组蛋白去乙酰基酶在前列腺癌中的作用及机制研究
2013-10-17黄勇闫骏马敏
黄 勇 闫 骏 马 敏
内蒙古医科大学附属医院泌尿外科,内蒙古呼和浩特 010050
前列腺癌是一种常见于老年男性的恶性肿瘤,在欧美等地发病率位居恶性肿瘤首位[1]。组蛋白修饰是组织细胞发育、分化基因转录调控的关键分子机制,组蛋白修饰状态改变在肿瘤发生及进展中具有重要作用。组蛋白修饰方式包括乙酰化、甲基化、泛素化、磷酸化修饰等[2-4],核心组蛋白乙酰化水平的稳态是由组蛋白乙酰转移酶和组蛋白脱乙酰化酶(HDACs)之间的动态平衡调节的[5]。HDACs在许多异常活化的癌细胞中均存在,可阻抑细胞周期抑制因子的作用。组蛋白乙酰化状态失衡参与众多信号转导通路调节,是细胞癌变中重要的转录后修饰过程[6]。因此,HDAC是重要的抗癌药物靶点,在抗癌药物研究中具有重要意义[6]。
丙戊酸(VPA)是一种HDAC抑制剂(HDACI)和抗癫痫药物,可抑制Ⅰ类和Ⅱ类组蛋白脱乙酰基转移酶(HDACs)[7]。研究发现,长期服用VPA可减少前列腺癌症细胞的增殖[8-10]。因此,本研究观察了HDAC抑制剂VPA对在体前列腺癌移植体的作用,以期明确组蛋白去乙酰基酶在前列腺癌中的作用及其机制。
1 材料与方法
1.1 材料
人前列腺癌的细胞系LNCaP购自American Type Culture Collection(Manassas,VA),雄性无胸腺 nu/nu 小鼠(20~25 g)购自天津医科大学实验动物中心。L-谷氨酰胺的 RPMI 1640 培养基为 Cellgro(Herndon,VA)公司产品,热灭活胎牛血清购自 Life Technologies,Inc.(Carlsbad,CA),盐酸环丙沙星和庆大霉素购自US Biological公司(Swampscott,MA),基底胶为 BD Biosciences(Palo Alto,CA)公司产品,VPA(1 mol/L 的 VPA 钠盐;Sigma,St.Louis,MO)。
一抗来源:乙酰化组蛋白H3为美国Upstate公司产品,p21WAF1/CIP1及雄激素受体(AR)为美国Santa Cruz公司产品,细胞周期蛋白D1为美国Ventana Medical Systems公司产品,p27KIP1为美国DAKO公司产品。
主要仪器:Leica CM 1850冰冻切片机为德国徕卡公司产品,Olympus BX51显微镜为日本奥林巴斯公司产品。
1.2 肿瘤细胞株培养及移植瘤建立
细胞培养环境:含L-谷氨酰胺的RPMI 1640培养基,10%热灭活胎牛血清,5 μg/mL的盐酸环丙沙星和50 μg/mL庆大霉素。细胞生长至铺满培养基80%~90%时,用含0.05%胰蛋白酶的EDTA(0.53 mmol/L)收集细胞,细胞重悬在PBS(pH 7.4),与混合 1×基底胶(BD Biosciences,Palo Alto,CA)混匀,皮下注入(1×106/次)雄性无胸腺nu/nu小鼠,建立肿瘤移植体。
1.3 动物分组及处理
