利用红外照相技术分析野生白冠长尾雉活动节律及时间分配
2013-12-09赵玉泽王志臣徐基良安丽丹
赵玉泽,王志臣,徐基良,*,罗 旭,安丽丹
(1. 北京林业大学自然保护区学院,北京 100083; 2. 国家林业局调查规划设计院,北京 100714;3. 国家林业局野生动植物保护与自然保护区管理司,北京 100714)
利用红外照相技术分析野生白冠长尾雉活动节律及时间分配
赵玉泽1,王志臣2,徐基良1,*,罗 旭1,安丽丹3
(1. 北京林业大学自然保护区学院,北京 100083; 2. 国家林业局调查规划设计院,北京 100714;3. 国家林业局野生动植物保护与自然保护区管理司,北京 100714)
2012年3月—2013年3月,利用红外相机技术在湖北省广水市蔡河镇对野生白冠长尾雉的活动节律和时间分配进行了研究。利用16台红外相机在40个相机位点对白冠长尾雉进行了监测。累计1774个相机日,拍摄到2242个独立视频,其中白冠长尾雉的视频占18%,共记录538只次,雌雄比为1.43∶1。结果表明,白冠长尾雉每日有两个活动高峰期,上午雄性个体的活动高峰期比雌性个体早两个小时。白冠长尾雉的主要行为是移动和觅食,分别占到总频次的40.71%和33.10%,其余5种行为依次为: 警戒9.29%,梳理7.14%,休息5.00%,对抗2.62%,育幼2.14%。雌性白冠长尾雉的警戒行为频次比例显著高于雄性个体(Plt;0.05)。不同季节之间取食行为、移动行为、对抗行为比例之间有显著差异,冬季的取食行为比例明显高于夏季(Plt;0.05),夏季移动行为比例显著高于秋季(Plt;0.05)和冬季(Plt;0.05),而警戒、梳理、休息和育幼行为比例则无显著差异。
红外相机技术;白冠长尾雉;活动节律;时间分配
活动节律与时间分配是动物行为学研究的重要内容[1],其结果可以反映出动物个体的营养状态、社会地位等状况,也有助于分析物种的生存状况或对策,对于制定保护措施具有十分重要的意义[2]。虽然瞬时扫描法和焦点动物取样法[2- 4]比较适用于大型哺乳动物或者是笼养野生动物的观察研究,但并不适合对人类活动敏感或数量稀少的濒危物种。红外线触发自动数码相机陷阱技术(以下简称红外相机技术)具有长期性、客观性、隐蔽性、无损伤性等特征[5- 6],在解决这一问题中具有天然优势。因此,其逐渐广泛应用于野生动物相关研究中,且在国外已经有很长的历史[7- 10],如红外相机技术在鸟类巢穴捕食、营巢行为等方面已经有过很多研究[5,11]。
白冠长尾雉(Syrmaticusreevesii)是我国特有珍稀雉类,隶属鸡形目(Galliformes)雉科(Phasianidae),被列为国家Ⅱ级重点保护野生动物和IUCN易危种[12- 14]。以往对白冠长尾雉的研究主要集中在数量调查[15- 16]、生态习性[17- 18]、集群行为[19- 20]、孵卵[21]、栖息地选择和活动区[22- 23]等方面,关于其行为特别是野生个体活动节律和时间分配方面的资料则相当缺乏。因此,本研究利用红外相机技术,在湖北省广水市研究了野生白冠长尾雉行为的日时间分配及其季节变化,以探讨白冠长尾雉的活动节律。
1 研究地点和方法
1.1 地域概况
