2008—2011年北京协和医院流感嗜血杆菌耐药性监测
2013-08-23杨启文
牛 翠,张 辉,杨启文,李 娜
2.邢台市第三医院检验科,河北邢台 054000。
流感嗜血杆菌(Haemophilus influenzae)是呼吸道感染的重要病原菌之一,可通过呼吸道在人与人之间传播。也可由鼻咽部侵入血流,导致多种感染性疾病如化脓性脑膜炎、小儿肺炎和血流感染、成人社区获得性肺炎、中耳炎、慢性支气管炎急性发作和急性鼻窦炎等。近年来由于抗菌药物的广泛应用,流感嗜血杆菌对抗菌药物的耐药率呈上升趋势,给临床抗感染治疗带来了许多困难。现对北京协和医院2008—2011年临床分离的流感嗜血杆菌的耐药情况作一分析,以指导临床采取有效的感染控制措施及合理用药,报道如下。
材料与方法
一、材料
(一)菌株来源 收集2008年1月至2011年12月北京协和医院临床标本分离的流感嗜血杆菌,剔除同一患者相同部位的重复分离菌株后共获223株。
(二)抗菌药物纸片 氨苄西林、氨苄西林-舒巴坦、阿莫西林-克拉维酸、头孢呋辛、头孢曲松、头孢噻肟、头孢克洛、氨曲南、环丙沙星、左氧氟沙星、莫西沙星、甲氧苄啶-磺胺甲口恶唑、氯霉素、阿奇霉素、四环素药敏纸片和V、X因子、V+X因子纸片均为英国OXOID公司产品。测定β内酰胺酶的头孢硝噻吩纸片为美国BBL公司产品。
(三)培养基 流感嗜血杆菌选择性培养基以哥伦比亚琼脂(OXOID公司产品)为基础制备巧克力平皿,浇注平皿前(约50℃)加入万古霉素(50 mg/L)。流感嗜血杆菌药敏培养基(HTM)和营养补充剂SR0158E为英国OXOID公司产品。
(四)VITEK2-NH 试卡 VITEK-2-NH 试卡由法国生物梅里埃公司提供。
二、方法
