三种温带森林大型土壤动物群落结构的时空动态
2013-12-09张雪萍张利敏
李 娜, 张雪萍, 张利敏
(哈尔滨师范大学地理科学学院,黑龙江省普通高等学校地理环境遥感监测重点实验室,哈尔滨 150025)
三种温带森林大型土壤动物群落结构的时空动态
李 娜, 张雪萍, 张利敏*
(哈尔滨师范大学地理科学学院,黑龙江省普通高等学校地理环境遥感监测重点实验室,哈尔滨 150025)
对帽儿山3种典型森林群落大型土壤动物进行了连续6个月的野外调查研究。通过系统分析,共获得大型土壤动物3604只,隶属于3门6纲17目50科。其中正蚓科(Lumbricidae)、线蚓科(Enchytraeidae)和石蜈蚣目(Lithobiomorpha)为优势类群,常见类群11类。结果表明:(1)水平分布上,密度和生物量红松人工林最高,其次为硬阔叶林,蒙古栎林最少;类群数硬阔叶林最多,蒙古栎林最少;香农指数和丰富度指数均为蒙古栎林最高,红松人工林最低;优势度指数与两者相反;均匀度指数蒙古栎林最高,硬阔叶林最低;(2)垂直分布上,个体密度、类群数及生物量均差异显著(Plt;0.001)。3个样地大型土壤动物个体密度表聚性明显;类群数红松人工林自凋落物层向下减少,硬阔叶林和蒙古栎林0—10 cm最多;生物量在0—10 cm土层最大;香农指数随深度增加而减小,优势度指数则相反;(3)在时间变化上,5月和10月个体密度和类群数较多,9月生物量最大;香农指数和优势度指数差异显著(Plt;0.01),其他指数各月间无明显差异;(4)与土壤环境因子关系上,总有机碳含量与类群数、个体密度及生物量显著正相关,容重与香农指数显著负相关;典型对应分析结果表明,不同类群大型土壤动物与环境相关性不同。
大型土壤动物;森林生态系统;群落结构;多样性;时空分布
大型土壤动物(体长gt;2 mm)个体大,种类繁多,数量巨大,是陆地生态系统重要消费者和分解者。它们通过自身的生存、活动和摄食行为调节土壤酶和微生物活性,参与土壤有机质分解和矿化,促进土壤腐殖质的形成,改善土壤结构和质量[1- 4],调控地表植物群落的结构和功能[5];此外,其对环境变化敏感,常作为反映环境变化的指示生物[6],在土壤生态系统物质循环和能量流动中起着至关重要的作用[7]。目前,大型土壤动物日益引起了中外学者的高度关注,研究主要集中在动物区系组成、数量变化[8- 9]、多样性及其评价指标[10]以及大型土壤动物与环境因素的相互关系[11]等方面。
以往研究结果表明,大型土壤动物类群、密度、生物量和多样性等特征受气候、土壤和植被等因子的影响及调控[12]。在森林生态系统中,植被组成对大型土壤动物起着决定性作用,即使同一土壤类型,在不同的空间上,不同植被类型也会通过改变土壤理化环境[13],进而影响土壤动物群落的组成及结构。因此,土壤动物在森林生态系统中具有较大的空间异质性[14]。同时,温带地区光照、温度、水分和植物群落的季节变化十分显著,导致不同群落大型土壤动物群落数量、组成及结构随环境因子的改变而发生季节性变化,因此温带大型土壤动物群落结构具有明显的时间动态。
处于中高纬度地区的东北森林是我国的重点林区,不仅在我国的生态环境建设和生物地球化学循环中具有举足轻重的作用,而且对于全球变化具有重要指示作用。与地上植物群落相比,地下大型土壤动物群落对气候变化更为敏感,因而能够更好地指示全球变化下温带森林生态系统结构与功能的演变特征和发展态势[15]。陈鹏、殷秀琴和张雪萍等学者[16- 17]对于帽儿山大型土壤动物群落进行了大量细致的研究,但关于其时空动态及其主要环境影响因素的研究还相当薄弱。本研究以我国东北地区的帽儿山3种典型森林群落为研究对象,进行连续6个月的野外调查,目的在于探讨温带不同植被类型大型土壤动物群落结构特征、动态及其影响机制。对于理解该地区大型土壤动物区系组成及生态地理分布规律,及其在生态系统中的功能作用具有重要意义,并为合理利用和保护温带森林土壤生物资源及森林生态系统可持续发展提供理论依据。
1 研究区自然概况
研究样地设在黑龙江省尚志市东北林业大学帽儿山森林生态系统定位研究站(45°24′N,127°40′E)。该地区气候为温带大陆性季风气候,冬季寒冷而漫长,夏季湿热而短促,年平均温度2.7 ℃,全年大于或等于10 ℃的积温为2638 ℃;早霜一般出现在8月末,晚霜一般在5月末止,无霜期约为120—140 d;年降水量600—800 mm,雨量集中于7、8月份;年蒸发量1093 mm,年平均湿度70%;地带性土壤为暗棕壤。帽儿山位于东北东部山地张广才岭余脉,平均海拔300 m,平均坡度10—15°。植被属于长白植被区系,破坏后形成现今的次生林和人工林,代表着东北东部山区较典型的森林类型[18- 20]。本研究选取了该地区不同立地条件下经过不同干扰而形成的3种典型森林群落:红松人工林(湿度中等的山坡)、硬阔叶林(潮湿的山谷)和蒙古栎林(干旱的山脊)为研究对象,立地状况、植被组成及土壤理化性质见表1[18,20- 21]。