研究地点位于湖北省随州广水市蔡河镇平靖关村,地理位置为东经113°54′09″—113°55′21″,北纬31°51′03″— 31°52′40″,与河南省信阳市浉河区接壤(图1)。年平均降水量865—1070 mm,年日照时长2009.6—2059.7 h,年平均气温15.5 ℃,无霜期220—240 d。植被以灌丛和人工林为主。灌丛主要由油桐(Verniciafordii)、小叶鼠李(Rhamnusparvifolia)、野桐(Mallotusjaponicus)、辽东楤木(AraliaelataSeem)和核桃楸(JuglansmandshuricaMaxim)等组成;人工植被主要有茶叶(Camelliasinensis)、板栗(Castaneamollissima)、少量毛白杨(Populustomentosa)和毛竹(Phyllostachyspubescens)以及少量的马尾松(Pinusmassoniana)和杉木(Cunninghamialanceolata)。根据当地气候和白冠长尾雉的习性[23],该地四季划分为春季3—5月、夏季6—8月、秋季9—11月和冬季11月至翌年2月。
1.2 相机布放
2012年3月—2013年3月,在白冠长尾雉活动区内布设ScoutGuard Trail Camera SG550V- 31B 红外相机16台。具体布设规则为:根据对8只白冠长尾雉的无线电跟踪遥测结果,在每只个体的活动区内布设两台红外相机,两台同时工作的红外相机之间相隔至少200 m。根据拍摄效果不定期更换红外相机的位置及拍摄方向,最终布设了40个相机位点(图1)。相机位点多选择郁闭度较高、林下较为开阔且人为活动较少的乔灌丛中,多固定于中小型灌木上,离地面约20—40 cm,确保视野的开阔性,镜头与地面大致平行[24]。拍摄时间为4:00—22:00,相机设置为30 s视频模式,统一相机的日期时间等其它设置。详细记录相机位点编号、经纬度、生境、布设日期和时间、安放时长,一台相机从4:00—22:00正常工作,则记为一个相机日。每1个月检查和更换1次电池及储存卡。
1.3 行为分类
根据相关鸟类行为文献[4,25]及拍摄到的视频资料,将野生白冠长尾雉的行为划分为取食、移动、梳理、警戒、休息、对抗和育幼等7种(表 1)。
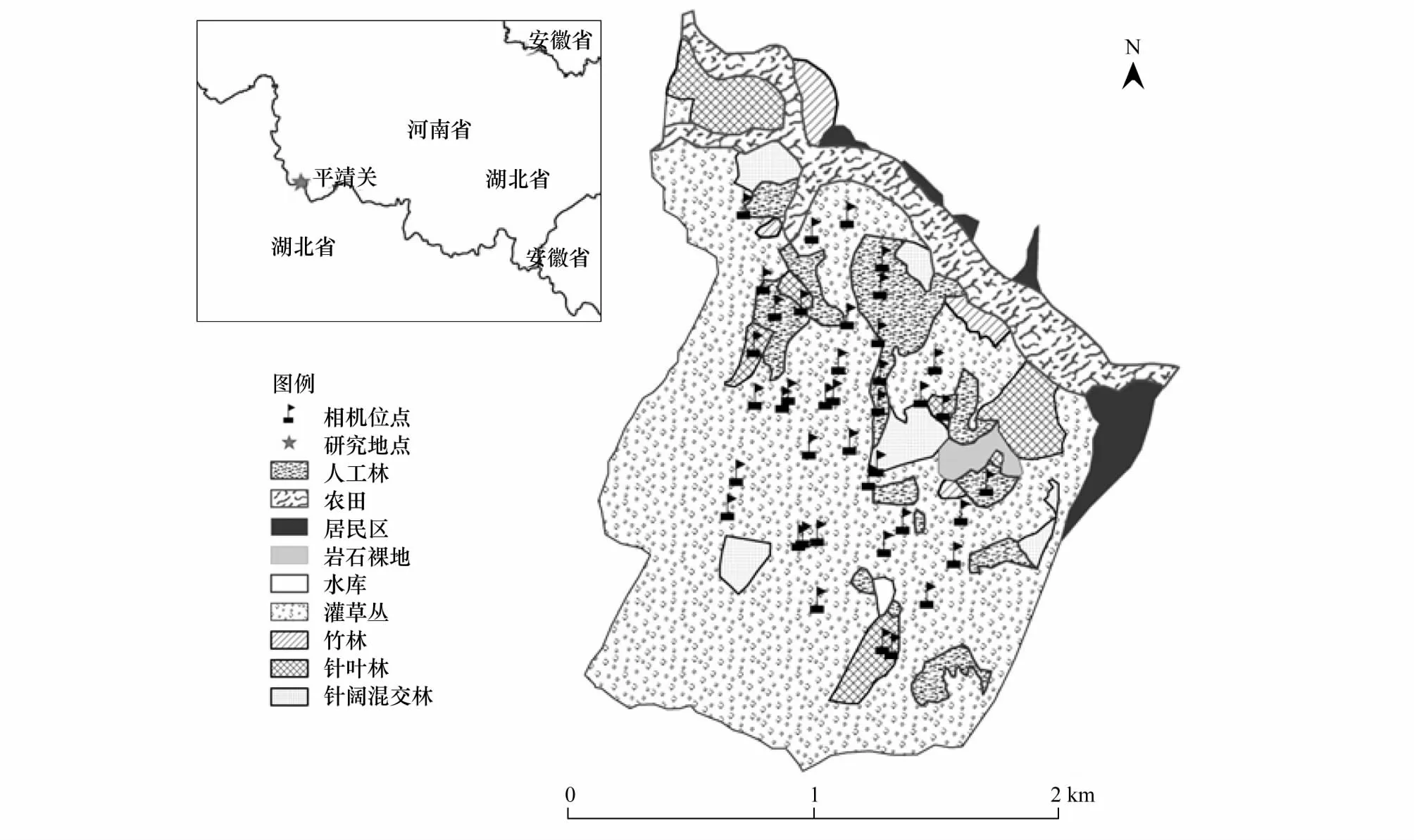
图1 野外研究地点位置、植被及相机布放位点概况 Fig.1 The status of the location, habitat and camera sites of the study area in Hubei Province

行为类型Behaviorclassification描述Descriptionofthecorrespondingofthetypeofbehavior取食行为Feeding在移动过程中啄取地面的各种食物或停留在某地扒掘土取食移动行为Moving双脚交替缓慢或急速地行进和飞腾,包括行走、疾走、飞腾(飞行,上树,下树,原地腾飞)梳理行为Grooming用喙啄理羽毛、用爪搔抓头部及颈前部分、用喙啄跗蹠及足、挺胸拍翅、抖羽等警戒行为Vigilance明显的站立张望,行走中急停观测后又疾走或受惊腾飞休息行为Resting整体基本不动,局部无明显活动,多数时间闭眼,包括躺卧静栖、蹲伏静栖和站立静栖对抗行为Conflicting同种之间或与其它种之间的攻击防御,包括争斗、偷袭、嘻斗、防御、逃匿等育幼行为Raising成年雌鸟与幼鸟共同活动
1.4 数据处理