(一)菌株分离及鉴定 送检标本立即接种血平皿和加万古霉素巧克力平皿,置于35℃,5%~10%CO2培养24 h。挑取血平皿上不生长、巧克力平皿上生长的无色透明或灰白色半透明、圆形隆起、光滑、边缘整齐,湿润不溶血、露滴状菌落、并有特殊闪光,直径0.5~1 mm的可疑菌落涂片。革兰染色呈阴性短小杆菌、球杆菌、少数呈细丝状等多形态性。同时做卫星试验,将可疑菌落同时接种于血平皿、普通琼脂(MH)平皿上,密涂划线,再用金葡菌点种,两点相距2 mm以上。35℃,5%~10%CO2培养箱内培养24 h。血平皿卫星试验阳性、MH平皿卫星试验阴性者,鉴定为流感嗜血杆菌。同时进行V、X、X+V因子需求试验,只能在V+X纸片周围生长者,鉴定为流感嗜血杆菌。疑难菌株用法国生物梅里埃公司提供的NH卡最终鉴定。
(二)药敏试验 药敏试验采用纸片扩散法。以HTM琼脂为培养基,按美国CLSI标准判断结果。
(三)β内酰胺酶测定 用头孢硝噻酚显色反应法测定β内酰胺酶。
(四)质控菌株 以金葡菌ATCC25923为阴性对照,流感嗜血杆菌ATCC49247为阳性对照。
(五)统计学方法 采用WHONET5.6对数据进行统计学分析,P<0.05为差异有统计学意义。
结 果
一、流感嗜血杆菌的分布
(一)年龄分布 2008年1月1日至2011年12月31日共收集到223株流感嗜血杆菌,其中分离自成人(≥18岁)的菌株为208株,占93.3%;分离自未成年人(<18岁)的菌株为15株,占6.7% 。
(二)科室分布 223株流感嗜血杆菌分离自门诊101株(45.3%);分离自住院患者122株(54.7%),其中呼吸内科17株(7.6%),其他内科共90株(40.4%),外科10株(4.5%),儿科5株(2.2%)。
(三)标本分布 223株流感嗜血杆菌分离自痰液194株(87.0%),咽拭子、支气管肺泡灌洗液等呼吸道标本28株(12.6%),血培养1株(0.4%)。
二、223株流感嗜血杆菌对各种抗菌药物的耐药性