2 研究方法
2.1 野外采样与室内鉴定
于2012年5月到10月,每月对3种典型森林群落进行取样调查,在每种林型中各设置3块20 m×30 m样地,构成3个重复,按照多点混合取样法3点混合取样。分别对凋落物层、0—10 cm和10—20 cm共3个层次取样,取样面积为50 cm×50 cm。采用手拣法分离大型土壤动物,放入75%酒精中固定,带回实验室在显微镜下进行分类鉴定。主要参考尹文英的《中国土壤动物检索图鉴》[8],一般鉴定到科,个别鉴定到目或纲。统计土壤动物的类群和数量,同时将已鉴定的土壤动物洗净泥土, 用滤纸吸干其表面的酒精, 在分析天平上分别称量并记录鲜重。
2.2 数据分析
土壤动物的香农指数(H′)、均匀度指数(E)、优势度指数(C)和丰富度指数(D) 根据如下公式计算[22]:
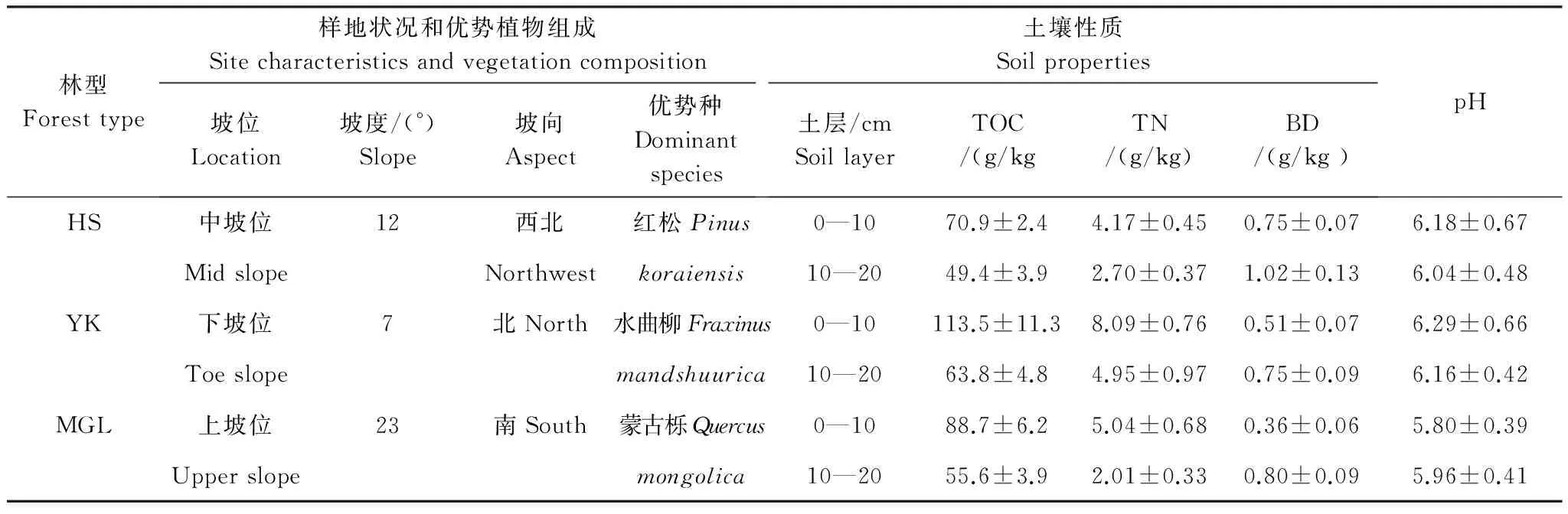
表1 3种森林群落的自然环境特征[18,20- 21]
HS:红松人工林Korean pine plantation forest;YK:硬阔叶林Hard-wood forest;MGL:蒙古栎林Mongolian oak forest;TOC:土壤总有机碳Total organic carbon;TN:全氮Total nitrogen;BD:容重Bulk Density
Shannon-Wiener多样性指数(H′)
H′=-∑PilnPi
(1)
Pielou均匀度指数(E)
E=H′/lnS
(2)
Simpson优势度指数(C)
C=∑(Pi)2
(3)
Menhinick丰富度指数(D)
D= lnS/lnN
(4)
式中,S为所有类群数;Pi为第i个种的多度比例;N为所有类群个体数。
采用方差分析(ANOVA,LSD)检验立地条件、土层深度及取样时间对3种群落大型土壤动物群落指标的显著性影响;双变量相关分析方法分析环境因子和土壤动物群落结构参数和多样性指数的相关性;采用典范对应分析(CCA)对优势类群和常见类群的数量与土壤环境因子进行分析。所有数据分析与作图采用SPSS19.0、Sigmaplot10.0及CANOCO 4.5软件。
3 研究结果
3.1 大型土壤动物的群落组成及数量特征
对帽儿山3种森林群落的6次土壤取样,共获得大型土壤动物59类、3604只,隶属于3门6纲17目50科,平均密度29.66只/m2(附表1)。优势类群(gt;总个体数10%)3类,分别为正蚓科(Lumbricidae)、线蚓科(Enchytraeidae)和石蜈蚣目(Lithobiomorpha),占总个体数的55.66%;常见类群(占总个体数1%—10%)11类,占总个体数的38.89%;稀有类群(lt;总个体数10%)45类,占总个体数的5.43%,它们虽然数量少,但是类群较多(附表1)。
3.2 大型土壤动物的水平分布
3个样地中,硬阔叶林大型土壤动物类群最多(8.59种/m2),但与红松人工林和蒙古栎林没有显著差异(表2)。不同样地共有类群21类,其中正蚓科为3个样地共有优势种,线蚓科和石蜈蚣目在3个样地所占比重均较大,但同一类群在不同样地多度有所不同(附表1)。此外,3个样地均有各自的特有类群,盲蝽科(Miridae)、水龟甲科(Hydrophilidae)和扁甲科(Cucujidae)等只出现在红松人工林;只出现在硬阔叶林的类群为猎蝽科(Reduviidae)、剑虻科(Therevidae)和舞虻科(Empididae)等;蒙古栎林的特有类群为毛蠓科(Psychodidae)、阎甲科(Histeridae)、和木蠹蛾科(Cossidae)等(附表1)。
不同样地大型土壤动物个体密度和生物量差异均不显著,其变化规律与土壤动物类群变化不同,均为红松人工林最大,蒙古栎林最小(表2)。不同植物群落中,一些类群个体密度差别不大,如地蜈蚣目(Geophilomorpha)(6.28—9.48只/m2),而一些类群在不同样地中个体密度差异较大(Pgt;0.05),如正蚓科在红松人工林个体密度达14.05只/m2,而在蒙古栎林个体密度仅4.84只/m2(附表1)。不同类群土壤动物的生物量差别悬殊,总平均生物量为27.88 g/m2;生物量最大的类群是正蚓科,其生物量占总生物量的78.92%;单只个体生物量最大的是木蠹蛾科,生物量为0.36 g,最小的为蓟马科(Thripidae),约0.0001 g。
大型土壤动物多样性指数分析表明,不同群落各多样性指数均无显著差异(表2),H′指数和D指数表现为蒙古栎林﹥硬阔叶林﹥红松人工林;C指数则正好相反,而E指数蒙古栎林最高(表2)。
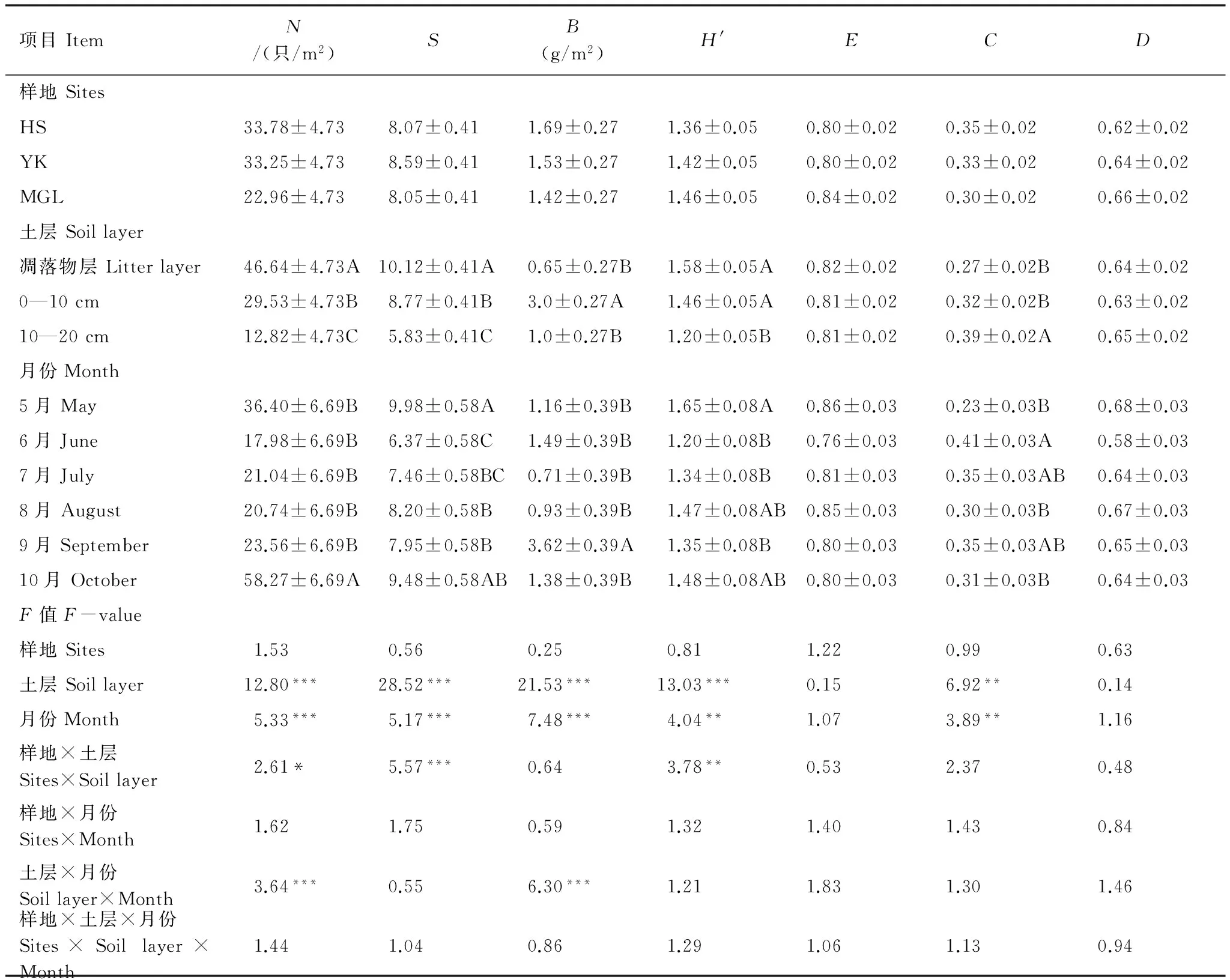
表2 大型土壤动物群落指标时空分布特征
同一列不同字母表示显著性差异组别(Plt;0.05),***Plt; 0.001,**Plt; 0.01,*Plt; 0.05;N:个体密度;S:类群数;B:生物量;H′:香农指数;E:均匀度指数;C:优势度指数;D:丰富度指数