统一用KMPlayer视频播放软件进行浏览,记录相机位点、视频编号、触发日期、触发时间、时间段、类别(包括白冠长尾雉、其它动物、人为活动等)、物种、数量、雌雄、成幼、行为等数据。在有白冠长尾雉的视频中,对于连续触发的视频,根据目标物种的性别、体型、毛色、尾羽、数量、行为等特征进行判别是否属于同一只(群)。在利用无线电对白冠长尾雉进行遥测的过程中已经发现,除去暴雨大雪等极端气候,同一个体在同一个地方停留的时间一般不超过5 min。因此,对于依据外形特征不能区分的个体或群体,连续超过5 min在同一相机位点出现的个(群)体则被定义为一个独立个(群)体进行分析。所有行为都是记录频次,如一段视频中既有取食行为又有对抗行为,则记录两条数据。
红外相机的拍摄率(CR)为:
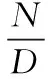
(1)
式中,N为拍摄到的白冠长尾雉的个体数量;D为相机日;CR为拍摄率,按月计算。
日活动强度指数(DII)为:
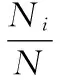
(2)
式中,i为时间段,如6:00—6:59则记为6;Ni为i时段拍摄到的白冠长尾雉个体数量;N为拍摄到的白冠长尾雉总个体数量;DII越大,则表示白冠长尾雉在i时段的活动越强。
日行为频次比例为:
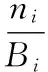
(3)
式中,i同上;k为1,2,…,7,分别代表取食、移动、梳理、警戒、休息、对抗、育幼7种行为;Bi为某日每种行为的总频次;ni为相应行为在该日i时段的频次;Pi表示某种行为在该日i时段的强度。
月行为频次比例:
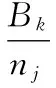
(4)
式中,j为月份;nj为j月各种行为的总频次;Bk为某种行为在j月的总频次;Pj表示k行为在j月所占的行为频次比例。
分析雌雄之间的差异时,先用Kolmogorov-Smirnov Z-test检验数据是否符合正态分布。当数据不符合正态分布时, 使用Mann-Whitney U-检验;当数据符合正态分布时, 使用独立样本的T-检验。用one-way ANOVA分析季节差异,多重比较使用Tukey法。
所有数据处理和统计分析在Microsoft Excel 2003和IBM SPSS statistics 20.0中进行,显著水平为P≤0.05。
2 研究结果
2.1 红外相机拍摄概况
在研究期间内,共安放1774个相机日,拍摄到2242个独立视频。其中,有白冠长尾雉的为408个,占18%;其它动物38%,无效触发35%,人为活动9%。排除相同个(群)体在同一时间内的连续触发及仅有幼鸟的视频,共有333个白冠长尾雉的独立视频(表2),538只次,雌雄比为1.43∶1,记录到396条行为样本。雌雄拍摄率之间不存在显著差异(t22=0.818,P=0.422)。
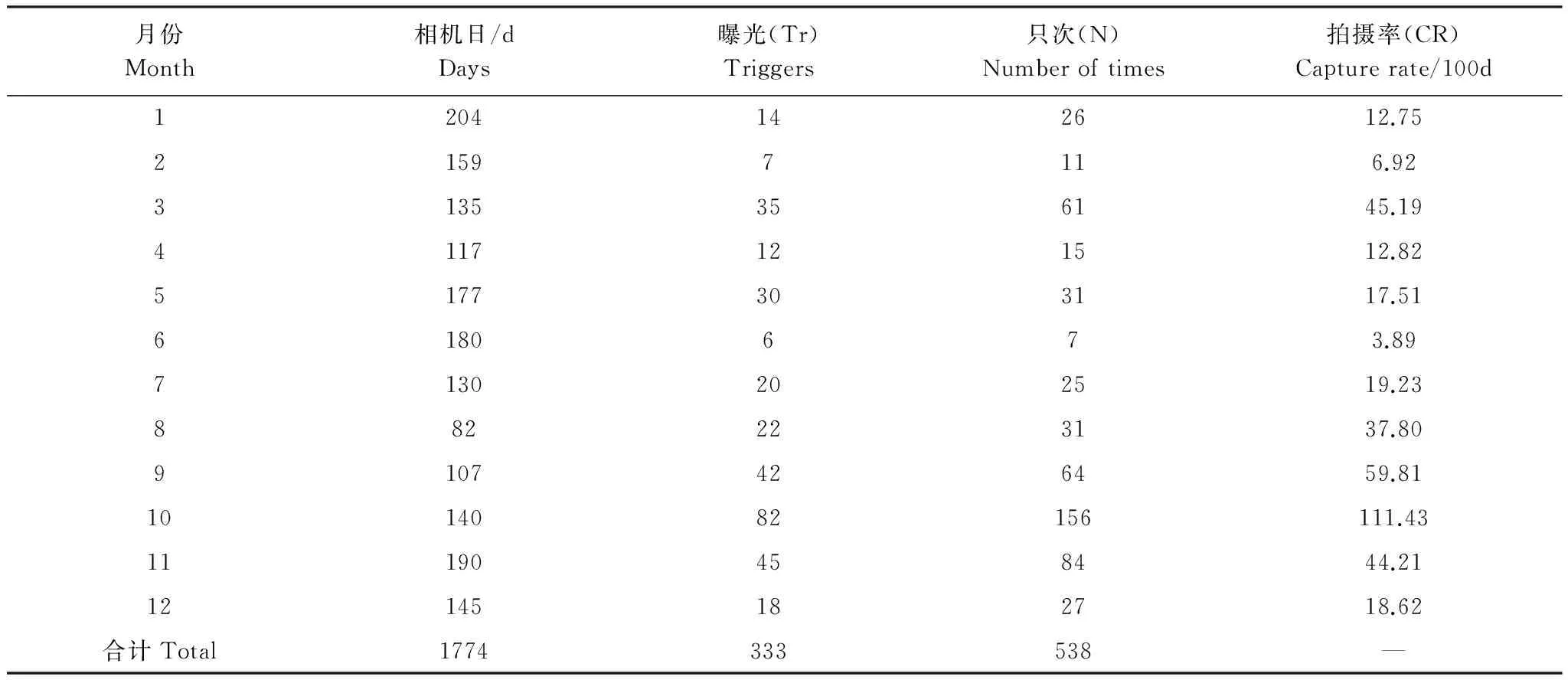
表2 研究区域内红外相机拍摄概况
2.2 日活动节律和时间分配
白冠长尾雉的主要行为是移动和觅食,分别占到总频次的40.71%和33.10%,其余5种行为依次为: 警戒9.29%,梳理7.14%,休息5.00%,对抗2.62%,育幼2.14%。
雌性和雄性白冠长尾雉日活动均有两个高峰期(图2)。雄性个体为7:00—9:00,17:00;雌性个体为9:00—11:00,17:00。雌雄个体在每天不同时间段的活动强度不存在显著差异(Independentttest,t28=-0.03,P=0.976)。取食、移动、警戒和梳理行为节律趋势大致相同,分别在7:00—11:00和17:00达到高峰期;休息行为时间分配相对均匀;育幼行为有一个高峰期,11:00;对抗行为也只有一个高峰期,9:00—10:00(图3)。
