丹麦钢头鳟染色体核型分析
2013-10-13李秉钧吴志昊
李秉钧, 吴志昊, 尤 锋, 王 波, 徐 彬
(1. 烟台大学 海洋学院, 山东 烟台264005; 2. 中国科学院 海洋研究所, 山东 青岛 266071; 3. 国家海洋局第一海洋研究所, 山东 青岛 266061)
钢头鳟(Oncorhynchus mykiss)俗称钢头三文鱼、丹虹三文鱼、钢头、钢头鲑、三文鳟等, 属于鲑形目(Salmoniformes)、鲑科(Salmonidae)、鲑亚科(Salmoniae)、大麻哈鱼属(Oncorhynchus)。与虹鳟为同一种的不同品系, 其生活史的大部分时间在大洋或大湖区度过, 具有洄游性, 繁殖时溯河产卵; 而虹鳟为陆封型的, 不需要在不同水体间进行长距离洄游[1]。钢头鳟经过驯化也可以完全在淡水中生长繁殖,无需洄游[2]。从外部形态看, 钢头鳟沿体侧侧线下半部为银白色, 上半部为暗绿色, 头背部则为铁灰色,身上斑点大而稀疏[3]; 而虹鳟成体沿体侧侧线有一明显的棕红色纵纹, 如同彩虹一样, 也因此得名“虹鳟”。钢头鳟的肉质爽滑且结实, 肉呈鲜橙色, 可做成与极品鲑鱼相媲美的生鱼片, 经济价值很高。在加拿大、美国、丹麦、英国、智利等国家都已经开展了大规模养殖[4-7]。北京康鑫水产公司1998年从美国将类似种硬头鳟引进中国, 并在北京密云水库成功地进行了培育[8]。由于其养殖业经济效益突出, 近几年在中国各省逐步推广, 也开展了苗种培育技术、重金属的急性毒性实验、细胞体外培养、三倍体诱导等相关的研究[3,9-11]。作者所分析的钢头鳟是丹麦降海型 DK新品系之一, 它与国内以前引进并已开展大规模养殖的陆封型虹鳟品种, 如金鳟、硬头鳟、道纳尔逊优质虹鳟等均属于同一物种虹鳟的不同品系, 具有耐高温、易海水驯化、速生、抗病、高产等特点, 因而更具广阔的养殖前景。
染色体作为生物体中遗传物质的载体, 能在一定程度上反映物种的分化与形成过程、亲缘关系、演化途径及进化历史等特征[12]。所以, 有必要弄清钢头鳟的染色体核型和遗传背景, 为进一步开展该鱼种种质、遗传育种研究和养殖生产打下良好基础。鲑亚科鱼类共5属22种[13], 其染色体主要分两种类群, 一类二倍体染色体数接近 80, 臂数约为100(2n=80,NF=100); 另一类二倍体染色体数接近60,臂数也约为100(2n=60,NF=100)[14]。有关钢头鳟的核型研究报道不多, Colihueque等[6]曾对5个品系的虹鳟染色体数进行了统计, 发现硬头鳟的染色体数在58~61, 但并未进行染色体组型分析; 国内则仅有对其红血细胞核大小和形状变化的研究[15], 也未见到其染色体核型的报道。作者通过由丹麦引进的钢头鳟的染色体数目和核型研究, 以期获得其细胞遗传学基础参数。