3.3 大型土壤动物的垂直分布
大型土壤动物个体密度、类群数及生物量在垂直方向上的分布差异均极显著(Plt;0.001)(表2)。个体密度在土层中的分布具有明显表聚性,但各生境的变化情况有所不同(图1)。红松人工林中,大型土壤动物类群数沿凋落物层向下呈递减的趋势,而硬阔叶林和蒙古栎林0—10 cm土壤动物类群最多(图1)。
不同土层深度,土壤动物类群组成存在差异,如一些类群广泛分布在各个土层,如正蚓科和线蚓科在各层土壤均为优势种,蜘蛛目(Araneida)、马陆目(Juliformia)、隐翅甲科(Staphylinidae)和步甲科(Carabidae)在各层均为常见种;一些类群则集中分布于特定土层,如蝽科(Pentatomidae)、郭公甲科(Cleridae)和胡蜂科(Vespidae)仅出现在凋落物层,而扁甲科、虎甲科(Cicindelidae)和蓟马科等仅出现在0—10 cm土层。红松人工林凋落物层个体密度显著高于其它两层,而硬阔叶林和蒙古栎林凋落物层与0—10 cm土层无显著差异(图1)。不同类群土壤动物个体密度在垂直方向上变化规律也不尽相同,金龟甲科(Scarabaeidae)随深度增加而增加,步甲科、腹足纲(Gastropoda)和蚋科(Siliidae)等随深度增加而减小,而长足虻科(Dolichopodidae)在各层的密度大致相当。
0—10 cm土层大型土壤动物生物量显著高于其它两层(表2),蒙古栎林和硬阔叶林10—20 cm土层大型土壤动物生物量大于凋落物层,红松人工林则正好相反,但均无显著差异(图1)。
不同深度土层大型土壤动物的H′指数和C指数差异显著(Plt;0.01),H′指数随深度增加而减小,C指数则正好相反(表2)。E指数和D指数则无显著差异(表2)。
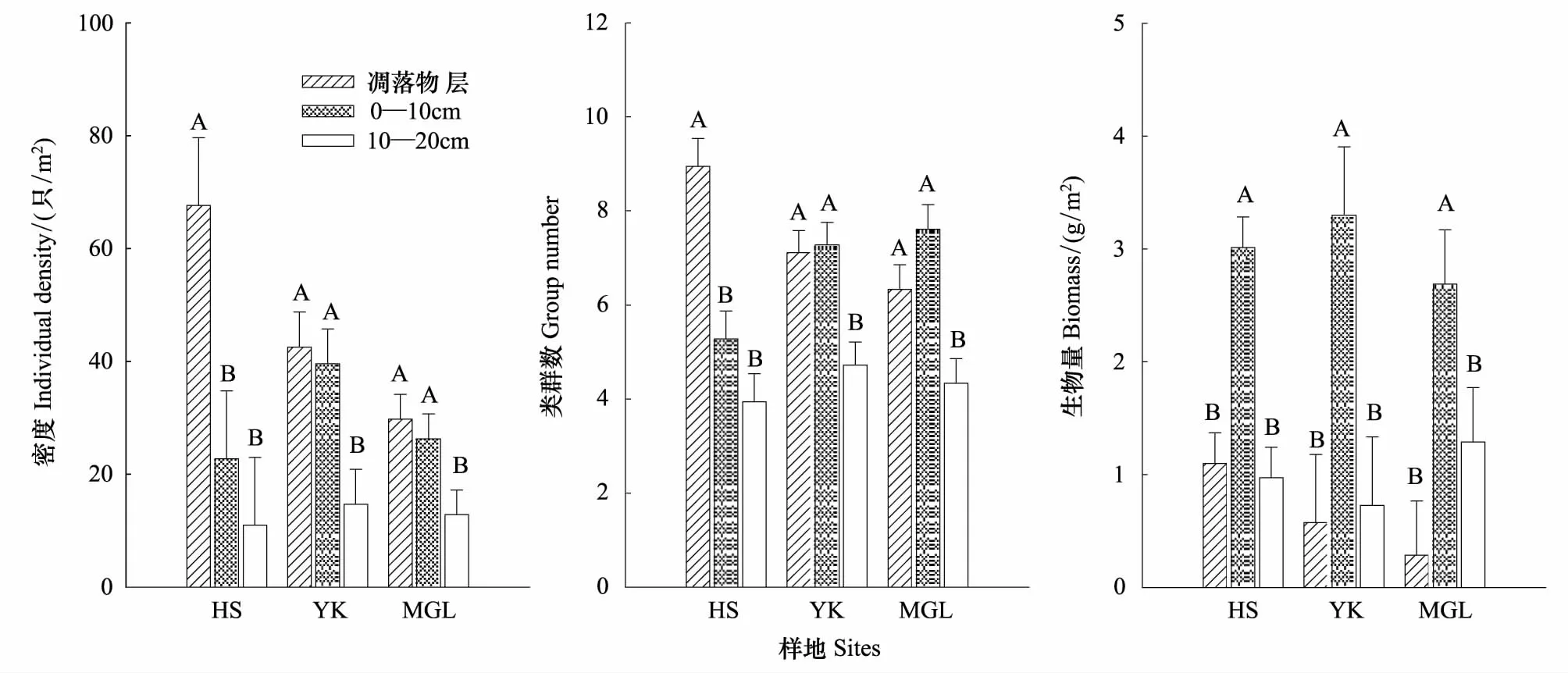
图1 不同样地大型土壤动物个体密度、类群数和生物量的垂直分布Fig.1 The vertical distribution of individual density, group number and biomass of soil macro-fauna in different sitesHS:红松人工林Korean pine plantation forest;YK:硬阔叶林Hard-wood forest;MGL:蒙古栎林Mongolian oak forest;TOC:土壤总有机碳Total organic carbon;TN:全氮Total nitrogen;BD:容重Bulk Density; A,B表示差异显著(Plt;0.05)
3.4 大型土壤动物的时间分布规律
大型土壤动物类群数、个体密度及生物量均随时间变化显著(Plt;0.001)(表2)。总体看来,大型土壤动物密度和类群数均以5月和10月较多(表2),但不同样地大型土壤动物个体密度和类群数时间变化规律存在差异,密度和类群数在红松人工林和硬阔叶林随时间变化显著,而其在蒙古栎林无显著差异(图2)。不同时间大型土壤动物的组成也不同,有些类群只出现在特定的月份,如蓟马科只出现在5月,剑虻科仅在10月出现等。
3种群落大型土壤动物9月生物量显著高于其它各月(图2)。H′指数和C指数时间变化差异极显著(Plt;0.01),H′指数最大值出现在5月,C指数则6月高于其他各月。其他指数各月份间无显著差异(表2)。
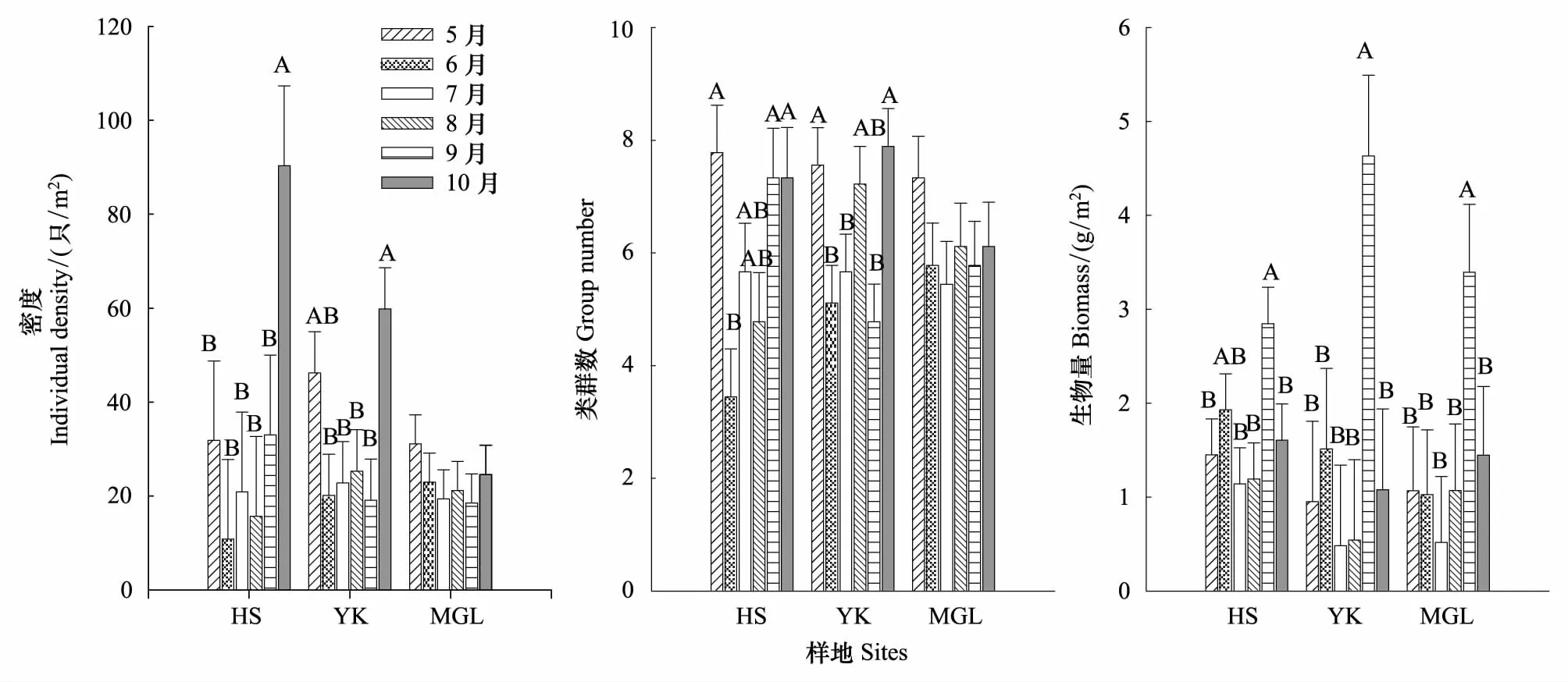
图2 大型土壤动物个体密度、类群数和生物量的时间动态Fig.2 The dynamics of individual density, group number and biomass of soil macro-fauna in different sitesA,B表示差异显著(Plt;0.05)
3.5 大型土壤动物群落指标与土壤环境因子的相关分析
双变量相关分析结果表明:大型土壤动物类群数呈现与总有机碳正相关,而与容重呈显著负相关关系且相关性极显著(Plt;0.01);个体密度与总有机碳和全氮均显著正相关;生物量则与总有机碳显著正相关;H′指数与容重极显著负相关,C指数则与容重极显著正相关(Plt;0.01)(表3)。
利用CCA分析大型土壤动物优势类群和常见类群的数量与土壤理化性质之间的关系(图3)。图中第一轴与第二轴的Eigenvalues值分别为0.079和0.017。总体而言,石蜈蚣目、蜘蛛目和马陆目等分布广泛,主要集中于图的中心位置;红蝽科(Pyrrhocoridae)与全氮的相关性较大,腹足纲与总有机碳和全氮的相关性较大,而与容重相关性较大的为蚋科和隐翅甲科(图3)。
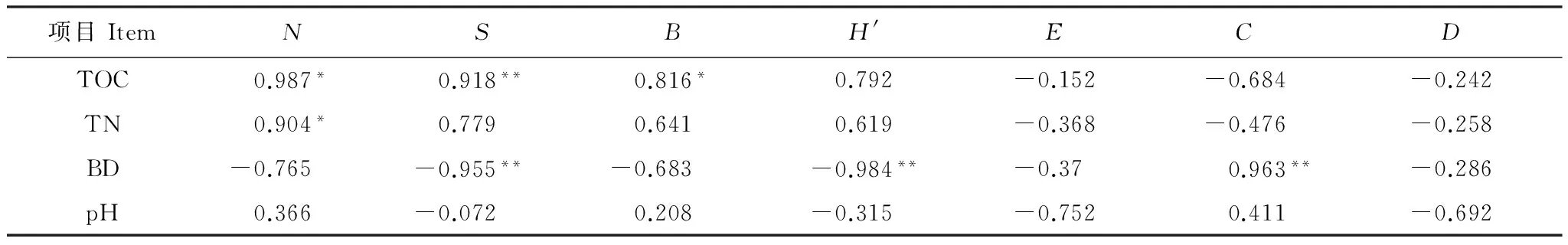
表3 大型土壤动物群落指标与土壤环境因子的相关性
* 在 0.05 水平(双侧)上显著相关;** 在0.01 水平(双侧)上显著相关
