藏东南地区种子植物的区系组成和物种多样性
2013-07-29游金娥冯建孟
游金娥,冯建孟
大理学院古城校区农学与生物科学学院, 云南 大理 671003
藏东南地区位于西藏东南雅鲁藏布江下游。由于喜马拉雅山脉、念青唐古拉山脉和横断山脉等巨大山体横亘其中,故地形复杂,海拔高差巨大(总体海拔高差超过7000 m)[1]。如此复杂的地形和巨大的海拔高差可能暗示着该地区拥有多样的植被类型和丰富的区系成分类型。因此,藏东南地区可能是进行生物多样性和植物区系研究的理想区域。2002年,韩景军等发现该地区的林芝云杉Picea likiangensis var. linzhiensis幼林拥有较高的物种多样性[2]。2003年,朱万泽、范建荣[3]发现该地区珍稀濒危植物区系地理成分十分复杂,包含了吴征镒属分布区类型系统中的所有分布区类型。最近,李晖等[4]探讨了藏东南森林植被界线问题。但是综观过去的研究,笔者发现,截至目前,有关于该地区植物区系的总体构成以及植物多样性的专题性研究还少见报道。目前人们还不清楚该地区的植物区系组成、重要科、属构成及其与周边地区的区系联系。因此,本研究的目的在于利用大尺度的植物分布数据,探讨藏东南地区种子植物的区系组成和物种多样性,为更好地理解藏东南地区植物的起源、进化历史和生物多样性的形成机制提供理论基础。
1 研究区域概况
关于“藏东南地区”这一地理位置名词, 目前还没有统一的定义。广义上的“藏东南地区”不仅包括西藏自治区东南部,还包括云南的西北部和四川的西南部。而狭义的“藏东南地区”则特指林芝地区。本研究采用狭义的“藏东南地区”,即林芝地区(含林芝、工布江达、米林、墨脱、波密、察隅、朗县等7县,图1)。林芝地区地处西藏自治区的东南部,雅鲁藏布江中下游,地理位置92°11′—98°40′E,27°48′—30°53′N,东西长646.7 km,南北宽353.2 km。该地区东邻昌都地区和云南滇西北地区,北连那曲地区,西部和西南部与拉萨市、山南地区接壤,南部与印度、缅甸两国交界[5]。研究区域国土面积为114585.29 km2,占西藏地区总面积的10%左右[5]。总人口约为16万人,年降水量650 mm左右,年平均温度8.7 ℃,平均日照时数为2022 h,无霜期180 d。区域平均海拔3 100 m,地势南低北高,海拔高低悬殊,海拔最高的是7756 m的南迦巴瓦峰,而海拔最低的下察隅仅为500 m,海拔高差达到了7256 m[5]。与此相一致,区域内立体气候明显,从低海拔到高海拔,依次受到热带、亚热带、温带和寒带气候的控制[5]。受多样的立体气候影响,区域内植被垂直带完整。从低海拔到高海拔,依次可见湿性季雨林、常绿阔叶林、常绿阔叶和落叶阔叶混交林、落叶阔叶林、针阔叶混交林、针叶林、灌丛、草甸、草原等[6]。从地质历史上看,该区的南迦巴马峰处于印度板块与欧亚板块碰撞的缝合线的东端,碰撞之后的新构造运动,使其上升幅度增大,由此对植被垂直带的形成和植物区系分化产生了深刻的影响,因此,该区域在植物区系地理研究方面具有十分重要的研究价值[7]。