1 材料与方法
1.1 实验材料
所用实验鱼耐高温钢头鳟由国家海洋局第一海洋研究所2011年从丹麦引进, 共10尾, 包括5尾雌鱼和5尾雄鱼, 体质量为90~150 g。
1.2 实验方法
1.2.1 染色体分裂相制备
采用稍加修改的PHA活体直接注射法制备染色体标本[16]。按 5 μg/g鱼体质量活体往腹腔内注射PHA, 20 h后按2.5 μg/g鱼体质量注射秋水仙素溶液,3 h后断尾放血, 解剖取其头肾并用生理盐水冲洗2次,充分剪碎后, 用 200目尼龙网过滤细胞悬液至刻度离心管中, 经低渗、固定、滴片、自然干燥、Giemsa染色等处理后, 光学显微镜下观察、计数、测量和拍照。
1.2.2 核型分析
核型分析选取实验鱼的染色体中期分裂相50个进行染色体计数(3~6个/尾实验鱼), 并选出 10个分散良好的中期分裂相进行染色体配对和长度测量,根据Levan等[17]的标准进行核型分析。染色体相对长度计算公式; 染色体相对长度=(实测单条长度×2/全部染色体长度总和)×100。利用测得的染色体相对长度, 在Excel中绘制实验鱼的核型模式图[18]。
2 结果
2.1 染色体数目的确定
在显微镜下对钢头鳟中期分裂相进行计数,共计数50个分裂相, 其染色体总数为58, 即2n=58(表1)。雌鱼和雄鱼染色体数目无差异, 也未观察到异形染色体。

表1 钢头鳟染色体数目统计结果Tab. 1 Chromosome numbers of Domsea steelhead trout O.mykiss
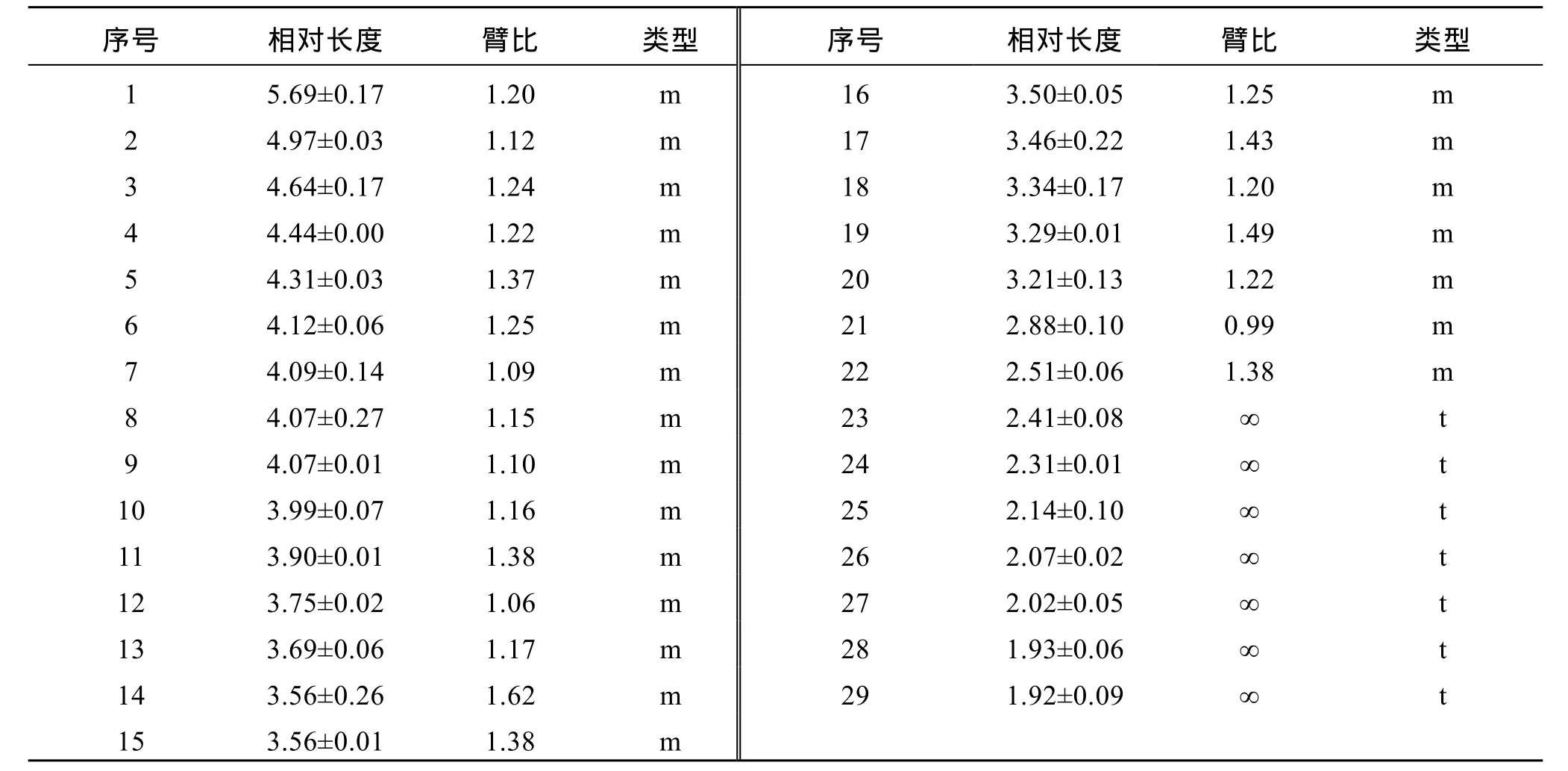
表2 钢头鳟染色体相对长度和臂比值Tab. 2 Relative length and ratio of chromosomes of Domsea steelhead trout O. mykiss
2.2 染色体相对长度的分析
对实验鱼的10个分散良好的中期分裂相进行显微摄像, 统计获得的染色体的相对长度和臂比见表 2,实验鱼中期染色体相对长度最长为(5.69±0.17), 最短为(1.92±0.09)。染色体配对和核型分析如图1, 染色体组型为 2n=44m+14t, 臂数NF=102。对实验鱼的 29对同源染色体按相对长度递减趋势进行编号, 获得了染色体核型模式图(图2)。

图1 钢头鳟染色体中期分裂相(左)和核型(右)Fig.1 Metaphase chromosomes (left)and karyotype (right)of Domsea steelhead trout O. mykiss