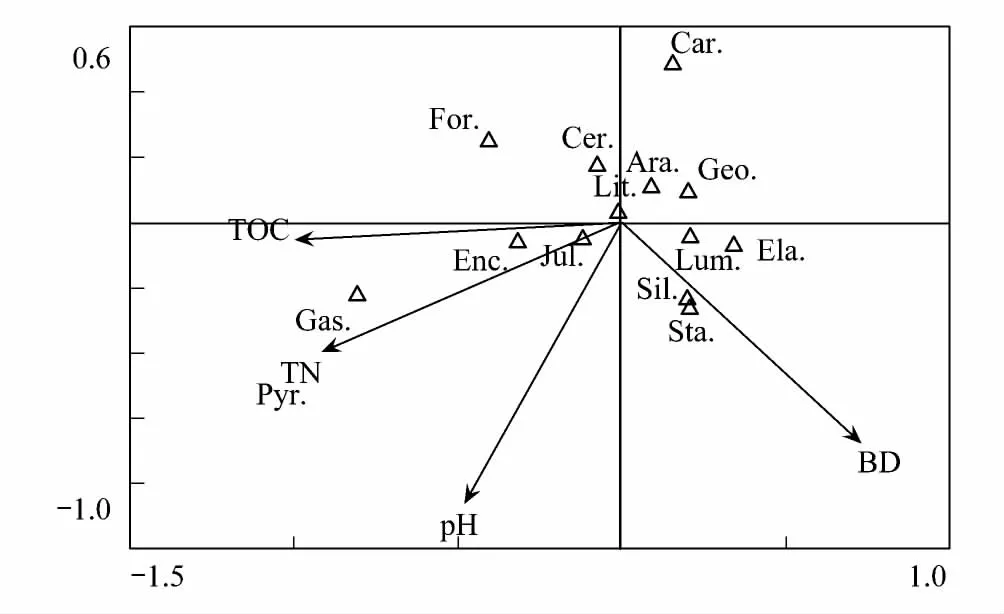
图3 大型土壤动物典范对应分析二维排序图 Fig.3 Canonical correspondence analysis ordination based on soil macro-faunaLum.:正蚓科Lumbricidae;Enc.:线蚓科Enchytraeidae;Lit.:石蜈蚣目Lithobiomorpha;For.:蚁科Formicidae;Geo.:地蜈蚣目Geophilomorpha;Ara.:蜘蛛目Araneida;Jul.:马陆目Juliformia;Sta.:隐翅甲科Staphylinidae;Gas.:腹足纲Gastropoda;Cer.:蠓科Ceratopogonidae;Ela. :叩甲科Elateridae;Car.:步甲科Carabidae;Pyr.:红蝽科Pyrrhocoridae;Sil.:蚋科Siliidae
4 讨论
4.1 大型土壤动物群落组成特征
通过对帽儿山3种温带森林群落大型土壤动物的调查表明,正蚓科、线蚓科和石蜈蚣目为优势类群,表现出对环境普适性。许振文等[23]在长白山地区的研究表明线蚓科为优势类群,而正蚓科为常见类群,与本研究结论不完全一致,说明不同地区温带森林土壤动物的类群组成存在差别。稀有类群对环境因子耐受较小,由于环境的异质性使其分布于不同的地区,因此3种森林群落以及不同土层深度均有各自的特有类群,这与以往学者[14]对帽儿山土壤动物群落结构研究结果基本一致,即稀有类群虽然数量少, 但是生态适应性弱,只有在一定条件下才能生存在个别群落中,因此能够反映环境的差异,对环境变化具有一定的指示作用[24]。由此可见,对于土壤动物的研究应该更加注重稀有类群的分析,基于物种水平的土壤动物稀有类群与环境关系的研究,能更好的对环境变化及发展趋势进行评价和预测。
4.2 大型土壤动物空间分布特征
不同群落大型土壤动物类群数、个体密度及生物量差异不显著,主要是因为土壤动物优势类群和常见类群多具有较强的环境适应性(图3),由此使温带森林生态系统大型土壤动物群落存在较大的稳定性,群落结构不随环境差异而显著变化。但微生境的变化也会引起土壤动物群落结构的细微差异,土壤环境因子是引起土壤动物个体密度和类群数变化的主要原因[25]。蒙古栎林坡向为南坡,在温带地区南坡一般为阳坡,林冠郁闭度小,热量条件较好,坡度在3个样地中最大,土壤最干燥,不利于喜湿的土壤动物生存,因此正蚓科和线蚓科密度最小,同时这两种类群均为土壤动物群落的优势类群,从而导致蒙古栎林大型土壤动物总密度小于其它两个森林群落。红松人工林与硬阔叶林的立地条件相似,坡向均为北坡,硬阔叶林树种密度较红松林低,凋落物层比较薄,因此土壤动物个体密度略少于红松人工林。在大型土壤动物各类群中,正蚓科生物量最大,占总生物量78.92%。生物量是能量研究中一个最基本的参数[26],生物量较大的类群能迅速改良或破坏土壤的物理结构,影响土壤食物网,从而影响营养元素在生态系统中的周转率[27],此结果表明正蚓科是帽儿山森林生态系统重要的分解者,在能量流动中具有重要作用,这与以往研究结果一致[28]。
相关分析表明容重与H′指数显著负相关,与C指数显著正相关,蒙古栎林容重显著低于其它两个样地,尤其是0—10 cm土层(表1),因此3个样地中蒙古栎林C指数最低,H′指数最高。土壤动物多样性反映土壤中食物资源和生境的多样性、环境压力强度和土壤动物对环境的耐力,多样性指数比类群和密度更能体现群落的结构,在理论和实践中都具有重要意义。
凋落物及其形成的腐殖质是土壤动物重要食物来源,且具有一定保温和保湿作用,有利于土壤动物群落的数量增长[29],随深度增加,土壤有机碳、全氮及养分下降(表1),且通气透水性及温度随之降低[29]。因此,在垂直方向上,大型土壤动物个体密度及类群数有明显表聚性特征,而蒙古栎林内干燥,凋落物不易分解,凋落物层湿度小,有机质含量低,0—10 cm土层养分充足,根系发达,通气良好,所以在蒙古栎林大型土壤动物类群数以0—10 cm土层最丰富。土壤动物垂直分布特征说明,维持土壤表层的质量在保护土壤动物种群方面具有重要意义。
4.3 大型土壤动物时间分布特征
温带地区环境条件(如光照、气温、降水、食物)的时间变化,既影响土壤动物的群落结构和多样性,又促使土壤动物形成适应机制。本研究结果显示大型土壤动物密度和类群数5月和10月较多,这与林英华[14]的结论是相似的。帽儿山位于温带地区,生物有明显的年周期节律,5月气温回升、积雪融水使土壤的温度和水分增加,永久栖居的土壤动物复苏,暂时栖居的土壤动物以幼虫形式存在土壤中;10月凋落物层达到最厚,气温和水分条件适宜,大气中的部分昆虫进入土壤中冬眠。因此,这两段时间土壤动物的类群、密度和H′指数最大。7月和8月土壤动物密度并未达到最高可能是由于夏季降水集中,土壤水分处于饱和状态,造成一些土壤动物直接淹没死亡或因土壤空气缺氧窒息死亡等。而大型土壤动物生物量则为9月最高,此时经过生长季的积累,大型土壤动物接近成熟、个体较大,随后有些成虫完成繁殖后死亡,生物量减少。生物量的时间变化反映各月份土壤动物对环境作用强度的变化[30- 31]。蒙古栎林的个体密度和类群数各月差异不显著,说明其环境因子随时间变化较其余两个样地稳定。可见,大型土壤动物的类群数、密度、生物量及多样性指数动态变化是食物(凋落物数量和分解程度)和温湿综合作用的结果,土壤动物群落指标可指示森林生态系统对全球气候变化的响应。
