, 叶文景, 朱珠, 陈瑶, 郑海雷. 利用数码相机和Photoshop软件非破坏性测定叶面积的简便方法. 生态学杂志, 2005, 24(6): 711- 714.
[20] 王建林, 于贵瑞, 房全孝, 姜德锋, 齐华, 王秋凤. 不同植物叶片水分利用效率对光和CO2的响应与模拟. 生态学报, 2008, 28(2): 525- 533.
[21] 孟猛, 倪健, 张治国. 地理生态学的干燥度指数及其应用评述. 植物生态学报, 2004, 28(6): 853- 861.
[22] 徐德聪, 吕芳德, 栗彬, 蒋瑶. 不同立地美国山核桃叶绿素荧光特性及叶性状比较. 经济林研究, 2005, 23(4): 17- 20.
[24] 刘全宏, 王孝安, 田先华, 肖娅萍. 太白红杉(Larixchinensis)叶的形态解剖学特征与环境因子的关系. 西北植物学报, 2001, 21(5): 885- 893.
VariationsinleafanatomyofLarixgmeliniireflectadaptationofitsphotosyntheticcapacitytoclimatechanges
JI Zijing, QUAN Xiankui, WANG Chuankuan*
CenterforEcologicalResearch,NortheastForestryUniversity,Harbin150040,China
Leaf anatomy, being susceptible to environmental factors, is closely related to physiological functions of the leaf, and also reflects responses and adaptation of the tree to environmental changes. Changes in leaf anatomical structure are bound to change the physiological functions of trees. The same species growing in heterogeneous environment for a long time through natural selection and adaptation will result in variations in morphological and physiological characteristics, forming distinctive geographic populations. In addition, Matrix subjected to environmental stress will affect its offspring growth, developmental and physiological characteristics. Therefore, to understand how the morphological structure of leaves respond and adapt to environmental changes is essential for exploring the response and adaptation mechanism of plants to environmental changes. Dahurian larch (LarixgmeliniiRupr.) is a dominant tree species in boreal forest in China, mainly distributed in northeast China, but going further climate changes may change its existing distribution. There are few studies to distinguish short-term responses of leaf anatomy and physiology from their potential long-term adaptation to climate changes in field. In this study, we conducted a common garden experiment that included six provenances of Dahurian larch with different climate and site conditions with three replicate trees each provenance. The trees were 32 years old, seeded in 1980, and transplanted in 1983 at identical site and climate conditions. We measured leaf anatomical parameters using paraffin section method (i.e., total leaf thickness, leaf adaxial epidermis cells thickness, leaf abaxial epidermis cells thickness, leaf adaxial mesophyll cells thickness, leaf abaxial mesophyll cells thickness, leaf vascular bundle tissue thickness, leaf endoderm cells thickness, leaf transporting tissue thickness, leaf epidermis cells thickness, leaf mesophyll cells thickness, and photosynthetic physiology (i.e., photosynthetic capacity [Pmax-a], stomatal conductance [gs], water use efficiency [WUE]) in order to address the question of whether leaf anatomical structure adapts to climate changes and consequently results in physiological adaptation. The results showed that the thickness of epidermis cells, mesophyll cells, transmission tissue cells, vascular cells and endothelial cells, and the total thickness of the needles differed significantly among the provenances (Plt;0.05). There was a significant difference in the proportion of thickness of each anatomical part. The thickness of mesophyll cell accounted for the largest proportion, with an average proportion ranging from 38.4% to 43.3%. The thickness of mesophyll cells was positively correlated (Plt;0.05) withPmax-a,gs, andWUE. The thickness of mesophyll cells and epidermal cells, the total thickness of the needles, and the proportions of thickness of mesophyll cells or epidermal cells in the total thickness of the needle were all positively correlated with the aridity index (the ratio of mean annual evaporation to mean annual precipitation) at the origins of the provenances. These results suggest that needles of the Dahurian larch trees from different provenances alter their anatomical structures significantly due to adaptation to local climate conditions, and accordingly change their physiological functions such as photosynthesis and water use efficiency.
climate change; provenance; leaf anatomy; photosynthetic characteristics; water use efficiency
国家“十二五”科技支撑项目(2011BAD37B01);教育部长江学者和创新团队发展计划(IRT1054);国家林业局重点项目(2006—77);中央高校基本科研业务费专项资金资助项目(DL10BA19)
2013- 01- 15;
2013- 07- 22
*通讯作者Corresponding author.E-mail: wangck-cf@nefu.edu.cn
10.5846/stxb201301150103
季子敬,全先奎,王传宽.兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性.生态学报,2013,33(21):6967- 6974.
Ji Z J, Quan X K, Wang C K.Variations in leaf anatomy ofLarixgmeliniireflect adaptation of its photosynthetic capacity to climate changes.Acta Ecologica Sinica,2013,33(21):6967- 6974.