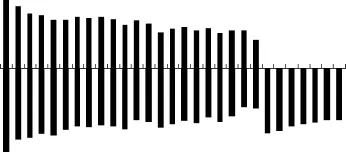
图2 钢头鳟的核型模式图Fig.2 The schema chart of chromosome of Domsea steelhead trout O. mykiss
3 讨论
染色体是遗传物质的主要载体, 特定的生物体具有特定数目、形态特征的染色体, 一般认为鱼类染色体的核型结构与其演化程度是一致的[12]。小岛吉雄[19]按染色体形态将真骨鱼类划分为低位类、中位类和高位类3个演化类群。高位类群的鱼类染色体的特点是其数目分布呈收敛状态, 较集中, 峰值是2n=48,M型染色体(包括m和sm染色体)少, 平均为7.5; A型染色体(包括st和t染色体)多, 平均为38.3, NF平均值为53.4; 而低位类群的鱼类染色体的数目相对较多, M型染色体多而A型染色体少。鲑科鱼类属典型的冷水性鱼类, 约有36种, 是世界三大养殖鱼类之一, 其养殖产量仅在鲤鱼和罗非鱼之后[20]。国内外许多研究均认为鲑形目鱼类是多倍体起源的鱼类, 在进化过程中经历了四倍化的进程[21]。目前已有染色体核型相关报道的鲑科鱼类中, 染色体数总体变化较大(表3)。其中白点鲑(Salvelinus pluvius)、哲罗鲑(Hucho taimen)和高白鲑(Coregonus peled)染色体数较多, 属于低位类群; 孟苏大马哈鱼(Oncorhynchus masu)、虹鳟和钢头鳟虽然染色体数略少于以上3种鱼, 但其M型染色体相对较多(超过70%), 其演化类群也趋向低位类群; 大西洋鲑(Salmo salar)的染色体数为56, M型染色体相对较少, 仅占32%, 其演化类群相对其他鲑科鱼类趋向高位类群。但其染色体数与高位类群鱼类染色体数的峰值48相比仍不够收敛, 并不能将其划分为高位类群。因此, 无法将鲑科鱼类在鱼类系统进化上统一归于某一位类群。李树深[22]研究认为,在特定的分类阶元中, 具有较多端部着丝粒染色体的为原始类群; 而具有较多中部或亚中部着丝粒染色体的为特化类群, 其中臂数少的为较特化类群,臂数多的为特化类群。在已报道的鲑科鱼类中(表3),染色体核型表现出高度的多样性, 进化并不保守。大西洋鲑具有较多端部着丝粒染色体, 属于较原始的类群。而虹鳟和钢头鳟具有较多中部或亚中部着丝粒, 属于特化类群。这种染色体数目和形态的差异可能是不同物种由于隔离造成的长期的基因交流缺失导致的。Philips等[14]在对鲑科鱼类的染色体进化研究表明, 鱼类栖息的环境对其染色体多态性有很大影响。