本次检测的流感嗜血杆对单环β内酰胺类(氨曲南),喹诺酮类(环丙沙星、左氧氟沙星、莫西沙星),加酶抑制剂(阿莫西林-克拉维酸、氨苄西林-舒巴坦),大环内酯类(阿奇霉素)抗菌药物的敏感率都在90.0%以上;对第二代头孢菌素(头孢呋辛、头孢克洛),第三代头孢菌素(头孢噻肟、头孢曲松)的敏感率在80.0%以上,而对氨苄西林、甲氧苄啶-磺胺甲口恶唑和四环素的敏感率较低,分别为78.1%、41.9%和34.3%。
三、连续4年流感嗜血杆菌对抗菌药物的敏感性分析
连续4年流感嗜血杆菌对大多数测试抗菌药物的耐药率差异无统计学意义,见表1。
四、连续4年分离自门诊和住院患者的流感嗜血杆菌对抗菌药物的敏感性分析
连续4年分离自门诊和住院患者的流感嗜血杆菌对大多数测试抗菌药物的耐药率差异无统计学意义,见表2。
五、β内酰胺酶检测结果
(一)β内酰胺酶阳性菌 223株流感嗜血杆菌中产β内酰胺酶阳性氨苄西林耐药菌株(BLPAR)为35株,产酶检出率为15.7%。
(二)β内酰胺酶阴性菌 β内酰胺酶阴性氨苄西林耐药菌株(BLNAR)为20株,占9.0%,见表3。
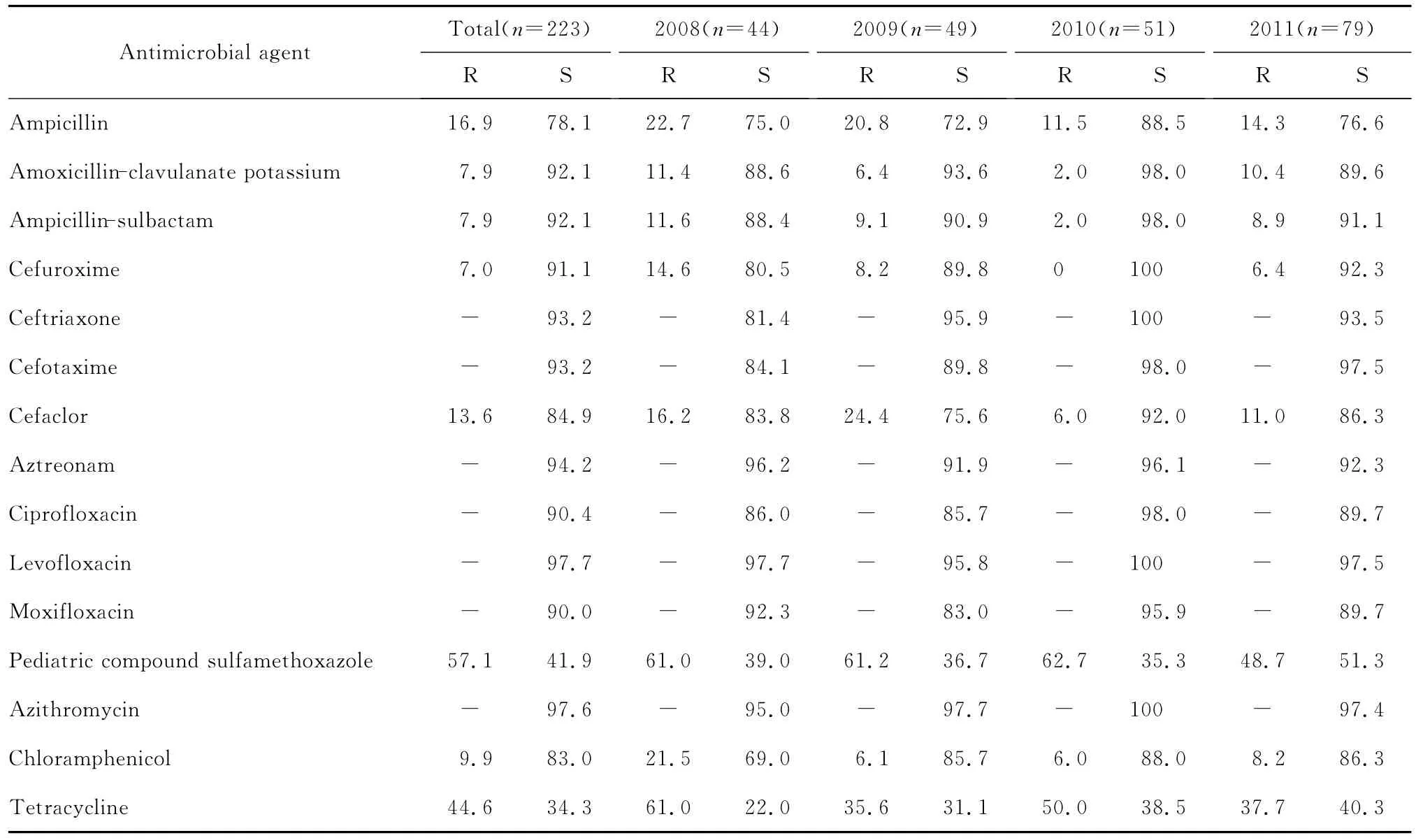
表1 2008—2011年流感嗜血杆菌对抗菌药物的耐药率和敏感率(%)。Table 1 The susceptibility of Haemophilus influenzae strains to various antimicrobial agents during 2008-2011(%)
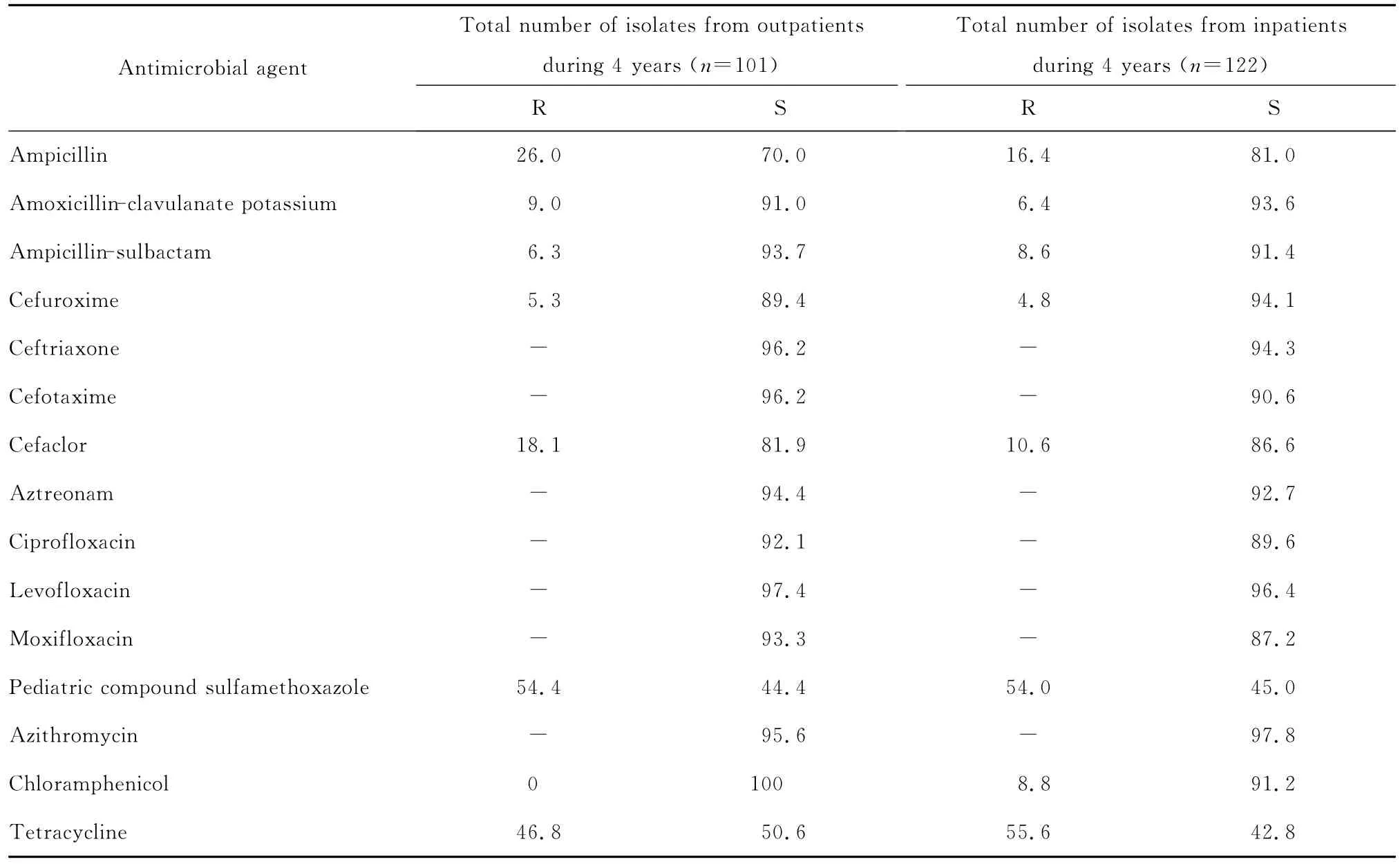
表2 4年间分离自门诊和住院患者的流感嗜血杆菌对抗菌药物的耐药率和敏感率(%)。Table 2 The suscetibility of Haemophilus influenzae isolated from outpatients and inpatients to antimicrobial agents during 4 consecutive years(%)
