广东省猪链球菌2型菌株毒力基因及分子分型分析
2013-09-26陈经雕刘美真柯碧霞谭海玲李柏生柯昌文
陈经雕,刘美真,柯碧霞,谭海玲,李柏生,柯昌文
猪链球菌(Streptococcus suis)是一种能在猪群中正常携带,偶尔导致猪、人发病的人兽共患的动物源性微生物。根据其荚膜多糖抗原的异同划分为35个血清型,其中猪链球菌2型为优势致病血清型,而不同猪链球菌2型菌株间的致病力又存在明显的差异,这种差异可能与各菌株所携带的毒力因子有很大关系[1]。目前认为猪链球菌2型的毒力因子主要包括荚膜多糖(cps2J)、溶菌酶释放蛋白(mrp)、溶血素(sly)、谷氨酸脱氢酶(gdh)、胞外因子(ef)、纤连蛋白/血纤蛋白原结合蛋白(FBPS)和毒力相关序列ORF2等[2]。为了解广东地区猪链球菌2型菌株毒力基因分布及分子分型情况,本研究选取了其中5个毒力基因对我省分离自病人和病猪的猪链球菌2型菌株进行PCR检测分析,并对菌株进行MLST分子分型,为诊断、治疗和制定猪链球菌疫情防控措施提供科学依据。
1 材料与方法
1.1 实验菌株 2005-2008年共22株,其中17株来自病人(其中2005年5株分别来自潮州、阳江、深圳、韶关和佛山,2006年5株分别来自江门、广州、佛山、中山和惠州,2007年7株分别来自佛山、江门、深圳各2株和广州1株),5株来自病猪(均为2008年分别来自肇庆2株、河源、茂名、惠州各1株),阳性对照的猪链球菌2型标准株SS2(来自德国)核酸由江苏省疾控中心惠赠。
1.2 试剂来源 诊断血清购自丹麦(STATENS SERUM INSTITUT)公司,API 20Strep生化鉴定试验条购自法国梅里埃公司,分子生物学试剂购自TaKaRa宝生物工程(大连)有限公司,引物由上海生工公司合成,均在有效期内使用。测序反应用PRISM BigDye Terminator Cycle Sequencing Kit(v3.1,ABI)试剂盒,操作按试剂盒说明书进行。
1.3 实验方法
1.3.1 菌株鉴定[3]从菌落形态、血清学实验及API 20Strep生化鉴定分别对菌株进行种、型鉴定。1.3.2 毒力因子检测[3]
1.3.2.1 菌株DNA提取 用煮沸法或细菌基因组DNA试剂盒提取所有菌株的基因组DNA,试剂盒提取可参考说明书操作。
1.3.2.2 引物 见表1。

表1 5个毒力基因的引物序列及产物大小Tab.1 Five virulence genes’primer sequences and product sizes
1.3.2.3 PCR反应体系 反应体系为25μL,10×PCR Premix缓冲液12.5μL,上、下游引物(10 mmol/L)各1μL,模板2μL,用去离子水调整终体积至25μL。反应结束后各取5μL于1% 琼脂糖凝胶电泳,用凝胶成像系统拍照。
1.4.1 多位点序列分析(MLST)
1.4.1.1 提取细菌基因组DNA 按照细菌DNA提取试剂盒说明书严格操作,提取的基因组-20℃保存待用。
1.4.1.2 PCR 扩增、测序、数据分析 参照文献[4-5]分 别 用 PCR 扩 增 aroA、cpn60、dpr、gki、mutS、recA和thrA7个管家基因DNA片段,PCR产物用1%琼脂糖凝胶电泳后测序。各管家基因序列经DNA star SeqMan软件处理后与数据库http://ssuis.mlst.net/进行比对,获得各位点的等位基因数值,并形成相应的菌株等位基因谱,提交MLST 网站,确定序列分型(Sequence Type,ST)。