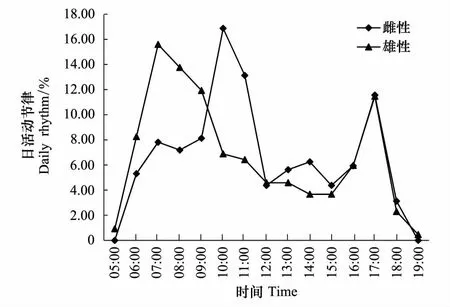
图2 研究区域内野生白冠长尾雉日活动强度及其性别差异Fig.2 The activity intensity and its sexual differences of wild Reeves′s Pheasants in the study area
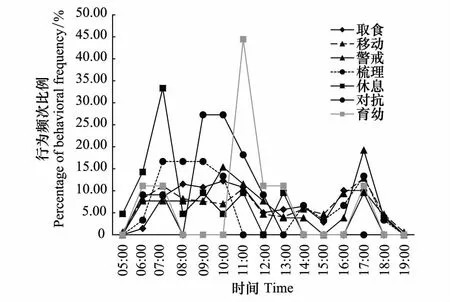
图3 研究区域内野生白冠长尾雉日活动节律Fig. 3 Diurnal behavioral rhythms of wild Reeves′s Pheasant in the study area
育幼是雌性白冠长尾雉单独完成的,对抗行为的频次比例不符合正态分布(Plt;0.05)。雌性白冠长尾雉的警戒行为频次比例显著高于雄性个体(图4,t22=2.782,P=0.011),取食、移动、梳理、休息行为则无显著差异(Pgt;0.05);对抗行为频次比例在雌雄间也不存在显著差异(图4,Mann-Whitney U test,Z=-0.569,P=0.569)。
2.3 季节差异
不同季节之间白冠长尾雉取食行为(One-way ANOVA,F3=6.552,P=0.015)、移动行为(One-way ANOVA,F3=5.268,P=0.027)、对抗行为(One-way ANOVA,F3=25.687,Plt;0.01)有显著差异,而警戒行为(One-way ANOVA,F3=0.148,P=0.928)、梳理行为(One-way ANOVA,F3=1.015,P=0.435)、休息行为(One-way ANOVA,F3=0.776,P=0.540)、育幼行为(One-way ANOVA,F3=0.682,P=0.588)则无显著差异(图5)。白冠长尾雉在冬季的取食行为比例明显高于夏季(Tukey post hoc test,P=0.013);移动行为比例夏季最高,显著高于秋季(Tukey post hoc test,P=0.046)和冬季(Tukey post hoc test,P=0.035);对抗行为仅仅在秋季被记录到。
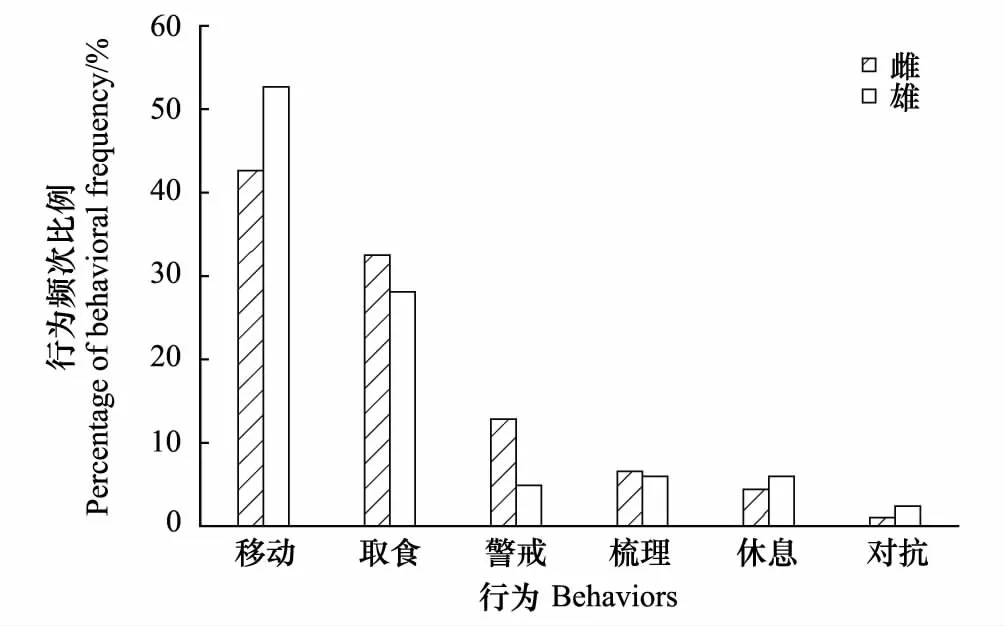
图4 研究区域内雌雄野生白冠长尾雉行为频次比例差异Fig. 4 Sexual differences of behavioral frequency rate of wild Reeves′s Pheasant in the study area
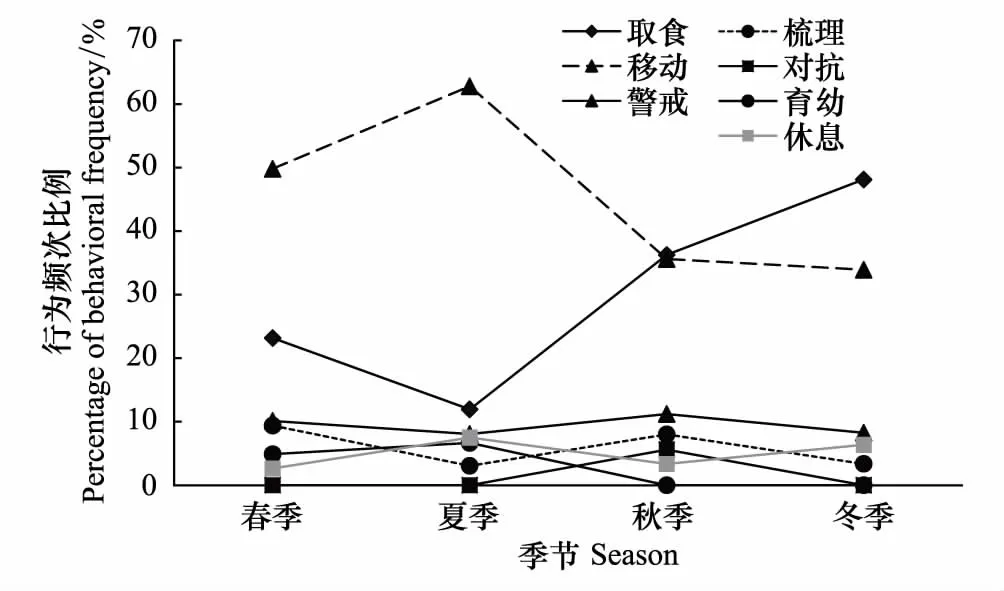
图5 研究区域内不同季节野生白冠长尾雉行为频次比例差异Fig. 5 Seasonal differences of behavioral frequency rate of wild Reeves′s Pheasant in the study area
3 讨论
根据野外拍摄的538只次成体,当地雌雄比例为1.43∶1。这是分析野生白冠长尾雉性比的一次尝试。由于野外研究的难度,白冠长尾雉性比方面的资料相对较为缺乏,并且不同的研究结果也不一样,如雌雄性比有2.18∶1[20],3∶1、0.67∶1[26],而利用标图法结合无线电遥测的调查得到的结果为1.1∶1[20],这与红外相机监测的结果比较接近。