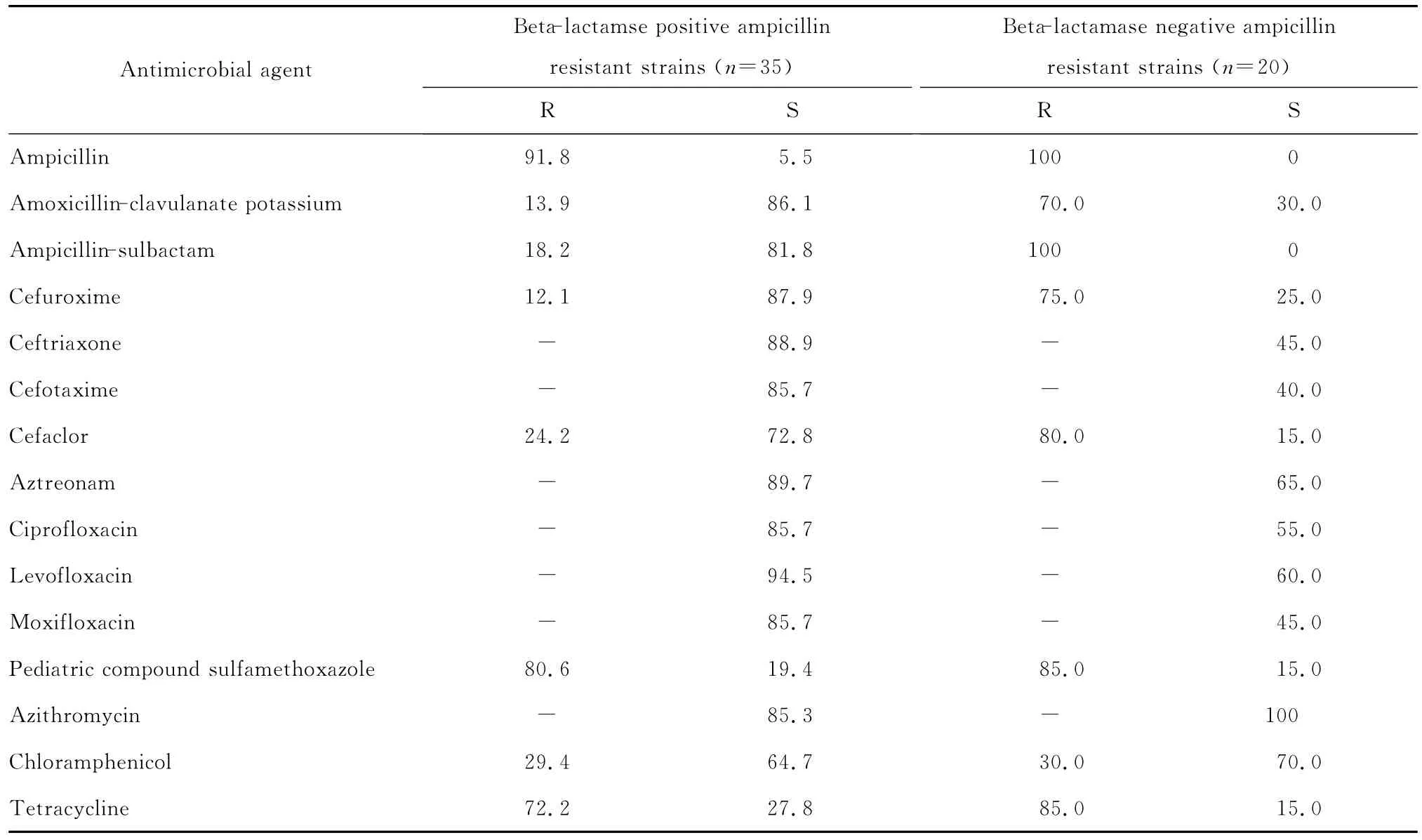
表3 4年间β内酰胺酶阳性和β内酰胺酶阴性氨苄西林耐药的流感嗜血杆菌对抗菌药物的耐药率和敏感率(%)Table 3 The susceptibility ofβ-lactamase-positive andβ-lactamase-negative ampicillin resistant Haemophilus influenzae strains to antimicrobial agents during 4 consecutive years(%)
讨 论
本次统计中流感嗜血杆菌对氨苄西林不敏感率为21.9%,4年间差异无统计学意义(P>0.05),略低于2010年中国CHINET细菌耐药监测数据(35.7%)[1]。对氨苄西林耐药的流感嗜血杆菌在1973年被首次报道,产β内酰胺酶是其主要耐药机制。编码β内酰胺酶的基因主要在质粒上,但也有些基因整合于细菌的染色体上,产生了2种β内酰胺酶:TEM-1和 ROB-1,以 TEM-1比较常见,而几乎所有产ROB-1酶的菌株出现在北美[2]。产β内酰胺酶的比率因地域分布而不同,如俄罗斯为4.2%,美国则为29.6%[3]。
β内酰胺酶阴性氨苄西林耐药,或者β内酰胺酶阳性,阿莫西林-克拉维酸耐药是由于青霉素结合蛋白(PBP)的改变,二型和未分型的菌株是由PBP3介导的,PBP3由ftsI基因整合[4]。根据2012 CLSI的规定,BLNAR应评价为对于这些菌株:阿莫西林-克拉维酸、氨苄西林-舒巴坦、头孢克洛、头孢他美、头孢尼西、头孢丙烯、头孢呋辛、氯碳头孢,头孢孟多以及哌拉西林-他唑巴坦均视为耐药,即使一些BLNAR株体外试验显示对这些药物敏感也应视为耐药。
超广谱β内酰胺酶(ESBL),是由TEM-1变异而来的,但是在流感嗜血杆菌中还未发现,尽管已经在克隆株上有表达[5],现已报道由2例南非患者体内分离的副流感嗜血杆菌是产ESBL的[6]。