4.4 大型土壤动物与土壤理化性质相关分析
双变量相关分析结果表明:大型土壤动物密度、类群数和生物量与总有机碳极显著正相关,密度与全氮显著正相关,这与刘继亮等[11]对左家自然保护区大型土壤动物与环境因子的关系研究结果一致。因此,在土壤环境各因子中,土壤有机质和全氮对大型土壤动物的分布影响较大。CCA结果表明,不同类群土壤动物与土壤理化环境的相关性不同:红蝽科与全氮显著相关,腹足纲与总有机碳和全氮显著正相关,而蚋科和隐翅甲科与土壤容重显著正相关。不同类群的土壤动物具有选择性吸收和富集某些元素的功能[32],因此研究土壤动物分布与生态环境的关系,对促进土壤动物的有益发展和抑制其有害活动,维持土壤生态系统平衡,具有重要意义。
5 结论
通过对帽儿山3种森林群落大型土壤动物连续动态研究发现,土壤动物的群落结构和多样性受植被、立地条件和土壤理化性质的影响。不同样地间大型土壤动物群落结构指标差异不显著,说明多数类群具有较大的环境适应性;土壤动物密度、类群数和多样性指数垂直分布基本表现出表聚性,即从凋落物层向下显著减小;时间变化表现为5月和10月个体密度较大,类群较丰富,9月生物量最大。由此可见,土壤微环境要素的特征及其时间和空间的动态变化规律直接决定了土壤动物的生活环境,大型土壤动物的群落结构及多样性等具有较大的时空异质性。
致谢:帽儿山森林生态定位站在实验过程中给予帮助,特此致谢。
[1] Sun R Y. Principles of animal ecology. Beijing: Beijing Normal University Press, 2001.
[2] Edwards C A. Assessing the effects of environmental pollutants on soil organisms, communities, processes and ecosystems. European Journal of Soil Biology, 2002, 38(3/4): 225- 231.
[3] Freckman D W, Ettema C H. Assessing nematode communities in agroecosystems of varying human intervention. Agriculture, Ecosystems and Environment, 1993, 45(3/4): 239- 261.
[4] Fu S L, Ferris H, Brown D, Plant R. Does the positive feedback effect of nematodes on the biomass and activity of their bacteria prey vary with nematode species and population size? Soil Biology and Biochemistry, 2005, 37(11): 1979- 1987.
[5] Wang S J, Ruan H H. Feedback mechanisms of soil biota to aboveground biology in terrestrial ecosystems. Biodiversity Science, 2008, 16(4): 407- 416.
[6] Li T, Liu W Q, Guo S M, Ke G Q, Zhang Z, Xiao X B, Liu W. Community traits of soil fauna in forestlands converted from cultivated lands in limestone red soil region of Ruichang, Jiangxi Province of China. Chinese Journal of Applied Ecology, 2012, 23(4): 910- 916.
[7] Kardol P, Reynolds W N, Norby R J, Classen A T. Climate change effects on soil microarthropod abundance and community structure. Applied Soil Ecology, 2011, 47(1): 37- 44.