图1 研究区域示意图 Fig.1 Location of the study area
2 数据来源
本研究所用的植物物种数据来自于吴征镒所编写的《西藏植物志》(1~5卷)[8-12],该植物志是目前国内有关西藏地区植物最为广泛接受的科学著作。本研究根据该植物志所记载的所有种子植物物种(含种及种下单位)的分类地位和分布信息,进行了归类、整理和分析。以此为基础,我们建立了藏东南地区种子植物物种数据库。此外,本研究所采用的林芝地区人口、国土面积和海拔信息均来自西藏自治区地图册[13]及相应文献[5]。用于区系联系对比分析的西双版纳地区植物物种数据、独龙江地区植物物种数据以及南滚河保护区植物物种数据分别来自《云南植物志》(1~16卷)[14-29]、《独龙江地区植物》[30]和《中国南滚河国家级自然保护区》[31]。
3 研究方法
通过对数据库的分析和统计,我们获得了藏东南地区种子植物的物种数及其科、属的组成,并按照所含物种的多少,获得比较重要的(前10位)的科、属,据此了解研究区域内植物物种的主要组成。同时,还我们进行了区系分化强度的分析,即种数与属数的比值和属数与科数的比值。此外,为分别获得科、属层次上的区系组成,我们根据吴征镒关于世界种子植物科的分布类型[32-33]以及吴征镒关于中国种子属的分布区类型[33],进行区系组成和联系的分析。15种分布区类型分别为广布(T1)、泛热带(T2)、东亚及热带南美间断(T3)、旧世界热带(T4)、热带亚洲至热带大洋洲(T5)、热带亚洲及热带非洲(T6)、热带亚洲(T7)、北温带(T8)、东北及北美间断(T9)、旧世界温带(T10)、温带亚洲(T11)、地中海区、西亚至中亚(T12)、中亚(T13)、东亚(T14)和中国特有(T15)。此外,T2-T15通称为非世界分布类型,T2-T7为热带区系成分,T8-T14为温带区系成分。通过对于各区系的比重分析,进行了科水平上和属水平上的区系分析。上述分析和制图主要通过MS Excel和ArcGIS Desktop 8.2TM实现。
4 研究结果
4.1 藏东南地区植物物种组成

表1 藏东南地区重要科的组成 Table 1 Composition of important families in Southeast Tibet