动物的活动强度与被相机拍摄到的概率呈正相关[27]。野生白冠长尾雉每天有两个活动高峰期,这与之前的报道相符[17]。虽然雌雄在不同时间段的活动强度不存在显著差异,但本研究也发现雄性上午的活动高峰期要早于雌性,这与在贺兰山地区对石鸡(Alectorischukar)的研究结果[28]相似:雄性石鸡的觅食最低峰和静息的最高峰出现在13:30, 雌性石鸡觅食最低峰和静息的最高峰则出现在12:30,雌雄个体间也存在一定差异。
基于自身营养的需要,野生动物通常将大部分的时间用于觅食,尤其是在食物缺乏的季节[29- 30]。野生白冠长尾雉的取食、移动、警戒行为日节律的变化与日活动强度相关,在上午和下午均分别各有一个峰值,且其移动和取食行为频次比例共占到了记录行为的73.81%,其中移动行为占40.71%。野生白冠长尾雉休息行为频次比例很低,尽管无法将其与笼养环境下的白冠长尾雉行为进行比较,但对笼养黄腹角雉(Tragopancaboti)的行为观察发现,休息行为占到了一半以上[25]。这可能是野外条件下食物资源丰富度、天敌分布及活动空间竞争等与笼养条件下明显不同而导致的[1]。雌性白冠长尾雉警戒行为频次比例显著高于雄性个体,这在一些哺乳动物的研究中也有发现[31]。野生白冠长尾雉取食、移动、梳理、休息行为在性别间并无显著差异,这可能跟这些行为的刚性强有关[32]。因为动物的行为有刚性与弹性之分,当动物的能量来源和生存空间受限制时, 一些诸如摄食、排遗等生存所必需的生理行仍将表现出来,则这类行为即为刚性行为[32]。
由于不同季节之间,食物资源不同,动物个体可根据能量需求采取最优行为时间分配策略[30]。白冠长尾雉在冬季的取食行为比例明显高于夏季(P=0.013),可能是因为夏季草本植物、樱桃和鼠李等落果资源较为丰富,而这个时候白冠长尾雉的集群率又比较低[20],个体容易满足自身的能量需求;拍摄的视频也发现,白冠长尾雉在夏季喜欢固定下来啄食青草,而冬季则大多数是走动觅食。夏季的移动行为显著高于秋季和冬季,这与利用无线电遥测技术监测白冠长尾雉的结果相似,即相对于繁殖期,白冠长尾雉冬季的活动区较小,活动能力也相对较弱[22]。
本研究所在区域位于村庄周围,人为干扰较重,在野外也发现很多诸如放牧、砍伐、采草药等现象,这可能对白冠长尾雉的行为产生影响。受研究时间限制,本研究并没有将其与人为干扰较少条件下的白冠长尾雉行为进行比较。目前,白冠长尾雉种群很多已经纳入自然保护区范围[12],研究这些自然保护区内野生白冠长尾雉的行为,将为进行相关比较研究提供良好的平台,而这对于评估人为活动对白冠长尾雉行为的影响并提出相应的保护对策将具有重要意义。
本研究的实践也证明,红外相机技术非常适合研究警觉性高、数量稀少的大型地栖林鸟[11,33],包括白冠长尾雉。虽然利用红外相机技术进行鸟类行为研究具有一定的局限性,例如其不能对同一只个体进行连续观测[11],但其特有的优点却能弥补传统方法的许多不足[5- 6],因为以往对野生动物行为的研究,很多都需要佩戴项圈或发射器等[34- 35],这可能会对野生动物的行为产生影响;而在野外进行隐蔽观测,则易受观测人员主观因素的影响,耗费时间且不具有长期性。
致谢:白洁同学参加了部分野外工作,视频整理得到了曹婉露和王秦韵同学的帮助,张鹏对监测工作提供后勤保障,特此致谢。
[1] Shang Y C. Ethology. Beijing: Peking University Press, 2005.