[8] Yin W Y. Pictorial Keys to Soil Animals of China. Beijing: Science Press, 1998.
[9] Liao C H, Li J X, Yang Y P, Zhang Z C. The community of soil animal in tropical rain forest in Jianfeng Mountain, Hainan Island, China: Relationship between seasonal change of community structure and climatic factors. Acta Ecologica Sinica, 2003, 23(1): 139- 147.
[10] Wolters V. Biodiversity of soil animals and its function. European Journal of Soil Biology, 2001, 37(4): 221- 227.
[11] Liu J L, Yin X Q, Qiu L L. Large-sized soil fauna and soil factors in Zuojia Nature Reserve. Acta Pedologica Sinica, 2008, 45(1): 130- 136.
[12] Jiao X L, Zhu J J, Yan Q L. Composition and seasonal dynamics of macro- and meso- soil fauna in secondary forest ecosystems of Liaodong montane area, China. Acta Ecologica Sinica, 2009, 29(5): 2631- 2638.
[13] Pang X Y, Liu Q, Liu S Q, Wu Y, Lin B, He H. Effect of human-induced disturbance on soil physical properties of subalpine coniferous forests in western Sichuan. Chinese Journal of Applied and Environmental Biology, 2002, 8(6): 583- 587.
[14] Lin Y H, Sun J B, Liu H L, Zhang F D, Sun L, Jin S. Composition of soil fauna community and its diversity analysis at Maoershan Mountains of Heilongjiang Province. Scientia Silvae Sinicae, 2006, 42(4): 71- 77.
[15] Liu J L, Cao J, Li S J, Pan C L, Pan C C. Characteristics of soil macrofaunal community structure in secondary forest and forest plantations in western Qinling Mountains of Northwest China. Chinese Journal of Applied Ecology, 2012, 23(9): 2459- 2466.
[16] Zhang X P, Huang C L, Li J K. Efficiencies of earthworms (Eiseniafoetida) decomposing forest litters. Acta Ecologica Sinica, 2005, 25(9): 2427- 2433.
[17] Zhong W Y, Yin X Q, Chen P. Relationship of litter decomposition and consumption with soil animals in Maoer Mountain forest. Chinese Journal of Applied Ecology, 1999, 10(4): 511- 512.
[18] Guo Q X, Zhang H Y, Wang X C, Wang C K. Simulation of soil respiration for typical forests in Northeast China: Localized application of IBIS. Acta Ecologica Sinica, 2010, 30(9): 2295- 2303.
[19] Yang J Y, Wang C K. Partitioning soil respitation of temperate forest ecosystems in Northeastern China. Acta Ecologica Sinica, 2006, 26(6): 1640- 1647.
[20] Yang J Y, Wang C K. Soil carbon storage and flux of temperate forest ecosystems in northeastern China. Acta Ecologica Sinica, 2005, 25(11): 2875- 2882.
[21] Liu S, Wang C K. Spatio-temporal patterns of soil microbial biomass carbon and nitrogen in five temperate forest ecosystems. Acta Ecologica Sinica, 2010, 30(12): 3135- 3143.
[22] Zhang X P, Huang L R, Jiang L Q. Characteristics of macro-soil fauna in forest ecosystem of northern Da Hinggan Mountains. Geographical Research, 2008, 27(3): 509- 518.
[23] Xu Z W, Liu G, Zuo W. Soil fauna in Tumenling of Changbaishan Mountain. Soil, 2003, (2): 152- 159.
[24] Yin X Q, Wu D H, Han X M. Diversity of soil animals community in Xiao Hinggan Mountains. Scientia Geographica Sinica, 2003, 23(3): 316- 322.
[25] Liu R T, Zhao H L, Zhao X Y. Effects of different afforestation types on soil faunal diversity in Horqin Sand Land. Chinese Journal of Applied Ecology, 2012, 23(4): 1104- 1110.
[26] Yan D K, Wang S L, Hu Y L, Gao H, Zhang X Y. A comparative study on soil fauna in native secondary evergreen broad-leaved forest and Chinese fir plantation forests in subtropics. Chinese Journal of Applied Ecology, 2004, 15(10): 1792- 1796.
[27] Yin X Q, Liu J L, Gao M. Relationships of nutrient elements in forest ecosysterm of Xiao Hinggan Mountains and role of soil fauna. Scientia Geographica Sinica, 2007, 27(6): 814- 819.
[28] Zhang X P, Hou W L, Chen P. Soil animal guilds and their ecological distribution in the northeast of China. Chinese Journal of Applied and Environmental Biology, 2001, 7(4): 370- 374.
[29] Cui L W, Liu S R, Liu X L, Wu P F, Zhang H Z, He X J. Changes of soil faunal communities during the restoration progress ofAbiesfaxonianaForests in Northwestern Sichuan. Acta Ecologica Sinica, 2012, 32(15): 4772- 4783.
[30] Guan H B, Guo L, Liu Y J. The vertical distribution, seasonal dynamics and community variety of soil animal in Horqin Sandy Land. Journal of Desert Research, 1999, 19(1): 110- 114.
[31] Liu Y J, Liu X M, Guo L, A L T Q Q G. The study on soil animal ecology in Inner Mongolia grassland. Grassland of China, 1999, (3): 51- 56.
[32] Chen P, Fu D Y. A preliminary approach to the effect of soil animals on the cycling of materials in the Changbai Shan Mountain. Acta Ecologica Sinica, 1984, 4(2): 172- 180.
参考文献:
[1] 孙儒泳. 动物生态学原理. 北京: 北京师范大学出版社, 2001.
[5] 王邵军, 阮宏华. 土壤生物对地上生物的反馈作用及其机制. 生物多样性, 2008, 16(4): 407- 416.
[6] 李涛, 刘苑秋, 郭圣茂, 柯国庆, 张钊, 肖绪宝, 刘武. 瑞昌石灰岩红壤区退耕还林土壤动物群落特征. 应用生态学报, 2012, 23(4): 910- 916.
[8] 尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
[9] 廖崇惠, 李健雄, 杨悦屏, 张振才. 海南尖峰岭热带林土壤动物群落——群落结构的季节变化及其气候因素. 生态学报, 2003, 23(1): 139- 147.
[11] 刘继亮, 殷秀琴, 邱丽丽. 左家自然保护区大型土壤动物与土壤因子关系研究. 土壤学报, 2008, 45(1): 130- 136.
[12] 焦向丽, 朱教君, 闫巧玲. 辽东山区次生林生态系统大、中型土壤动物组成与季节动态. 生态学报, 2009, 29(5): 2631- 2638.
[13] 庞学勇, 刘庆, 刘世全, 吴彦, 林波, 何海. 人为干扰对川西亚高山针叶林土壤物理性质的影响. 应用与环境生物学报, 2002, 8(6): 583- 587.
[14] 林英华, 孙家宝, 刘海良, 张夫道, 孙龙, 金森. 黑龙江帽儿山土壤动物群落组成与多样性分析. 林业科学, 2006, 42(4): 71- 77.
[15] 刘继亮, 曹靖, 李世杰, 潘春林, 潘成臣. 秦岭西部山地次生林和人工林大型土壤动物群落结构特征. 应用生态学报, 2012, 23(9): 2459- 2466.
[16] 张雪萍, 黄初龙, 李景科. 赤子爱胜蚓对森林凋落物的分解效率. 生态学报, 2005, 25(9): 2427- 2433.
[17] 仲伟彦, 殷秀琴, 陈鹏. 帽儿山森林落叶分解消耗与土壤动物关系的研究. 应用生态学报, 1999, 10(4): 511- 512.
[18] 国庆喜, 张海燕, 王兴昌, 王传宽. 东北典型森林土壤呼吸的模拟- IBIS模型的局域化应用. 生态学报, 2010, 30(9): 2295- 2303.
[19] 杨金艳, 王传宽. 东北东部森林生态系统土壤呼吸组分的分离量化. 生态学报, 2006, 26(6): 1640- 1647.