研究结果表明,藏东南地区拥有种子植物3 114种,分属于876属,153科。从科的分化强度来看,区系分化强度为5.73;从属的分化强度来看,区系分化强度为3.55。从表1可以看出,研究区域内较为重要的科)前10名)依次为菊科Rompositae 242种、杜鹃花科Ericaceae 226种、蔷薇科Rosaceae 188种、禾本科Poaceae 144种、毛茛科Ranunculaceae 107种、兰科Orchidaceae 97种、蝶形花科Papilionaceae 96种、虎耳草科Saxifragaceae 91种、玄参科Scrophulariaceae 83种、唇形科Labiatae 81种、龙胆科Gentianaceae 81种。另外,我们也发现,除蝶形花科所占比重为37%以外,其他各科所含物种数均占西藏地区各科总物种数的40%以上。尤其是杜鹃花科,研究区域所含总物种数占西藏地区的82%。进一步分析,我们发现这11个较大的科所含种子植物物种之和为1436种,占了藏东南地区总物种的46%左右。从表2可以看出,藏东南地区较大的属前10名依次为杜鹃属Rhododendron 146种、报春花属Primula 57种、龙胆属Gentiana 55种、虎耳草属Saxifraga 52种、马先蒿属Pedicularis 49种、柳属Salix 38种、小蘖属Berberis 37种、紫堇属Corydalis 36种、蓼属Polygonum 33种、委陵菜属Potentilla 31种、悬钩子属Rubus 31种。除紫堇属外,其他各属所含物种数均占西藏各属所含物种总数的40%以上。其中,杜鹃属与悬钩子属所占比重均较高,分别占77%与78%。
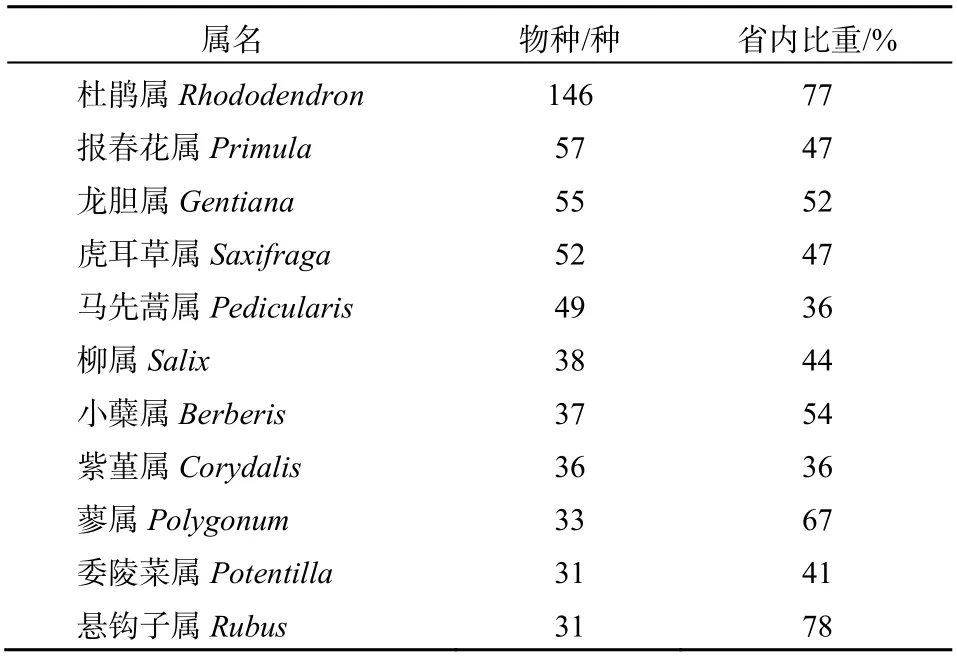
表2 藏东南地区重要属的组成 Table 2 Composition of important genera in Southeast Tibet
4.2 科水平上的区系组成

表3 藏东南地区种子植物科的区系组成 Table 3 Flora composition at family level in Southeast Tibet
从表3可以看出,比重最大的分布区类型是泛热带分布成分(T2),所占比重高达51.9%。这可能意味着,藏东南地区种子植物区系在起源上与全球大部分热带地区有着十分密切的联系。另外, 北温带(T8)分布成分所占比重位居第二,为22.6%,拥有24科。其次,研究区域内东亚及热带南美间断(T3)分布成分拥有11科,所占比重为10.4%。除此之外,其他区系成分所占比重均比较小。从区系总体构成来看,热带区系成分所占比重为70%左右,而温带区系成分的比重则仅为30%左右。
4.3 属水平上的区系组成
从表4可以看出,所占比重最大的区系分布类型是北温带分布(T8),所占比重为21%,拥有171属,这可能意味着藏东南地区的植物区系与北温带地区之间存在着密切联系。其次,所占比重较大的分布区类型为泛热带分布成分(T2),所占比重为15.5%,共拥有126属。其他所占比重较大的区系分布类型为热带亚洲(T7)和东亚分布成分(T14),分别为12.4%和14.4%。所占比重较小的区系分布类型为温带亚洲分布(T11)和地中海、西亚至中亚分布(T12)以及中亚分布(T13),分别为1.8%、1.8%和1.2%。