流感嗜血杆菌对大环内酯类抗生素的耐药率在过去30年里一直没有改变,尽管已经发现了少量高耐药株[7-8]。
流感嗜血杆菌对喹诺酮类抗菌药物耐药非常罕见,广谱喹诺酮类抗菌药物通过干扰DNA复制而发挥抗菌作用,编码DNA解螺旋酶或者拓扑异构酶Ⅳ的耐药决定区的改变导致流感嗜血杆菌对喹诺酮药物耐药。喹诺酮类药物由于其不良反应目前不推荐给儿童使用。但是一项监测表明,一些喹诺酮类药物MIC值在增加,并且在一个长期护理的设备上发现了1株高耐药的克隆株[9]。相反,其对甲氧苄啶-磺胺甲口恶唑的耐药率在过去的20年间不断上升,2003年比利时为8.5%,肯尼亚为55.2%[3],2010年我国为64.5%[1],我院为57.1%。
同样,流感嗜血杆菌对四环素的耐药率较高,四环素类抗菌作用是药物结合到细菌核糖体的30S并阻止tRNA接合到A和P位点上[10]。对四环素的耐药性是由于流感嗜血杆菌的细胞膜上的tet(B)基因产生的外排机制,这种基因位于接合质粒上[11-12]。tet(B)基因编码的外排蛋白可以对四环素和米诺环素耐药。由氨苄西林、卡那霉素的耐药基因介导的质粒经常转移到四环素上,导致其耐药,这一现象已在比利时、西班牙和古巴分离的B型流感嗜血杆菌中报道[13-14]。流感嗜血杆菌对四环素和氯霉素的耐药性增高,与质粒介导的氨苄西林、氯霉素、四环素、卡那霉素耐药基因有关。根据2012 CLSI标准,如果细菌对四环素耐药,同时也对强力霉素和米诺环素耐药。
从我院分离的流感嗜血杆菌4年来对头孢噻肟、氯霉素和四环素的耐药率有所降低,说明医师应用抗菌药物合理。但由于我国流感嗜血杆菌分离率较低,尚未对该菌造成的感染及其耐药性引起重视,如果不合理使用抗菌药物的现象持续下去,将会有高耐药株出现[15]。
[1] 朱德妹,汪复,胡付品,等.2010年中国CHINET细菌耐药性监测[J].中国感染与化疗杂志,2011,9(5):321-329.