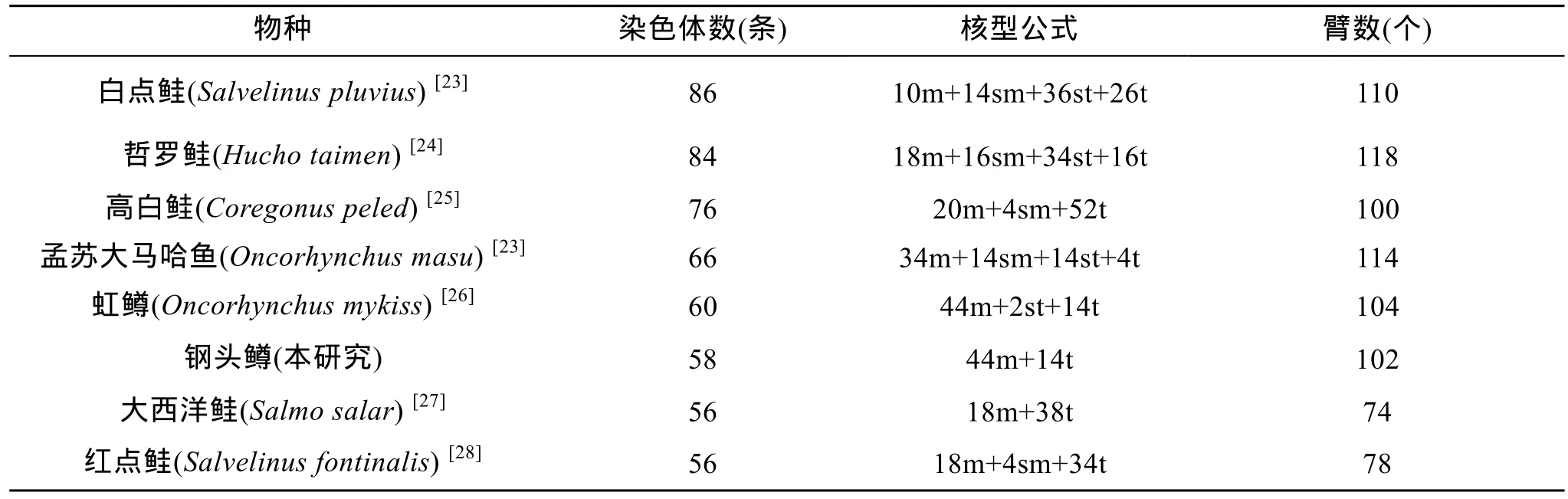
表3 几种鲑科鱼类的染色体核型Tab. 3 Chromosomal numbers and karyotypes of some species of Salmonidae
作者研究的钢头鳟为丹麦学者经过几十年选育出来的降海型新品系。不同品系的虹鳟除在外形上略有差异外, 在染色体核型上也存在一定差异。对于不同品系虹鳟染色体数目的研究已有较多报道,1955年Wright[29]首次利用囊胚细胞研究了虹鳟的染色体, 显示其染色体数为 60。之后又有许多学者研究了不同虹鳟地理群体或品系的核型, 得到不尽相同的结果。Minciu等[30]对采自罗马尼亚的两个虹鳟群体染色体的研究发现其数目在群体间存在差异,有 2n=60和 2n=61两个群体。Vasilyev等[31]发现勘察加半岛的虹鳟染色体数目为 2n=62。Thorgaard[32]分析了从阿拉斯加到加利福尼亚的 29个虹鳟群体,发现其染色体数目从 58~64不等。Colihueque等[6]对包括硬头鳟在内的 5个虹鳟品系染色体数进行了统计, 发现它们的染色体数在 58~63, 其中硬头鳟的染色体数在58~61。作者所分析的钢头鳟染色体数目为 58。这种不同品系间染色体数目的差异可能与罗伯逊易位(Robertsonian translocation)有关。罗伯逊易位现象是指同一细胞核内两条端部着丝点染色体在着丝点位置融合为一条中部或亚中部着丝点染色体,以及由一条中部或亚中部着丝点染色体在着丝点处断裂为两条端部着丝点染色体的现象, 这一变化或易位是正反两方向变化的。从物种进化看, 罗伯逊易位会导致种内出现染色体核型的多样性, 再经过一定的地理隔离, 这些具有不同染色体核型的群体间经过独立的进化, 进而衍生出亚种, 乃至新物种[33]。
已有文献报道[26], 虹鳟属雌性同配雄性异配型XX-XY, 但是否有异形性染色体还缺乏足够的证据,目前大多研究报道中均未发现性染色体, 仅Thorgaard[34]发现虹鳟雄性个体的亚端部着丝点染色体在短臂存在异形现象, 并认为其为虹鳟性染色体分化的早期阶段。本研究中同时对雄性和雌性刚头鳟的染色体分裂相进行了观察, 也未发现异形染色体。
[1]杨贵强, 徐绍刚, 王跃智,等. 硬头鳟幼鱼部分形态性状和体重的关系[J]. 动物学杂志, 2011, 46(1): 16-22.