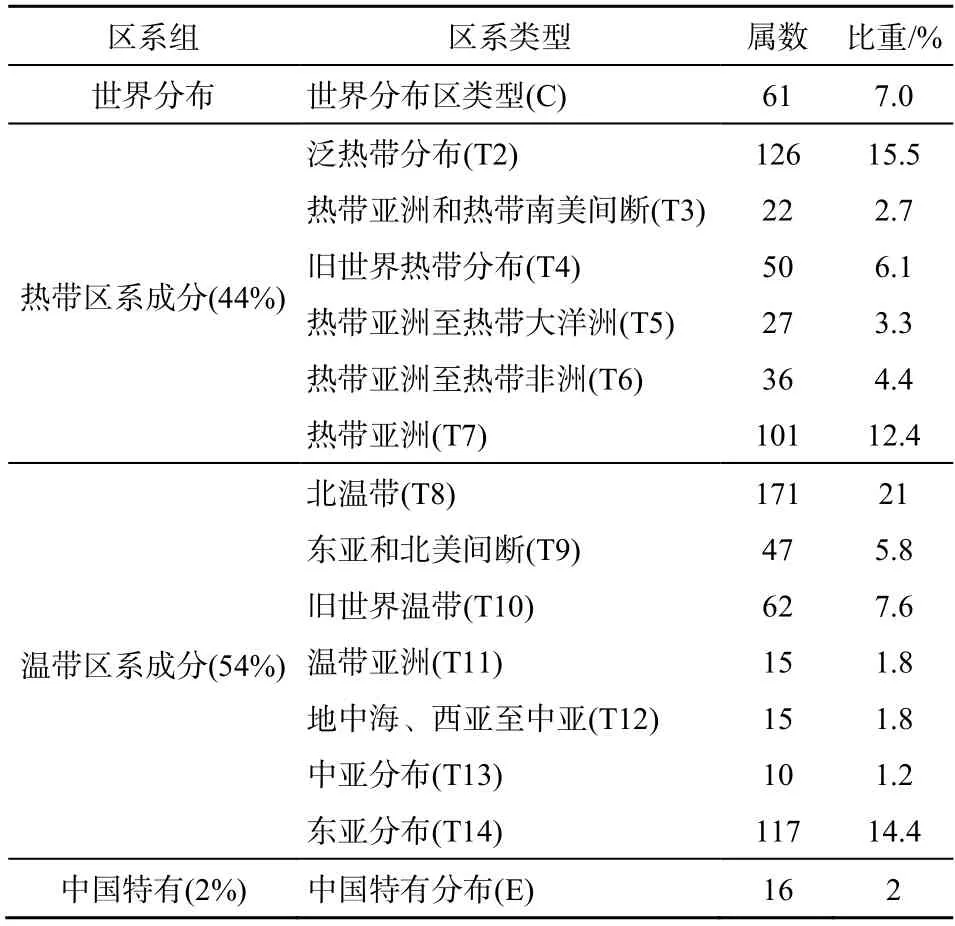
表4 藏东南地区种子植物属的区系组成 Table 4 Flora composition at genus level in Southeast Tibet
4.4 与邻近地区的比较
由表5可以看出,在重要科的前10名中,独龙江与藏东南地区拥有7个相同的科,即菊科Compositae、兰科Orchidaceae、杜鹃花科Ericaceae、禾本科Gramineae、蔷薇科Rosaceae、龙胆科Gentianaceae和玄参科Scrophulariaceae。这说明了独龙江与藏东南地区有着非常相似的一级科组成。西双版纳地区与藏东南地区拥有5个相同的科,即兰科Orchidaceae、蝶形花科Papilionaceae、菊科Compositae、禾本科Gramineae和唇形科Labiatae;而南滚河自然保护区与藏东南地区仅仅有3个相同的科。由表6可以看出,在重要属的前10名中,独龙江与藏东南地区拥有7个相同的属,即杜鹃属Rhododendron、报春花属Primula、悬钩子属Rubus、蓼属Polygonum、柳属Salix、龙胆属Gentiana和马先蒿属Pedicularis。而在西双版纳地区与南滚河自然保护区,与藏东南地区相同的属仅分别为1个和2个。这可能意味着相比独龙江地区,西双版纳地区与南滚河自然保护区与藏东南地区之间的区系联系并不密切。
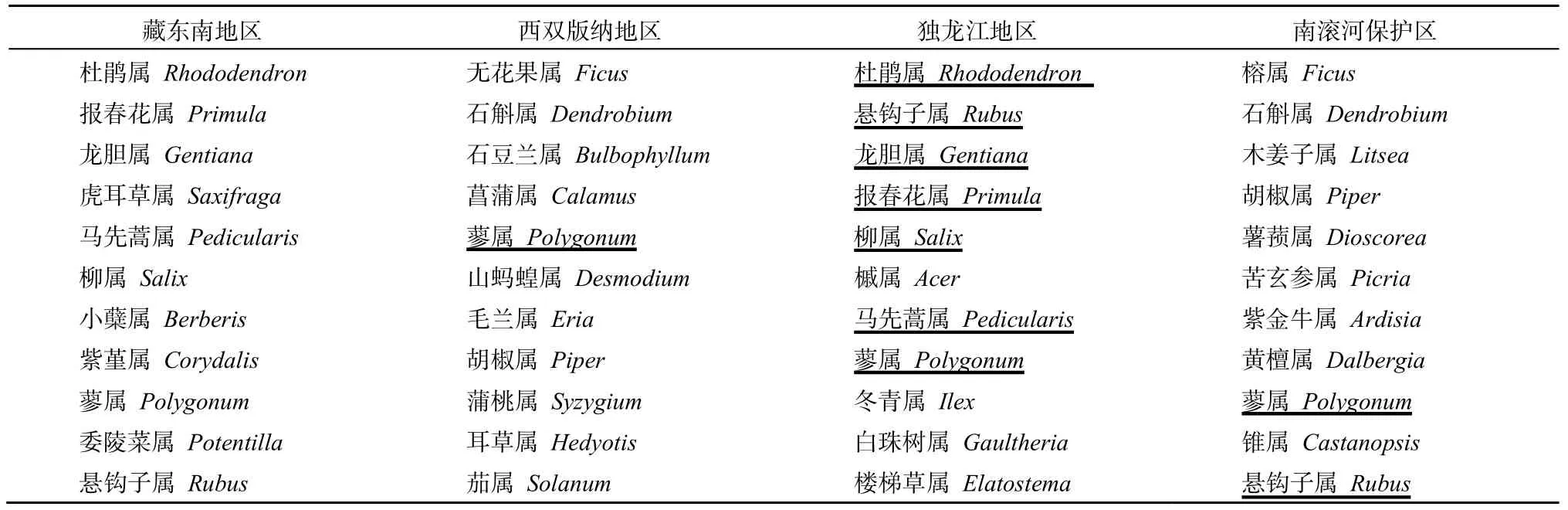
表6 藏东南地区与西双版纳、独龙江地区、南滚河自然保护区优势属的比较 Table 6 Comparison of dominant genera between Southeast Tibet and Xishuangbanna regions, Drung River regions and Nangun River Nature Reserve
5 讨论
藏东南地区由于其巨大的海拔高差、特殊的地质历史和复杂的植物区系组成,相关研究具有重要意义。本研究根据《西藏植物志》(1~5卷)[8-12],分别在科、属水平上探讨了该地区植物区系的组成、起源和植物多样性,并在一定程度上证实了青藏高原隆起对植物区系分化的影响,这对于我们进一步理解藏东南地区植物区系的起源、分布和迁移以及当地植物多样性的保护均具有重要意义。