[2] Farrell DJ,Morrissey I,Bakker S,et al.Global distribution of TEM-1 and ROB-1 beta-lactamases in Haemophilus influenzae [J].J Antimicrob Chemother,2005,56(4):773.
[3] Jacobs MR,Felmingham D,Appelbaum PC,et al.The Alexander Project 1998-2000:susceptibility of pathogens isolated from community-acquired respiratory tract infection to commonly used antimicrobial agents[J].J Antimicrob Chemother,2003,52(1):229-246.
[4] Tristram S,Jacobs MR,Appelbaum PC.Antimicrobial resistance Haemophilus influenzae [J].Clin Microbiol Rev,2007,20(2):368-389.
[5] Bozdogan B,Tristram S,Appelbaum PC.Combination of altered PBPs and expression of cloned extended-spectrum betalactamases confers cefotaxime resistance in Haemophilus influenzae[J].J Antimicrob Chemother,2006,57(4):747-749.
[6] Pitout M,MacDonald K,Musgrave H,et al.Characterization of extended spectrum beta-lactamase(ESBL)activity in Haemophilus influenzae[M]//Program and Abstracts of the 42nd Interscience Conference on Antimicrobials and Chemotherapy,San Diego,CA.American Society for Microbiology,Washington,DC,USA.2002:96.
[7] Kim IS,Ki CS,Kim S,et al.Diversity of ampicillin resistance genes and antimicrobial susceptibility patterns in Haemophilus influenzae strains isolated in Korea[J].Antimicrob Agents Chemother,2007,51(2):453-460.
[8] Garcia-Cobos S,Campos J,Lazaro E,et al.Ampicillin-resistant non-beta-lactamase-producing Haemophilus influenzae in Spain:recent emergence of clonal isolates with increased resistance to cefotaxime and cefixime[J].Antimicrob Agents Chemother,2007,51(7):2564-2573.
[9] Nazir J,Urban C,Mariano N,et al.Quinolone-resistant Haemophilus influenzae in a long-term care facility:clinical and molecular epidemiology[J].Clin Infect Dis,2004,38(11):1564-1569.
[10] Chopra I,Hawkey PM,Hinton M.Tetracyclines,molecular and clinical aspects[J].J Antimicrob Chemother,1992,29(3):245-277.
[11] Marshall B,Roberts M,Smith A,et al.Homogeneity of transferable tetracycline-resistance determinants in Haemophilus species[J].J Infect Dis,1984,149(6):1028-1029.
[12] Chopra I,Roberts M.Tetracycline antibiotics:mode of action,applications,molecular biology,and epidemiology of bacterial resistance[J].Microbiol Mol Biol Rev,2001,65(2):232-260.
[13] Campos J,Chanyangam M,deGroot R,et al.Genetic relatedness of antibiotic resistance determinants in multiply resistant Hemophilus influenzae[J].J Infect Dis,1989,160(5):810-817.
[14] Levy J,Verhaegen G,De Mol P,et al.Molecular characterization of resistance plasmids in epidemiologi-cally unrelated strains of multiresistant Haemophilus influenzae [J].J Infect Dis,1993,168(1):177-187.
[15] 楚文瑛.临床微生物实验室需重视嗜血杆菌的分离培养[J].国际检验医学杂志,2011,32(18):2160-2162.