[2] Long S, Zhou C Q, Wang W K, Pan L, Hu J C. Diurnal behavioral rhythm, time budgets and group behavior of dwarf blue sheep in summer. Zoological Research, 2009, 30(6): 687- 693.
[3] Yi G D, Yang Z J, Liu Y, Zuo B, Zhao J, Hao X L. Behavioral time budget and daily rhythms for winteringMergussquamatus. Acta Ecologica Sinica, 2010, 30(8): 2228- 2234.
[4] Xing M Z, Zhang Z M, Tian H J, Wen J B. Time budget and activity rhythm of captive great spotted woodpecker. Chinese Journal of Zoology, 2013, 48(1): 95- 101.
[5] Tricia L C, Don E S. Using remote photography in wildlife ecology: a review. Wildlife Society Bulletin, 1999, 27(3): 571- 581.
[6] Lu X L, Jiang Z G, Tang J R, Wang X J, Xiang D Q, Zhang J P. Auto-trigger camera traps for studying giant panda and its sympatric wildlife species. Acta Zoologica Sinica, 2005, 51(3): 495- 500.
[7] Karanth K U. Estimating tigerPantheratigrispopulations from camera-trap data using capture-recapture models. Biological Conservation, 1995, 71(3): 333- 338.
[8] Foster M L, Humphrey S R. Use of highway underpasses by Florida panthers and other wildlife. Wildlife Society Bulletin, 1995, 23(1): 95- 100.
[9] Karanth K U, Nichols J D. Estimation of tiger densities in India using photographic captures and recaptures. Ecology, 1998, 79(8): 2852- 2862.
[10] Silver S C, Ostro L E T, Marsh L K, Maffei L, Noss A J, Kelly M J, Wallace R B, Gomez H, Ayala G. The use of camera traps for estimating jaguarPantheraoncaabundance and density using capture/recapture analysis. Oryx, 2004, 38(2): 148- 154.
[11] O′Brien T G, Kinnaird M F. A picture is worth a thousand words: the application of camera trapping to the study of birds. Bird Conservation International, 2008, 18(S1): S144-S162.
[12] Zheng G M, Wang Q S. China Red Data Book of Endangered Animals (Aves). Beijing: Science Press, 1998: 182- 184.
[13] Collar N J. Threatened Birds of Asia: The Birdlife International Red Data Book (Part A). Cambridge: Bird Life International, 2001: 1001- 1009.
[14] IUCN. IUCN Red List of Threatened Species. (2012- 02) [2013- 05- 07]. http://www.iucnredlist.org/details/100600271/0.
[15] Wu Z K, Xu W S. On the geographical distribution and abundance of white crowned long tailed pheasants in Guizhou Province. Zoological Research, 1987, 8(1): 13- 19.
[16] Wu Z K, Li Z M, Xu W S. Distribution and population density of the Reeves′s Pheasant. Guizhou Science, 1991, 9(3): 215- 219.
[17] Wu Z K. The ecology of Reeves′s pheasant.. Chinese Journal of Zoology, 1979, (3): 16- 18.
[18] Fang C L, Ding Y H. Over-wintering ecology of the white-crowned long-tailed pheasant. Chinese Journal of Ecology, 1997, 16(2): 67- 68.
[19] Sun Q H, Zhang Z W, Ruan X F, Zhang K Y, Zhu J G. Studies on flocking behavior of Reeves′s Pheasant in Dongzhai nature reserve, Henan province. Journal of Beijing Normal University: Natural Science, 2001, 37(1): 111- 117.
[20] Zhang X H, Xu J L, Zhang Z W, Zheng G M, Ruan X F, Xie F L. Flocking behavior of Reeves′s Pheasants (Syrmaticusreevesii) at two sites in Henan and Shaanxi. Zoological Research, 2004, 25(2): 89- 95.
[21] Zhang X H, Xu J L, Zhang Z W, Xie F L, Zhang K Y, Zhu J G. A study on the incubation behavior of Reeves′s Pheasant (Syrmaticusreevesii) by radio tracking. Journal of Beijing Normal University: Natural Science, 2004, 40(2): 255- 259.