研究结果表明,藏东南地区以仅占西藏地区10%左右的国土面积,拥有远远超过其国土面积所占比例的植物多样性。首先,在科的水平上,藏东南地区拥有153个科,占到了西藏地区种子植物总科数的88%。其次,在属的水平上,藏东南拥有876个属,占到了西藏地区种子植物总属数的78%。最后,从种的水平上,藏东南地区拥有种子植物3114种,占到了西藏地区种子植物总物种数的54%。由此可见,藏东南地区可能是西藏地区植物多样性最为丰富的地区之一。除此之外,我们也发现排名前 10的重要科中(表1),除蝶形花科以外,其他重要科所含物种数均占西藏地区的相应科的40%以上,尤其是杜鹃花科,其所含总物种数占西藏地区的82%。在属的水平上,排名前10的重要属中(表2),除紫堇属以外,其他各属所拥有的物种数均占西藏地区的40%以上,尤其是杜鹃属与悬钩子属所占比重均较高,分别占77%与78%。这可能意味着藏东南地区也可能是上述重要科、属在西藏地区的主要分布中心之一。
藏东南地区种子植物在科的水平,热带区系成分所占比重为70%,表现出明显的热带区系性质;但在属的水平,温带区系成分所占比重为54%,表现出明显的温带区系性质。如此区系性质的巨大反差,可能与科、属所代表的不同地质历史和青藏高原过去的地质历史有关。科区系一般被认为代表着相对久远的地质历史,而属区系则被认为指示相对较近的地质历史[34-35]。在隆起之前,青藏高原被认为是热带低地[36-37]。与此相一致,本研究结果表明,在科的水平上,植物区系以热带区系为主,表现出明显的热带区系性质,这可能正反映了这一久远的地质历史阶段。自始新世开始,由于喜马拉雅山及青藏高原的强烈隆升,气候系统发生明显的转变,相应的植物区系亦发生强烈的分化。青藏高原的隆起导致热带低地气候被相对温凉的高原气候所代替,植物为适应这一巨变,发生了区系成分的强烈分化[38]。因此,从科的水平到属的水平,热带区系比重的降低和温带区系比重的增加,正反映了青藏高原隆起的地质历史过程,并在一定程度上也证实了这一地质历史过程。
在属的水平上,热带区系成分和温带区系成分所占比重分别为44%与54%,比较接近,这可能意味着,在属的水平上,研究区域内的植物区系表现出一定的过渡性。这与藏东南地区的巨大海拔高差是一致的。藏东南地区海拔高差高达7000 m以上,最低海拔为500 m,而最高海拔却达到了7756 m。这就意味着在巨大的海拔梯度上,低海拔地区可能分布着具有明显热带区系性质的热带或亚热带植被,而在高海拔地区,又可能分布着具有明显寒温性质的高寒植被。两者的交汇导致了研究区域表现出一定的区系过渡性。从地质历史上来看,研究区域的区系过渡性也可能与青藏高原的隆升以及随之受影响的气候系统有关。有学者认为,在青藏高原隆升过程中区系分化存在着一定的区内差异性,如藏东南等地保留了部分热带、亚热带成分[38-39]。上述热带、亚热带区系成分与隆起过程中的区系分化和隆起过程中保留下来的温带区系成分相互交汇亦可能导致了一定的区系过渡性[40-41]。