移植瘤建立后,动物随机分成对照组和HDAC抑制剂组,每组6只。HDAC抑制剂VPA溶于PBS,经0.22 μm过滤器灭菌,HDAC抑制剂组受试动物接受0.4%w/v的VPA饮用水,对照组为不含VPA正常饮用水,35 d后切除肿瘤。
1.4 免疫组化染色
切除肿瘤移植体经福尔马林液固定,组织块依次浸入10%、20%、30%的蔗糖溶液脱水,然后经OCT包埋,行连续冠状恒冷箱切片,厚度16 μm,组织切片经水化后,3%的过氧化氢孵育 10 min,PBS冲洗(3 min×3次),滴加马血清封闭液,孵育10 min,一抗4℃孵育过夜(一抗使用浓度:乙酰化组蛋白 H3 为 1∶1000,p21WAF1/CIP1为 1∶500,细胞周期蛋白 D1 为 1∶1000,p27KIP1为 1∶1000,AR 为 1∶250),37℃复温45 min,PBS洗 5 min×3 次,滴加相应二抗,37℃孵育 45 min,PBS冲洗,5 min×3次,DAB显色,苏木精复染,梯度酒精脱水,二甲苯透明,中性树胶封片,光镜下观察。应用Image Pro Plus6.0图像分析软件阳性细胞进行平均光密度(MOD)分析。
1.5 统计学方法
采用SPSS 13.0统计学软件进行数据分析,计量资料数据用均数±标准差(±s)表示,两组间比较采用t检验;以P<0.05为差异有统计学意义。
2 结果
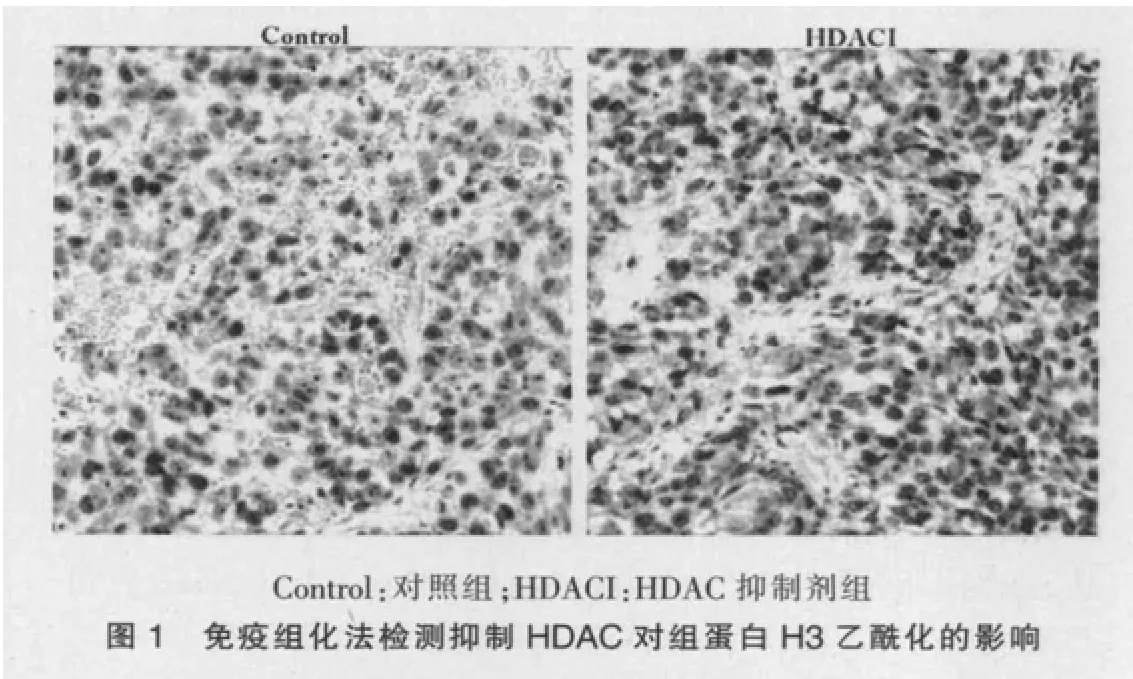
2.1 抑制HDAC对前列腺癌移植体组蛋白H3乙酰化的影响
给予VPA饮水后,肿瘤移植体乙酰化组蛋白H3与未处理的对照组相比,阳性染色细胞数明显增加,乙酰化组蛋白H3水平显著升高(P<0.05),表明抑制HDAC可提高乙酰化组蛋白H3的表达。见图1、表1。
2.2 抑制HDAC对前列腺癌移植体细胞周期的影响
免疫组化检测结果表明:给予组蛋白乙酰化酶抑制剂VPA后,前列腺癌移植体细胞p21WAF1/CIP1水平显著升高(P<0.05,图2、表1),提示细胞周期被有效阻滞;进一步对细胞周期蛋白D1及p27KIP1的检测结果显示,HDAC抑制剂组D1蛋白水平与对照组相比显著下降(P<0.05,图2、表1),而p27KIP1表达水平则比对照组明显增加(P<0.05,图2、表1),这进一步提示,抑制HDAC可明显阻滞细胞周期,抑制癌细胞生长。

2.3 抑制HDAC对前列腺癌移植体AR表达的影响
AR是雄激素作用的中介,与前列腺癌的发生发展密切相关。本研究结果显示,给予组蛋白乙酰化酶抑制剂VPA后,前列腺癌移植体细胞AR的表达水平较对照组显著降低(P<0.05,图3、表1),提示抑制HDAC可通过阻断雄激素的作用途径抑制前列腺癌细胞发展。
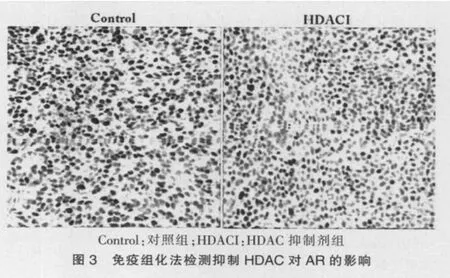
表1 抑制HDAC对前列腺癌移植体各标志物免疫组化阳性染色MOD 的影响(±s,n=6)

表1 抑制HDAC对前列腺癌移植体各标志物免疫组化阳性染色MOD 的影响(±s,n=6)
注:AR:雄激素受体