2 结 果
2.1 菌株鉴定 对22株菌株进行菌落形态、血清学实验及API 20Strep生化鉴定,全部菌株均为猪链球菌2型。
2.2 毒力基因检测 22株菌分为5种毒力基因型,其中病人及病猪菌株均有cps2J+/mrp+/sly+/gdh+/ef+型别(病人7株、病猪3株),cps2J+/mrp+/sly-/gdh-/ef+ (6 株)、cps2J+/mrp+/sly-/gdh+/ef+(4株)为病人分离株所特有,cps2J+/mrp-/sly+/gdh+/ef+(1株)、cps2J+/mrp-/sly-/gdh+/ef-(1株)为病猪分离株所特有,图1为部分菌株的cps2J基因片段结果。结果见图1、表2。
2.3 MLST分型 对所有菌株的7个管家基因片段进行扩增,经测序、比对分析显示,均能够在数据库找到对应的基因型,共有3个基因型,分别为ST1型(病人9株、病猪2株)、ST7型(病人8株、病猪2株)和ST28(病猪1株)型,其中ST1、ST7型菌株亲缘关系极近,均同属于ST1克隆复合物。结果见图2。
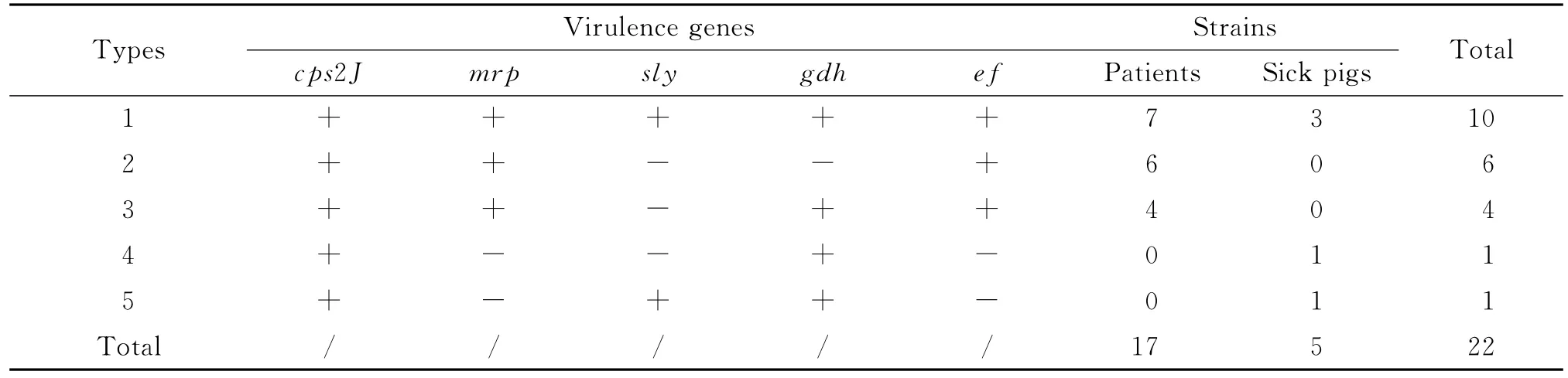
表2 广东地区猪链球菌2型的毒力基因类型Tab.2 Detection of virulence genes in Ss2in Guangdong
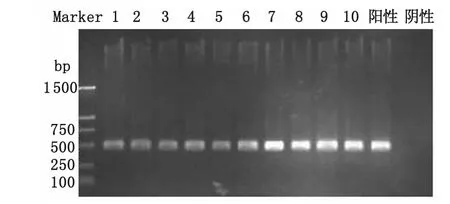
图1 部分菌株夹膜多糖基因片段(cps2J)结果 -450bpFig.1 Strains capsular polysaccharide gene fragment(cps2J)-450bpThe 1-7are part of strains from patients;8-10 are part of isolates from sick pigs
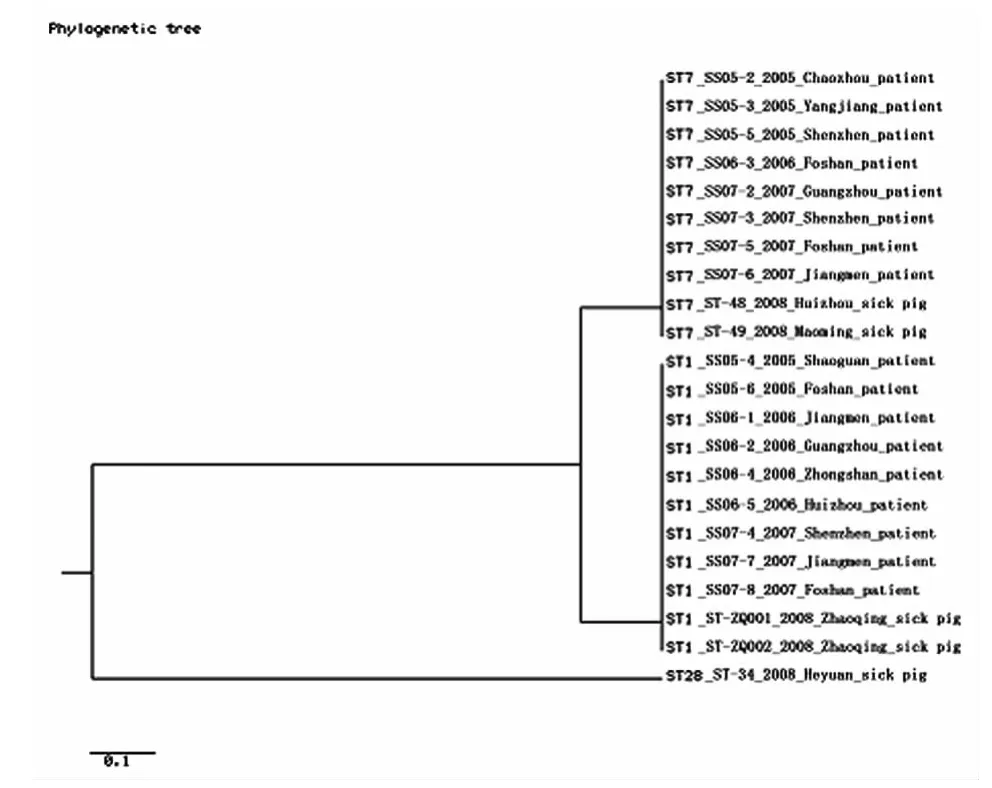
图2 广东地区猪链球菌2型菌株MLST分型聚类图Fig.2 MLST type cluster map of Streptococcus suis type 2in Guangdong Province
3 讨 论
猪链球菌2型菌株曾引起我国1998年江苏省和2005年四川省的部分地区的人感染猪链球菌病疫情的暴发流行。广东省猪链球菌在猪间的流行早有报告,但从病原学上证实人感染猪链球菌2型尚属首次[6],且与江苏、四川人感染猪链球菌病疫情暴发不同,广东地区的猪链球菌2型菌株均来源于散发的病例和病猪。
研究发现并不是所有的猪链球菌2型菌株都具有致病性,其致病力取决于它的毒力因子,根据菌株毒力因子的差异,将其分为强毒力株、弱毒力株和无毒力株,其致病机理目前还不很清楚,但普遍认为其致病力的差异与该菌株的毒力因子分布有着密切的关系[7]。Cps2J是猪链球菌2型菌株抵抗巨噬细胞吞噬的重要物质,在猪链球菌致病过程中有重要作用,也是猪链球菌进行分型的标志。Smith等[8]研究发现通过插入突变得到的无荚膜的等位基因突变株在体外很容易被猪肺泡巨噬细胞摄入,更重要的是,通过鼻腔接种无菌仔猪的突变株对其完全无致病性,由此可见,荚膜多糖与菌株的致病力有关。