研究结果表明,独龙江地区无论在科的水平还是在属的水平,均与藏东南地区拥有高度相似的重要科、属组成,这可能意味着独龙江地区的植物区系与藏东南地区之间存在着密切的联系,或者说两者拥有相似的区系地理背景。独龙江地区位于滇西北三江弧形构造带上,并由此向青藏高原过渡,而且与藏东南地区的察隅县相邻[30,42]。而且,在地质历史上,独龙江地区与藏东南地区同为冈瓦纳古陆的一部分,这意味着两者可能有着共同的区系起源。自新生代以来,独龙江地区与藏东南地区均深受印度板块和欧亚板块碰撞这一重大地质历史事件的影响,拥有相似的地质历史过程。因此,两者的植物区系具有相似的起源,经历着相似的区系分化过程[30]。因此,两者之间的区系联系十分密切。
6 结论
本文利用大尺度的物种分布数据,分析了藏东南地区的植物区系组成和物种多样性。主要结论如下。
1)藏东南地区可能是西藏地区植物多样性的热点地区之一。
2)在科水平上,热带区系成分比重占到了70%左右;但在属的水平上,热带区系比重仅占44%左右。
3)科、属水平上不同的区系组成,尤其是温、热带区系成分比重的巨大差异, 可能与科和属所代表的不同地质历史以及青藏高原的隆升有关。
4)相比西双版纳地区和南滚河自然保护区,独龙江地区的植物区系与藏东南之间存在着更为紧密的区系联系。
[1] 卢杰,兰小中, 罗建, 等. 林芝地区珍惜濒危藏药植物资源调查与评价[J]. 资源科学, 2011, 33(12): 2362-2369.
[2] 韩景军,肖文发,郭泉水, 等. 西藏林芝县林芝云杉幼林更新与物种多样性指数研究[J]. 林业科学, 2002, 38(5): 166-168.
[3] 朱万泽, 范建荣. 西藏珍稀濒危植物区系特征及其保护[J]. 山地学报, 2003, 21(增刊): 31-39.
[4] 李晖, 扎西群培. 藏东南森林植被西界的刍议[J]. 西藏科技, 2011(2): 70-71.
[5] 杨文凤, 张格杰, 张永青, 等. 藏东南生态旅游资源及其开发利用[J]. 林业调查规划, 2006, (6): 120-124
[6] 何关福. 漫谈西藏植物资源[J]. 植物学杂志, 1975 (3): 21-23.
[7] 彭补拙. 南迦巴马峰地区垂直自然带的初步研究[J]. 山地研究, 1984, 2(3): 182-189.
[8] 中国科学院青藏高原综合科学考察队. 西藏植物志 (卷1)[M]. 北京: 科学出版社, 1983: 1-791.
[9] 中国科学院青藏高原综合科学考察队. 西藏植物志 (卷2) [M]. 北京: 科学出版社, 1985: 1-956.
[10] 中国科学院青藏高原综合科学考察队.西藏植物志 (卷3) [M]. 北京: 科学出版社, 1986: 1-1046.
[11] 中国科学院青藏高原综合科学考察队. 西藏植物志 (卷4) [M]. 北京: 科学出版社, 1985: 1-1021.