组别 H3 P21WAF1/CIP1P27KIP1细胞周期蛋白D1 AR对照组HDAC抑制剂组P值0.63±0.12 0.75±0.11 0.27±0.06 0.79±0.10 0.31±0.08 1.09±0.18 0.69±0.09 0.58±0.08 0.62±0.10 0.53±0.07 0.0360.0250.020.0430.046
3 讨论
以HDAC为抑制靶标的药物可重新激活沉默的肿瘤抑制基因,并具有选择性、多效性的抗肿瘤作用[11]。大部分人类癌症高度异质性,可能涉及许多不同的信号通路。体外和在体研究表明,HDACI可抑制多种活性[12-14],长期服用HDAC抑制剂VPA可显著减少前列腺癌细胞增殖,肿瘤体积明显减小[8]。为了进一步探讨HDAC在前列腺癌作用及机制,笔者研究了HDACI对异种移植前列腺癌细胞周期和增殖相关标志物表达的影响。
细胞周期由周期蛋白和周期蛋白依赖激酶(cyclindependent kinase,CDK)的相互作用而完成。 P21WAF1/CIP1是细胞周期负调节因子家族周期蛋白依赖激酶抑制因子(cyclin-dependent kinase inhibitors,CDKI) 家族的一员,转化细胞中p21WAF1/CIP1的表达对细胞周期阻滞起重要作用[15],其可通过抑制Cyclin-CDK2复合体的活性及阻断DNA复制阻滞细胞周期[16]。p27KIP1同样被证实在阻滞G1期到S期细胞周期中发挥重要作用[17]。本研究结果证实,抑制HDAC可促进前列腺癌移植体细胞p21WAF1/CIP1和p27KIP1表达,表明HDAC与细胞周期相关蛋白密切相关,通过抑制HDAC而上调细胞周期负调节因子家族周期蛋白依赖激酶抑制因子的表达,可抑制癌细胞增殖。
Cyclin D1是G1~S细胞周期进程中重要的正向调控因子,其可通过与CDK4/6结合形成复合体而激活后者,进而使视网膜母细胞瘤抑制基因失活,加速癌细胞周期进程[18-19]。在多种肿瘤组织中,均发现Cyclin D1高表达。本研究结果显示,抑制HDAC可降低前列腺癌移植体细胞Cyclin D1表达水平,这可能是HDACI抑制癌细胞增殖的机制之一。
前列腺癌的生长是雄激素依赖性的,AR及其信号转导通路在前列腺癌的发生发展中具有重要作用,AR高表达可增强前列腺癌细胞对低水平雄激素的敏感性,进而导致前列腺癌在抗雄激素治疗一段时间后,大多数最终发展为雄激素非依赖性前列腺癌[20-22]。本研究表明,抑制HDAC可下调前列腺癌移植体细胞AR表达水平,这提示HDACI可通过AR信号转导通路抑制癌细胞生长。
综上,HDAC参与前列腺癌细胞的细胞周期调控并与AR过表达有关,HDACI可通过诱导癌细胞周期阻滞及下调AR表达水平等机制抑制前列腺癌肿瘤移植体生长,HDAC可作为前列腺癌治疗药物的研制靶标。
[1]Suikki HE,Kujala PM,Tammela TL,et al.Genetic altera tions and changes in expression of histone demethylases in prostate cancer[J].Prostate,2010,70(8):887-898.