mrp与ef也是猪链球菌2型菌株重要的毒力因子,曾巧英等[9]研究证明mrp可单独作为猪链球菌2型菌株的毒力因子。猪链球菌血清型众多,ef与血清型有相关性,ef是一种粘附素,能黏附上皮细胞并诱导上皮细胞融合和凋亡[10]。Henk J等[11]研究欧洲的猪链球菌菌株结果显示:猪链球菌血清1型、2型、1/2型和14型菌株表达ef蛋白的百分率很高,菌株的毒力可能与ef相关。本研究中除了1株来自病猪的菌株为ef阴性外,其余菌株均为ef阳性。由于mrp和ef在强毒力菌株中检出率很高,在无毒力菌株中检出率很低,因而携带mrp和ef常被认为是猪链球菌2型高致病性的标志[12]。Vecht U[13]等研究发现mrp+、epf+ 的表现型菌株感染小猪时可以引起典型的脑炎、多发性浆液炎及多发性关节炎,而mrp-、ef-的表现型菌株对猪则没有致病力。本研究中的菌株mrp+、epf+ 高达90.9%(20/22),其中1株来自病猪的菌株为mrp-、ef-,这可能是由别的毒力基因所导致发病。
Sly对Hep22细胞、内皮细胞和Vero细胞均具有一定的毒性作用,且经纯化后溶血素免疫的小鼠和猪均能耐受强毒力株的攻击,所以推测其可能也是猪链球菌2型的一种重要的致病因子,也是一种保护性抗原[14]。然而,猪链球菌2型的溶血素基因分布具有很强的地域性,来自欧洲的大多数菌株能产生溶血素,而来自北美的菌株不产生溶血素,但仍然具有致病性[1]。本研究中的菌株sly+ 只有50.0%(11/22)。据相关研究证实基于gdh蛋白的检测可区分猪链球菌2型的强毒力株、弱毒力株和无毒力株,同时,gdh也与猪链球菌的种属有关[15]。
本研究通过对广东地区病人、病猪的猪链球菌2型分离株进行上述5种主要毒力因子的PCR检测,研究发现22株菌株包含了5种毒力基因型,其中cps2J+/mrp+/sly+/gdh+/ef+型别为病人及病猪菌株所共有,共占45.5%(10/22),cps2J+/mrp+/sly-/gdh-/ef+、cps2J+/mrp+/sly-/gdh+/ef+为病人菌株所特有,各占27.3%(6/22)、18.2%(4/22),cps2J+/mrp-/sly+/gdh+/ef+、cps2J+/mrp-/sly-/gdh+/ef-为病猪菌株所特有,各占4.5%(1/22)、4.5%(1/22),这就提示我省目前猪链球菌2型流行菌株的毒力基因型主要为同时具有上述5种毒力基因的基因型,但低于赵冉等[16]研究的5种毒力基因96% 均为阳性的结果。本次研究的病例中有两例(毒力基因型别分别为:cps2J+/mrp+/sly+/gdh+/ef+、cps2J+/mrp+/sly-/gdh-/ef+)死亡,共占本次研究的毒力基因型别的72.8%(16/22)。但由于分离的菌株数量较少,且收集到病例、病猪的临床表现和病理改变资料也有限,因此广东地区的猪链球菌2型分离株是否还有更多的毒力基因型,各种毒力基因型的菌株与临床病人或病猪的临床表现和病理改变有何关联等,还有待对更多的猪链球菌2型分离株进行检测研究。本研究结果表明广东地区猪链球菌2型菌株毒力基因型呈现多样化,部分毒力基因型为病人及病猪所共有,cps2J+/mrp+/sly+/gdh+/ef+型别是目前广东地区猪链球菌2型菌株毒力基因型别的优势型别。