[20] 杨金艳, 王传宽. 东北东部森林生态系统土壤碳贮量和碳通量. 生态学报, 2005, 25(11): 2875- 2882.
[21] 刘爽, 王传宽. 五种温带森林土壤微生物生物量碳氮的时空格局. 生态学报, 2010, 30(12): 3135- 3143.
[22] 张雪萍, 黄丽荣, 姜丽秋. 大兴安岭北部森林生态系统大型土壤动物群落特征. 地理研究, 2008, 27(3): 509- 518.
[23] 许振文, 刘刚, 左伟. 长白山余脉土们岭土壤动物调查与分析. 土壤, 2003, (2): 156- 159.
[24] 殷秀琴, 吴东辉, 韩晓梅. 小兴安岭森林土壤动物群落多样性的研究. 地理科学, 2003, 23(3): 316- 322.
[25] 刘任涛, 赵哈林, 赵学勇. 科尔沁沙地不同造林类型对土壤动物多样性的影响. 应用生态学报, 2012, 23(4): 1104- 1110.
[26] 颜绍馗, 汪思龙, 胡亚林, 高洪, 张秀永. 亚热带天然次生常绿阔叶林与杉木人工林土壤动物群落特征比较. 应用生态学报, 2004, 15(10): 1792- 1796.
[27] 殷秀琴, 刘继亮, 高明. 小兴安岭森林生态系统中营养元素关系及土壤动物的作用. 地理科学, 2007, 27(6): 814- 819.
[28] 张雪萍, 侯威岭, 陈鹏. 东北森林土壤动物同功能种团及其生态分布. 应用与环境生物学报, 2001, 7(4): 370- 374.
[29] 崔丽巍, 刘世荣, 刘兴良, 吴鹏飞, 张洪芝, 何先进. 川西北冷杉林恢复过程中土壤动物群落动态. 生态学报, 2012, 32(15): 4772- 4783.
[30] 关宏斌, 郭砺, 刘永江. 科尔沁沙地土壤动物垂直分布、季节动态及群落多样性探讨. 中国沙漠, 1999, 19(1): 110- 114.
[31] 刘永江, 刘新民, 郭砺, 阿拉坦其其格. 内蒙古草原带土壤动物生态学研究. 中国草地, 1999, (3): 51- 56.
[32] 陈鹏, 富德义. 长白山土壤动物在物质循环中作用的初步探讨. 生态学报, 1984, 4(2): 172- 180.
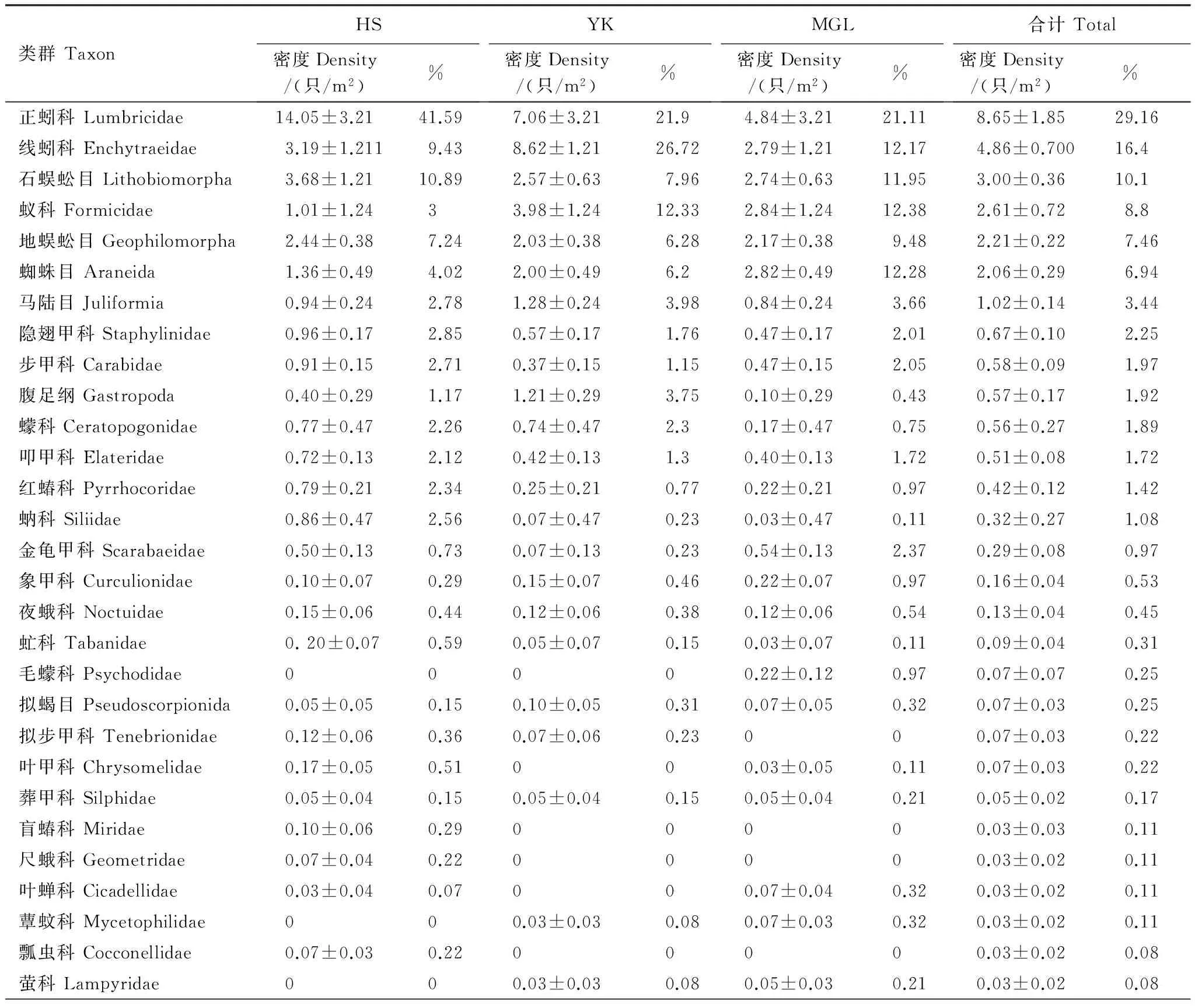
附表1 不同森林群落大型土壤动物类群和数量组成
续表
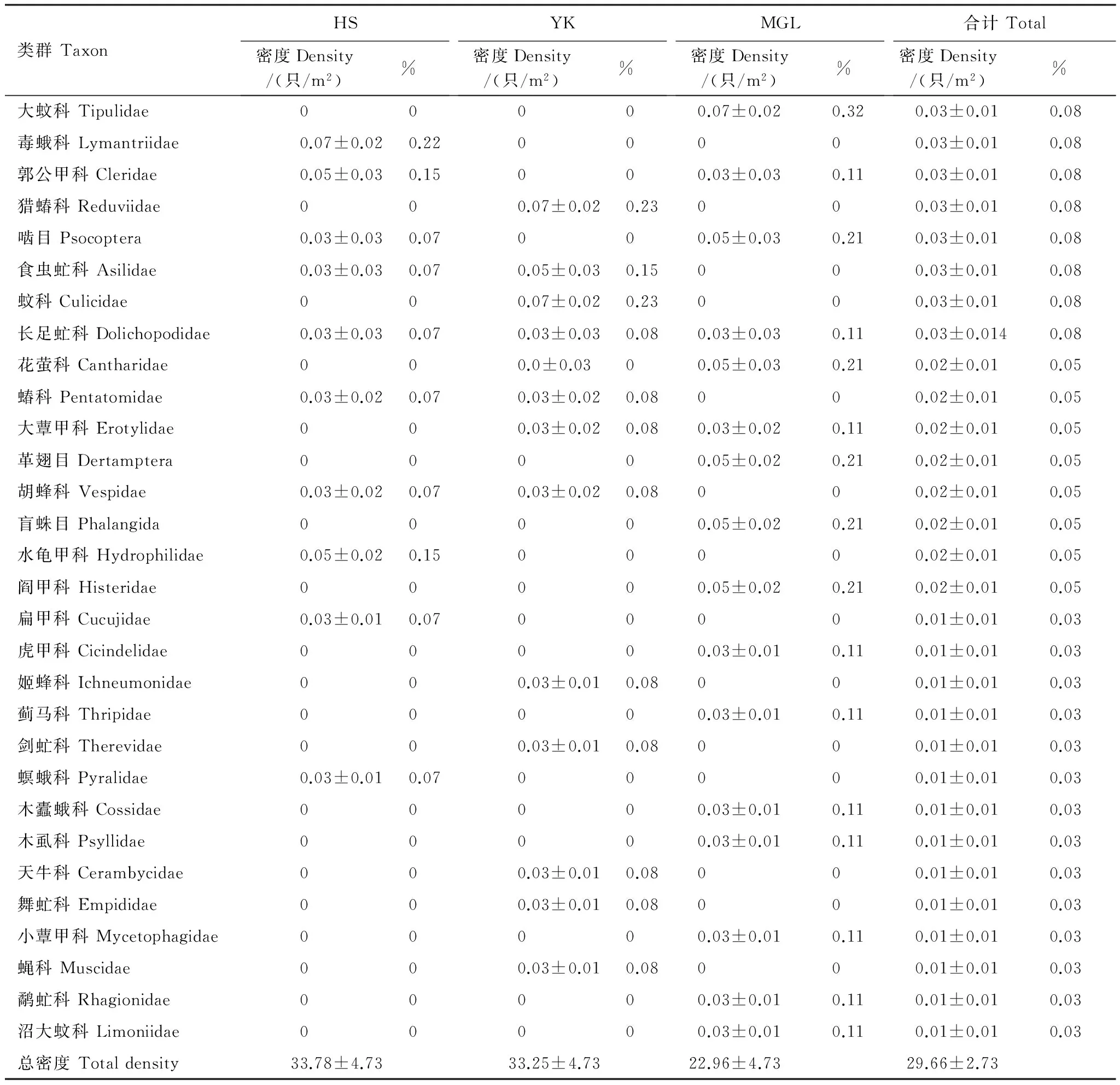
类群TaxonHS密度Density/(只/m2)%YK密度Density/(只/m2)%MGL密度Density/(只/m2)%合计Total密度Density/(只/m2)%大蚊科Tipulidae00000.07±0.020.320.03±0.010.08毒蛾科Lymantriidae0.07±0.020.2200000.03±0.010.08郭公甲科Cleridae0.05±0.030.15000.03±0.030.110.03±0.010.08猎蝽科Reduviidae000.07±0.020.23000.03±0.010.08啮目Psocoptera0.03±0.030.07000.05±0.030.210.03±0.010.08食虫虻科Asilidae0.03±0.030.070.05±0.030.15000.03±0.010.08蚊科Culicidae000.07±0.020.23000.03±0.010.08长足虻科Dolichopodidae0.03±0.030.070.03±0.030.080.03±0.030.110.03±0.0140.08花萤科Cantharidae000.0±0.0300.05±0.030.210.02±0.010.05蝽科Pentatomidae0.03±0.020.070.03±0.020.08000.02±0.010.05大蕈甲科Erotylidae000.03±0.020.080.03±0.020.110.02±0.010.05革翅目Dertamptera00000.05±0.020.210.02±0.010.05胡蜂科Vespidae0.03±0.020.070.03±0.020.08000.02±0.010.05盲蛛目Phalangida00000.05±0.020.210.02±0.010.05水龟甲科Hydrophilidae0.05±0.020.1500000.02±0.010.05阎甲科Histeridae00000.05±0.020.210.02±0.010.05扁甲科Cucujidae0.03±0.010.0700000.01±0.010.03虎甲科Cicindelidae00000.03±0.010.110.01±0.010.03姬蜂科Ichneumonidae000.03±0.010.08000.01±0.010.03蓟马科Thripidae00000.03±0.010.110.01±0.010.03剑虻科Therevidae000.03±0.010.08000.01±0.010.03螟蛾科Pyralidae0.03±0.010.0700000.01±0.010.03木蠹蛾科Cossidae00000.03±0.010.110.01±0.010.03木虱科Psyllidae00000.03±0.010.110.01±0.010.03天牛科Cerambycidae000.03±0.010.08000.01±0.010.03舞虻科Empididae000.03±0.010.08000.01±0.010.03小蕈甲科Mycetophagidae00000.03±0.010.110.01±0.010.03蝇科Muscidae000.03±0.010.08000.01±0.010.03鹬虻科Rhagionidae00000.03±0.010.110.01±0.010.03沼大蚊科Limoniidae00000.03±0.010.110.01±0.010.03总密度Totaldensity33.78±4.7333.25±4.7322.96±4.7329.66±2.73
Spatialandtemporalvariationofsoilmacro-faunacommunitystructureinthreetemperateforests
LI Na, ZHANG Xueping, ZHANG Limin*
KeyLaboratoryofRemoteSensingMonitoringofGeographicEnvironment,CollegeofGeographicalScience,HarbinNormalUniversity,CollegeofHeilongjiangProvince,Harbin150025,China