[12] 中国科学院青藏高原综合科学考察队. 西藏植物志 (卷5) [M]. 北京: 科学出版社, 1987: 1-955.
[13] 尹嘉珉. 西藏自治区地图册[M]. 北京: 中国地图出版社, 2005: 1-151.
[14] 中国科学院昆明植物所. 云南植物志 (卷1)[M]. 北京: 科学出版社, 1977: 1-870.
[15] 中国科学院昆明植物所. 云南植物志 (卷2) [M]. 北京: 科学出版社, 1979: 1-889.
[16] 中国科学院昆明植物所. 云南植物志 (卷3) [M]. 北京: 科学出版社, 1983: 1-795.
[17] 中国科学院昆明植物所. 云南植物志 (卷4) [M]. 北京: 科学出版社, 1986: 1-823.
[18] 中国科学院昆明植物所. 云南植物志 (卷5) [M]. 北京: 科学出版社, 1991: 1-809.
[19] 中国科学院昆明植物所. 云南植物志 (卷6) [M]. 北京: 科学出版社, 1995: 1-910.
[20] 中国科学院昆明植物所. 云南植物志 (卷7) [M]. 北京: 科学出版社, 1997: 1-888.
[21] 中国科学院昆明植物所. 云南植物志 (卷8) [M]. 北京: 科学出版社, 1997: 1-778.
[22] 中国科学院昆明植物所. 云南植物志 (卷11) [M]. 北京: 科学出版社, 2000: 1-754..
[23] 中国科学院昆明植物所. 云南植物志 (卷9) [M]. 北京: 科学出版社, 2003: 1-907.
[24] 中国科学院昆明植物所. 云南植物志 (卷14) [M]. 北京: 科学出版社, 2003: 1-885.
[25] 中国科学院昆明植物所. 云南植物志 (卷15) [M]. 北京: 科学出版社, 2003: 1-847.
[26] 中国科学院昆明植物所. 云南植物志 (卷13) [M]. 北京: 科学出版社, 2004: 1-918.
[27] 中国科学院昆明植物所. 云南植物志 (卷16) [M]. 北京: 科学出版社, 2005: 1-876.
[28] 中国科学院昆明植物所. 云南植物志 (卷10) [M]. 北京: 科学出版社, 2006: 1-929.
[29] 中国科学院昆明植物所. 云南植物志 (卷12) [M]. 北京: 科学出版社, 2006: 1-884.
[30] 李恒. 中国科学院昆明植物研究所. 独龙江地区植物[M]. 昆明:云南科技出版社, 1993: 1-381.
[31] 杨宇明, 杜凡. 中国南滚河国家级自然保护区[M]. 昆明:云南科技出版社, 2004: 1-385.
[32] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245-257.
[33] 吴征镒. 中国种子植物区系统计分析[J]. 云南植物研究, 1991, 4(增刊): 1-139.
[34] 王荷生. 植物区系地理[M]. 北京: 科学出版社, 1992: 1-180.
[35] 吴征镒, 朱彦丞, 姜汉侨. 云南植被[M]. 北京: 科学出版社, 1987: 1-200.
[36] 唐领余, 沈才明. 青藏高原上新世以来植被与气候研究进展[J]. 地球科学进展, 1996, 11(2): 198-203.
[37] 耿国仓. 从植物化石看西藏地质的今昔[J]. 植物杂志, 1978, (2): 41-42.
[38] 唐领余, 沈才明. 青藏高原晚新生代植被史及其气候特征[J]. 微体古生物学报, 1996, 13(4): 321-337.
[39] 魏明建, 王成善, 万晓樵, 等. 第三纪青藏高原面高程与古植被变迁[J]. 现代地质, 1998, 12(3): 318-326.
[40] 郭双兴. 西藏新生代植物区系和植被演替[C]//中国青藏高原研究会. 中国青藏高原研究会2006学术年会论文摘要汇编. 北京: 中国青藏高原研究会, 2006: 45-47.
[41] 冯建孟, 徐成东, 查凤书, 等. 长江上游滇西北地区植物区系组成及物种多样性[J]. 长江流域资源与环境, 2010, 19(1): 65-72.
[42] 李先之. 独龙江流域地貌[J]. 云南地理环境研究, 1996, 8(1): 59-72.