多位点序列分型技术(multilocus sequence typing,MLST)是基于PCR扩增、基因测序的基因分型方法。MLST方法具有很高的分辨能力,不需用参考菌株对实验室间和实验室内的结果进行标准化,适用于流行病学研究[17]。并且可以通过互联网进行快速的数据共享,有利于全球范围的流行病学的比较与分析。King S J等[18]对英国294株猪链球菌分离株进行了MLST分析,共获得了92个序列型(ST),其中ST1克隆复合物是最主要的ST分组,它包含了全世界各地的绝大多数的高致病性分离株,ST1型多来自脑膜炎、关节炎和败血症的样本,ST27、ST87型则主要来自肺脏,与临床背景相关良好。Princivalli M S等[19]对意大利2003-2007年分离的菌株研究时也发现ST1型主导着这几年的流行,并且大都携带mrp、ef和sly基因。与欧洲报道相比,我国分离株则以ST7型为主,其次是ST1型。叶长芸等对国内两次暴发(1998年江苏、2005年四川)的猪链球菌进行研究,发现114株猪链球菌中有106株属于ST7型。王洪敏等[20]的研究表明广东2005年的5株人源猪链球菌2型株菌中有3株属于ST7型,2株属于ST1型,其中ST7型与2005年四川疫情株相同,而病人的ST1型与香港地区发现的猪链球菌相同;这与四川省暴发的人间猪链疫情明显不同,四川仅由单一的ST7型猪链球菌克隆引起。研究表明,ST7型刺激T细胞增殖和诱导致炎因子释放的能力比欧洲流行株ST1型更强[21]。不难看出,MLST最大特点是不同的实验室获得的结果可以比较,易于统一、标准化,并可经互联网共享,现已有许多学者将该技术用于流行病学研究中[22]。
此研究中对分离到的猪链球菌2型菌株进行MLST分析发现22株菌分布在3个ST型别:11株(病人9株、病猪2株)属于ST1型,10株(病人8株、病猪2株)属于ST7型,1株来自病猪的猪链球菌2型属于ST28型,其中ST1、ST7型为病人和病猪菌株所共有,均同属于ST1克隆复合物,显示两者菌株间的亲缘关系极近。ST28型为病猪所特有,病猪菌株的ST型别呈现多样化,部分型别与病人菌株出现交叉。
[1]Lu LL,Li R,Zheng YL,etal.Virulence factors of Streptococcus suis type 2[J].Chin J Health Lab Technol,2008,18(3):570-572.(in Chinese)路玲玲,李蓉,郑玉玲,等.2型猪链球菌毒力因子研究进展[J].中国卫生检验杂志,2008,18(3):570-572.
[2]Fang QM,Huang SQ,Yu FS,etal.Genetic analysis on the main virulence genes in the Fujian strain of Streptococcus suis type 2from slaughtered pigs[J].Chin J Zoonoses,2010,26(1):65-68.(in Chinese)方勤美,黄绍谦,俞伏松,等.猪链球菌2型福建分离株的主要毒力基因分析[J].中国人兽共患病学报,2010,26(1):65-68.