Soil macro-faunas are important decomposers in ecosystem and play a significant role in nutrient cycling and energy flow. In Maoer Mountain, the soil macro-faunas were investigated in three typical forest types (Korean pine plantation forest, Hard-wood forest and Mongolian oak forest). Our object was to explore the community structure, spatial and temporal dynamic of soil macro-fauna, and its impact factors in temperate forest. Soil samples were collected from the litter layer and the soil layers at depth of 0—10 cm and 10—20 cm. Sampling was carried out continuously from May to October in 2012. A total of 3604 individuals, belonging to 3 Phyla, 6 Classes, 17 Orders and 50 families were extracted and identified. Among them, the Lumbricidae, Enchytraeidae and Lithobiomorpha were the dominant communities. The results showed that: (1)for horizontal distribution, the decreasing order of individual density and biomass was Korean pine plantation, Hard-wood forest and Mongolian oak. The group number of soil macro-faunas was the largest in Hard-wood forest, and the least in Mongolian oak forest. The Shannon and richness indices of soil macro-faunas were the highest in Mongolian oak forest and the lowest in Korean pine plantation forest, but the trend in the dominance index was opposite. The evenness index was the largest in Mongolian oak forest and the least in Hard-wood forest; (2) for vertical distribution, the individual density, group number, biomass and diversity of soil macro-fauna were significantly affected by the soil layer (Plt;0.001). The individual density distribution appeared significant surface-aggregation, and the maximum biomass of soil macro-fauna was found at depth of 0—10 cm. The group number showed a decrease trend with the increasing layer in Korean pine plantation forest and that of highest value in both Hard-wood forest and Mongolian oak forest occurred at 0—10 cm depth, not the litter layer. The Shannon index decreased with the increasing soil depth, while the dominance index appeared the opposite trend; (3) the Shannon and dominance index of soil macro-fauna were significantly different among different months(Plt;0.01). Individual density and group number were higher in May and September than other months, and biomass of soil macro-fauna was the largest in September; (4) the correlation analysis showed that the individual density, group number and biomass of the soil macro-fauna were positively correlated with total organic carbon (Plt;0.05), there was the significantly negative correlation between the Shannon index and soil bulk density (Plt;0.01). It can be conclude that there was obviously spatial and temporal heterogeneity in soil macro-fauna community structure and the total organic carbon was one of the important impact factors.
soil macro-faunas; forest ecosystem; community structure; diversity; spatial and temporal distribution
国家科技部“十二五”科技支撑项目(2011BAD37B01);国家自然科学基金项目(41101048);国家自然科学基金项目(41071033)
2013- 05- 24;
2013- 07- 29
*通讯作者Corresponding author.E-mail: zlmjhb@163.com
10.5846/stxb201305241166
李娜, 张雪萍, 张利敏.三种温带森林大型土壤动物群落结构的时空动态.生态学报,2013,33(19):6236- 6245.
Li N, Zhang X P, Zhang L M.Spatial and temporal variation of soil macro-fauna community structure in three temperate forests .Acta Ecologica Sinica,2013,33(19):6236- 6245.