[22] Xu J L, Zhang X H, Zhang Z W, Zhang G M, Ruan X F, Zhang K Y. Home range and habitat use of male Reeves′s pheasant (Syrmaticusreevesii) in winter in Dongzhai National Nature Reserve, Henan Province. Biodiversity Science, 2005, 13(5): 416- 423.
[23] Xu J L, Zhang Z W, Wang Y, Connelly J W. Spatio-temporal responses of male Reeves′s PheasantsSyrmaticusreevesiito forest edges in the Dabie Mountains, central China. Wildlife Biology, 2011, 17(1): 16- 24.
[24] Zhang S S, Bao Y X, Wang Y N, Fang P F, Ye B. Comparisons of different camera trap placement patterns in monitoring mammal resources in Gutianshan National Nature Reserve. Chinese Journal of Ecology, 2012, 31(8): 2016- 2022.
[25] Wen Z Q, Zheng G M. A study on the behaviours of cabot′s tragopan in breeding season. Journal of Beijing Normal University: Natural Science, 1997, 33(2): 263- 269.
[26] Lu T C. The Rare and Endangered Gamebirds in China. Fuzhou: Fujian Science and Technology Press, 1991: 314- 355.
[27] Li M F, Li S, Wang D J, William J M, Guan T P, Chen L M. The daily activity patterns of takinBudorcastaxicolorin winter and spring at Tangjiahe Nature Reserve, Sichuan Province. Sichuan Journal of Zoology, 2011, 30(6): 850- 855.
[28] Zhou X Y, Wang X M, Jiang Z H. Time budget and activity rhythm ofAlectorischukarin winter. Journal of Northeast Forestry University, 2008, 36(5): 44- 46.
[29] Pépin D, Renaud P C, Dumont B, Decuq F. Time budget and 24-h temporal rest-activity patterns of captive red deer hinds. Applied Animal Behaviour Science, 2006, 101(3/4): 339- 354.
[30] Lian X M, Li X X, Yan P S, Zhang T Z, Su J P. Behavioural time budgets and diurnal rhythms of the female Tibetan gazelles in the Kekexili National Nature Reserve. Acta Ecologica Sinica, 2012, 32(3): 663- 670.
[31] Liu Z S, Wu J P, Teng L W. Time budget and behavior pattern of semi-freeCervusnipponin spring. Chinese Journal of Ecology, 2002, 21(6): 29- 32.
[32] Jiang Z G, Li C W, Peng J J, Hu H J. Structure, elasticity and diversity of animal behavior. Biodiversity Science, 2001, 9(3): 265- 274.
[33] Li S, Mcshea W J, Wang D J, Shao L K, Shi X G. The use of infrared-triggered cameras for surveying phasianids in Sichuan Province, China. Ibis, 2010, 152(2): 299- 309.
[34] Moen R, Pastor J, Cohen Y, Schwartz C C. Effects of moose movement and habitat use on GPS collar performance. The Journal of Wildlife Management, 1996, 60(3): 659- 668.
[35] Wong S T, Servheen C W, Ambu L. Home range, movement and activity patterns, and bedding sites of Malayan sun bearsHelarctosmalayanusin the Rainforest of Borneo. Biological Conservation, 2004, 119(2): 169- 181.
参考文献:
[1] 尚玉昌. 动物行为学. 北京: 北京大学出版社, 2005.
[2] 龙帅, 周材权, 王维奎, 潘立, 胡锦矗. 矮岩羊夏季活动节律、时间分配和集群行为. 动物学研究, 2009, 30(6): 687- 693.
[3] 易国栋, 杨志杰, 刘宇, 左斌, 赵匠, 郝锡联. 中华秋沙鸭越冬行为时间分配及日活动节律. 生态学报, 2010, 30(8): 2228- 2234.
[4] 邢茂卓, 张志明, 田恒玖, 温俊宝. 笼养大斑啄木鸟行为时间分配和活动节律. 动物杂志, 2013, 48(1): 95- 101.
[6] 卢学理, 蒋志刚, 唐继荣, 王学杰, 向定乾, 张建平. 自动感应照相系统在大熊猫以及同域分布的野生动物研究中的应用. 动物学报, 2005, 51(3): 495- 500.
[12] 郑光美, 王岐山. 中国濒危动物红皮书(鸟类). 北京: 科学出版社, 1998: 182- 184.
[15] 吴至康, 许维枢. 白冠长尾雉 (Syrmaticusreevesii) 在贵州的分布与数量. 动物学研究, 1987, 8(1): 13- 19.
[17] 吴至康. 白冠长尾雉的生态. 动物学杂志, 1979, (3): 16- 18.
[18] 方成良, 丁玉华. 白冠长尾雉的越冬生态. 生态学杂志, 1997, 16(2): 67- 68.
[19] 孙全辉, 张正旺, 阮祥峰, 张可银, 朱家贵. 白冠长尾雉集群行为的初步研究. 北京师范大学学报: 自然科学版, 2001, 37(1): 111- 117.
[20] 张晓辉, 徐基良, 张正旺, 郑光美, 阮祥锋, 谢福录. 河南陕西两地白冠长尾雉的集群行为. 动物学研究, 2004, 25(2): 89- 95.
[21] 张晓辉, 徐基良, 张正旺, 谢福录, 张可银, 朱家贵. 白冠长尾雉孵卵行为的无线电遥测研究. 北京师范大学学报: 自然科学版, 2004, 40(2): 255- 259.