[2]Neal G A, Edward R S, Doran M M. Diet, feeding rate,growth, mortality, and production of juvenile steelhead in a Lake Michigan Tributary[J]. North American Journal of Fisheries Management, 2007, 27: 578-592.
[3]徐绍刚, 田照辉, 杨贵强. 硬头鳟苗种培育技术[J].中国水产, 2009, 4: 47-48.
[4]Palstra P F, O’Connell F M, Ruzzante E D. Population structure and gene flow reversals in Atlantic salmon(Salmo salar)over contemporary and long-term temporal scales: effects of population size and life history[J]. Molecular Ecology, 2007, 16: 4504-4522.
[5]Fleming A I, Jonsson B, Gross R M. Phenotypic divergence of sea-ranched, farmed, and wild salmon[J].Canadian Journal of Fisheries and Aquatic Sciences,1994, 51: 2808-2824.
[6]Colihueque N, Iturra P, Estay F, et al.Diploid chromosome number variations and sex chromosome polymorphism in five cultured strains of rainbow trout(Oncorhynchus mykiss)[J]. Aquaculture, 2001, 15:63-77.
[7]Volpe1 P J, Taylor B E, Rimmer W D, et al. Evidence of natural reproduction of aquaculture-escaped Atlantic salmon in a coastal British columbia river[J]. Conservation Biology, 2000, 14: 899-903.
[8]刘澧津. 硬头鳟(Oncorhynchus mykiss)苗种养殖试验[J]. 水产学杂志, 2001, 14(2): 72-74.
[9]童裳亮. 硬头鳟巨噬细胞的体外长期培养[J]. 实验生物学报, 22(2): 241-245.