[2]Secombe J,Eisenman RN.The function and regulation of the JARID1 family of histone H3 lysine 4 demethylases:the Myc connection[J].Cell Cycle,2007,6(11):1324-1328.
[3]Blair LP,Cao J,Zou MR,et al.Epigenetic regulation by lysine demet hylase 5(KDM5)enzymes in cancer[J].Cancers(Base1),2011,3(1):1383-1404.
[4]孙圣坤,于晓妉,史立新,等.组蛋白去乙酰化酶抑制剂FK228对前列腺癌LNCaP细胞抑制效应的细胞信号通路研究[J].现代泌尿生殖和肿瘤杂志,2009,1(1):32-33.
[5]Rephaeli A,Blank-Porat D,Tarasenko N,et al.In vivo and in vitro antitumor activity of butyroyloxymethyl-diethylphosphate(AN-7),a histone deacetylase inhibitor,in human prostate cancer[J].Int J Cancer,2005,116(2):226-235.
[6]Konstantinopoulos PA,Karamouzis MV,Papavassiliou AG.Focus on acetylation:the role of histone deacetylase inhibitors in cancer therapy and beyond[J].Expert Opin Invest Drugs,2007,16(5):569-571.
[7]Gottlicher M,Minucci S,Zhu P,et al.Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells[J].EMBO J,2002,20(24):6969–6978.
[8]Xia Q,Sung J,Chowdhury W,et al.Chronic administration of valproic acid inhibits prostate cancer cell growth in vitro and in vivo[J].Cancer Res,2006,66(4):7237–7244.
[9]Shabbeer S,Kortenhorst MSQ,Kachhap S,et al.Multiplemolecular path ways explain the anti-proliferative effect of valproic acid on prostate cancer cells in vitro and in vivo[J].Prostate,2007,6(10):1099-1110.
[10]Chen CL,Sung J,Cohen M,et al.Valproic acid inhibits invasiveness in bladder cancer but not in prostate cancer cells[J].J Pharmacology Exp Therap,2006,319(2):533–542.
[11]Rosato RR,Grant S.Histone deacetylase inhibitors:insights intome chanisms of lethality[J].Expert Opin Therapeutic Targets,2005,9(4):809-824.
[12]Sambucetti LC,Fischer DD,Zabludoff S,et al.Histone deacetylase inhibition selectively alters the activity and expression of cell cycle proteins leading to specific chromatin acetylation and antiproliferative effects[J].J Biolo Chem,1999,274(49):34940-34947.
[13]Iwata S,Saito T,Ito Y,et al.Antitumor activities of valproic acid on Epstein-Barr virus-associated T and natural killer lymphoma cells[J].Cancer Sci,2012,103(2):375-381.
[14]Kuendgen A,Gattermann N.Valproic acid for the treatment of myeloid malignancies[J].Cancer,2007,110(5):943-954.
[15]Elledge SJ,Winston J,Harper JW.A question of balance: the role of cyclin-kinase inhibitors in development and tumorigenesis[J].Trends Cell Biol,1996,6(3):388-392.
[16]Ocker M,Schneider-Stock R.Histone deacetylase inhibitors:signalling towards p21cip1/waf1[J].Int J Biochem Cell Biology,2007,39(7-8):1367-1374.
[17]Li W,Sanki A,Karim RZ,et al.The role of cell cycle regulatory proteins in the pathogenesis of melanoma[J].Pathology,2006,38(4):287-301.
[18]Bartek J,Bartkova J,Lukas J.The retinoblastoma protein pathway and the restriction point[J].Curr Opin Cell Biol,1996,8(6):805-814.
[19]Parry D,Mahony D,Wills K,et al.Cyclin DCDK subunit arrangement is dependent on the availability of competing INK4 and p21 class inhibitors[J].Mol Cell Biol,1999,19(3):1775-1783.
[20]Isaacs JT,Isaacs WB.Androgen receptor outwits prostate cancer drugs[J].Nat Med,2004,10(1):26-27.
[21]Loriot Y,Zouheidi A,Gleave ME.Targeted therapies in m etastatic castration-resistant prostate cancer:beyond the androgen receptor[J].Urol Clin North Am,2012,39(4):517-531.
[22]Kahl P,Gullotti I,Heukamp LC,et al.Androgen receptor coactivators lysine-specific histone demethylase 1 and four and a half IAM domain protein 2 predict risk of prostate cancer recurrence[J].Cancer Res,2006,66(23):11341-11347.