[22] 徐基良, 张晓辉, 张正旺, 郑光美, 阮祥锋, 张可银. 白冠长尾雉雄鸟的冬季活动区与栖息地利用研究. 生物多样性, 2005, 13(5): 416- 423.
[24] 章书声, 鲍毅新, 王艳妮, 方平福, 叶彬. 不同相机布放模式在古田山兽类资源监测中的比较. 生态学杂志, 2012, 31(8): 2016- 2022.
[25] 温战强, 郑光美. 黄腹角雉的繁殖期行为研究. 北京师范大学学报: 自然科学版, 1997, 33(2): 263- 269.
[26] 卢汰春. 中国珍稀濒危野生鸡类. 福州: 福建科学技术出版社, 1991: 314- 355.
[27] 李明富, 李晟, 王大军, William J M, 官天培, 谌利民. 四川唐家河自然保护区扭角羚冬春季日活动模式研究. 四川动物, 2011, 30(6): 850- 855.
[28] 周晓禹, 王晓明, 姜振华. 贺兰山石鸡越冬期昼间行为时间分配及活动规律. 东北林业大学学报, 2008, 36(5): 44- 46.
[30] 连新明, 李晓晓, 颜培实, 张同作, 苏建平. 夏季可可西里雌性藏原羚行为时间分配及活动节律. 生态学报, 2012, 32(3): 663- 670.
[31] 刘振生, 吴建平, 滕丽微. 散放条件下春季梅花鹿行为时间分配的研究. 生态学杂志, 2002, 21(6): 29- 32.
[32] 蒋志刚, 李春旺, 彭建军, 胡慧建. 行为的结构、刚性和多样性. 生物多样性, 2001, 9(3): 265- 274.
ActivityrhythmandbehavioraltimebudgetsofwildReeves′sPheasant(Syrmaticusreevesii)usinginfraredcamera
ZHAO Yuze1, WANG Zhichen2, XU Jiliang1,*, LUO Xu1, AN Lidan3
1CollegeofNatureConservation,BeijingForestryUniversity,Beijing100083,China2AcademyofForestInventoryandPlanning,StateForestryAdministration,Beijing100714,China3DepartmentofWildFaunaandFloraConservationandNatureReserveManagement,StateForestryAdministration,Beijing100714,China
The researches on activity rhythm and behavioral time budgets of vulnerable or endangered species were of great importance to form the effective conservation strategies, Reeves′s Pheasant (Syrmaticusreevesii) was a vulnerable species endemic to China, whereas little information on activity rhythm and behavioral time budgets of this species was available. Therefore, we used infrared camera technology, from March 2012 to March 2013, to survey the activity rhythm and behavioral time budgets of wild Reeves′s Pheasant (Syrmaticusreevesii) in Caihe Town, Guangshui City, Hubei province. We radio tracked and outlined 8 Reeves′s Pheasants in the study area at first, and then we determined 4—5 sites as candidate camera sites in each home range. We selected two camera sites with a distance apart from at least 200 m in a home range synchronously, and irregularly moved these cameras across the candidate camera sites in this home range. In consequence, we set 16 infrared cameras at 40 candidate camera sites. The daily work time of a camera was set from 4:00 to 22:00 with the reference to some previous reports on this pheasant behavior, and we defined it as a camera day. We also divided the behavior of wild Reeves′s Pheasant into seven types, including feeding, moving, grooming, vigilance, resting, conflicting, and raising According to the available information on pheasant behavior. As a result, the total work time of these cameras in the field added up to 1774 d. We obtained totally 2242 videos in relation to Reeves′s Pheasants that were responsible for about 18% with a total of 538 individuals in relation to 396 behavior samples. The sex ratio between the female and male photoed is 1.43∶1. Two diurnal activity peaks were observed for the wild individuals, and the activity peak in the morning was around 9:00 and that in the afternoon was around 17:00. In particular, the activity peak of the males in the morning was much earlier about two hours than that of females, i.e. 7:00—9:00 for the males and 9:00—11:00 for the females. The behavior of moving and feeding dominated the diurnal behaviors of Reeves′s Pheasant, accounting for 40.71% and 33.10%, respectively, and then followed in order by vigilance (9.29%), grooming (7.14%), resting (5.00%), conflicting (2.62%), raising (2.14%). The frequency of vigilance of females was significantly higher than that of the males. The wild Reeves′s Pheasant also showed significant seasonal behavior variations, and the rate of feeding, moving, and conflicting varied among seasons. For example, the feeding frequency in winter was much higher than that in summer (Plt;0.05), and the moving frequency in summer was higher than that in autumn (Plt;0.05) and winter (Plt;0.05), whereas the frequency of conflicting, grooming, resting, and raising did not differ significantly. Given this study was conducted in an area with higher human disturbance, it was reasonable to recommend these results to that happened in an area with less human disturbance (e.g. nature reserves) in the future. The practice of this study also proved that infrared camera technology is very suitable for studying highly alert and rare terrestrial forest birds.
infrared triggered camera technique;Reeves′s Pheasant;activity rhythm; behavioral time budget
林业公益性行业科研专项(200904003);国家自然科学基金项目(31172115)
2013- 06- 04;
2013- 07- 25
*通讯作者Corresponding author.E-mail: xujiliang@bjfu.edu.cn
10.5846/stxb201306041335
Zhao Y Z, Wang Z C, Xu J L, Luo X, An L D.Activity rhythm and behavioral time budgets of wild Reeves′s Pheasant (Syrmaticusreevesii) using infrared camera.Acta Ecologica Sinica,2013,33(19):6021- 6027.