[10]蔡国雄, 伊格·所罗, 爱德华·唐纳森. 热休克和水静压刺激诱导三倍体硬头鳟(Oncorhynchus mykiss)对比研究[J]. 水产学杂志, 1990, 3(2): 31-352.
[11]姚再旺. 日本研究锌对硬头鳟的急性毒性[J]. 水产科技情报, 1984 (1): 26.
[12]余先觉, 周暾, 李渝成. 中国淡水鱼类染色体[M].北京: 科学出版社, 1989.
[13]孟庆闻, 苏锦祥, 缪学祖. 鱼类分类学[M]. 北京: 科学出版社, 1994, 309-318.
[14]Philips R, Rab P. Chromosome evolution in the Salmonidae(Pisces): an update[J]. Biological Reviews,2001, 7: l-25.
[15]蔡国雄. 硬头鳟(Oncorhynchus mykiss)早期发育过程中红血细胞核的尺度变化和类型转换[J]. 水产学杂志1996, 9(1): 1-6.
[16]林义浩.快速获得大量鱼类肾细胞中期分裂相的 PHA体内注射法[J]. 水产学报, 1982, 6(3): 201-208.
[17]Levan A, Fredga K, Sandberg A A. Nomenclature for centromeric position on chromosomes[J]. Hereditas,1964, 52(2): 201-220.
[18]乔永刚, 宋芸. 利用 EXCLE制作核型模式图[J]. 农业网络信息, 10: 97-98.
[19]小岛吉雄. 鱼类细胞遗传学[M]. 广州: 广东科技出版社, 1985.
[20]孙大江, 王炳谦. 鲑科鱼类及其养殖状况[J]. 水产学杂志, 2010, 23(2): 56-63.
[21]Ojima Y, Takai A. The occurrence of spontaneous polyloidy in the Japanese common loach,Misqurnus anguicandatus[J]. Proceedings of the Japan Academy,1979, 55: 487-491.
[22]李树深. 鱼类细胞分类学[J]. 生物科学动态, 1981, 2:8-15.
[23]王清印, 余来宁, 杨宁生. 中国水产生物种质资源与利用(第1 卷)[M]. 北京:海洋出版社, 2005, 47-55.
[24]薛淑群, 尹洪滨, 尹家胜. 哲罗鱼(Hucho taimen)染色体组型与 DNA 含量分析[J]. 广东海洋大学学报,2010, 30(3): 6-10.
[25]薛淑群, 孙中武, 尹洪滨. 高白鲑的细胞遗传学分析[J]. 水生生物学报, 2011, 35(5): 869-873.
[26]宋苏祥, 孙大江, 刘洪柏. 虹鳟鱼核型及 DNA 含量分析[J]. 水产学杂志, 1995, 8 (1): 27-31.
[27]尹洪滨, 牟振波, 孟繁华. 大西洋鲑的染色体核型[J].东北林业大学学报, 2003, 31(1): 27-28.
[28]虎永彪, 张艳萍, 娄忠玉, 等. 美洲红点鲑染色体核型分析[J]. 水产科技情报, 2010, 37(3): 154-156.
[29]Wright E J. Chromosome numbers in trout[J]. The Progressive Fish-Culturist, 1955, 17(4): 172-176.
[30]Minciu R D, Bara I. A study of mitotic chromosomes of two populations ofSalmo gairdneri[J].Rich Revue Roumaine de Biology, 1986, 3l(1): 55-59.
[31]Vasilyev P V. Karyotypes of different forms of the kamchatka trout,Salmo mykissand the rainbow trout,Salmo gairdneri[J].Ichology, 1975, 15: 889-900.
[32]Thorgaard H G. Chromosomal differences among rainbow trout populations[J].Copeia, 1983, 3: 650-662.
[33]朱必才, 高焕, 屈艾,等.罗伯逊易位研究的进展[J].遗传, 2001, 23(6): 599-604.
[34]Thorgaard H G. Heteromorphic sex chromosomes in male rainbow trout[J].Science, 1977, 196: 900-902.