[3]GB/T 19915.2-2005.Methods for detection of Streptococcus suis type 2[S].Beijing:General Administration of Quality Supervision,Inspection and Quarantine of the People's Republic of China,Standardization Administration of the People's Republic of China,2005.(in Chinese)GB/T 19915.2-2005,猪链球菌2型分离鉴定操作规程[S].北京:中华人民共和国国家质量监督检验检疫总局、中国国家标准化管理委员会,2005.
[4]Samantha JK,James AL,Peter JH,etal.Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis:identification of virulent clones and potential capsular serotype exchange[J].J Clin Microbiol,2002,40(10):3671-3680.DOI:10.1128/JCM.40.10.3671-3680.2002
[5]Hill JE,Gottschalk M,Brousseau R,etal.Biochemical analysis,cpn 60and 16SrDNA sequence data indicate that Streptococcus suis serotypes 32and 34,isolated from pigs,are Streptococcus orisratti[J].Vet Microbiol,2005,107(1-2):63-69.
[6]He JF,Kang M,Chen JD,etal.The epidemiological characteristics of cases of Streptococcus suis infection five cases of Guangdong Province[J].Chin J Epidemiol,2006,10(10):864.(in Chinese)何剑峰,康敏,陈经雕,等.广东省五例人感染猪链球菌病例流行特征分析[J].中华流行病学杂志,2006,10(10):864.
[7]Staats JJ,Feder I,Okwumabua O,etal.Streptococcus suis:past and present[J].Vet Res Commun,1997,21:381-407.DOI:10.1023/A:1005870317757
[8]Smith HE,Damman M,Van der Velde J,etal.Identification and characterization of the cpslocus of Streptococcus suis serotype 2:the capsule protects against phagocytosis and is an important virulence factor[J].Infect Immun,1999,67(4):1750-1756.
[9]Zeng QY,Lu CP.Induced cell fusion and apoptosis of epithelial muramidase-released protein of Streptococcus suis type 2[J].Acta Microbiologica Sinica,2003,43(3):407-411.(in Chinese)曾巧英,陆承平.猪链球菌2型溶菌酶释放蛋白诱导上皮细胞融合和凋亡[J].微生物学报,2003,43(3):407-411.
[10]Gao SQ,Lei LC,Han WY.Progress on major virulence factors of Streptococcus suis type 2[J].Chin J Biologicals,2006,19(6):657-659.(in Chinese)高尚庆,雷连成,韩文瑜.猪链球菌2型主要毒力因子研究进展[J].中国生物制品学杂志,2006,19(6):657-659.
[11]Wisselink HJ,Smith HE,Stockhofe-Zurwieden N.etal.Distribution of capsular types and production of muramidase reaseased protein (MRP)and extracellular factor (EF)of Streptococcus suis strains isolated from diseased pigs seven European countries[J].Vet Microbiol,2000,74(3):237-248.
[12]Norton PM,Rolph C,Ward PN,etal.Epithelial invasion and celllysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin[J].FEMS Immunol Med Microbiol,1999,26(1):25-35.
[13]Vecht U,Isselink HJ,Ellema MJ,etal.Identification of two proteins associated with virulence of Streptococcus suis type 2[J].Infect Immun,1991,59(9):3156-3162.
[14]Staats JJ,Plattner BL,Stewart GC,etal.Presence of the Streptococcus suis suilysin gene and expression of MRP and EF correlates with high virulence in Streptococcus suis type 2isolates[J].Vet Microbiol,1999,70(3-4):201-211.
[15]Okwumabua O,Persaud J,Reddy P.Cloning and characterization of the gene encoding the glutamate dehydrogenase of Streptococcus suis serotype 2[J].Clin Diagn Lab Immunol,2001,8(2):251-257.
[16]Zhao R,Sun JH,Lu CP.Distribution characteristics of the isolates of Streptococcus suis virulence factor[J].J Shanghai Jiaotong Univ(Agri Sci),2006,24(6):495-498.(in Chinese)赵冉,孙建和,陆承平.猪链球菌国内分离株毒力因子的分布特征[J].上海交通大学学报(农业科学版),2006,24(6):495-498.
[17]Liao YL,Zou QM.Progress in study of gene of pathogenic microorganisms of multilocus sequence typing[J].Prog in Microbiol Immunol,2007,5(4):65-68.(in Chinese)廖亚玲,邹全明.病原微生物基因多位点序列分型的研究进展[J].微生物免疫学进展,2007,5(4):65-68.
[18]King SJ,Leigh JA,Heath PJ,etal.Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis:identification of virulent clones and potential capsular serotype exchange[J].J Clin Microbiol,2002,40(10):3671-3680.DOI:10.1128/JCM.40.10.3671-3680.2002
[19]Princivalli MS,Palmieri C,Magi G.Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy(2003-2007)[J].Euro Surveill,2009,14(33):pii:19310.
[20]Wang HM,Ke CW,Pan WB,etal.MLST typing of Streptococcus suis isolated from clinical patients in Guangdong Province in 2005[J].J South Med Univ,2008,28(8):1438-1441,1445.(in Chinese)王洪敏,柯昌文,潘武滨,等.2005年广东省临床分离猪链球菌的 MLST分子分型研究[J].南方医科大学学报,2008,28(8):1438-1441,1445.
[21]Ye C,Zheng H,Zhang J,etal.Clinical,experimental,and genomic differences between intermediately pathogenic,highly pathogenic,and epidemic Streptococcus suis[J].Infect Dis,2009,199(1):97-107.DOI:10.1086/594370
[22]Ye CY,Bai XM,Zhang J,etal.Spread of Streptococcus suis sequence type 7in China[J].Emerg Infect Dis,2008,14(5):787-791.DOI:10.3201/eid1405.070437
