不同种群高山松1年生播种苗木生长节律及其变异1)
2013-03-05蔡年辉许玉兰白青松张瑞丽段安安
蔡年辉 许玉兰 白青松 张瑞丽 魏 巍 段安安 李 悦
(西南山地森林保育与利用省部共建教育部重点实验室(西南林业大学),昆明,650224) (北京林业大学)
高山松(Pinus densata)是我国西部高山地区的特有树种,地理分布北接油松(P. tabulaeformis)分布区的南缘(四川西北部、青海南部),南依云南松(P. yunnanensis)的北界(云南西北部、四川西南部、西藏东部),其形态和解剖性状介于云南松和油松之间[1]。因此,曾有人推测它可能是云南松和油松的天然杂种[2]。这一推测于1990年首次获得遗传学证据,Wang 等[3]用所获得遗传标记证实高山松种群包含了云南松和油松的叶绿体基因组,有力地支持了高山松杂交起源的假设,并探讨其生长于两个亲本种都不能正常生长的高海拔地带,后来陆续得到多方面的证实[4-10],并从等位酶、cp DNA 和mt DNA 等方面揭示种群间复杂多样的杂交起源和遗传组成,不同种群亲本种贡献差异较大[4-7],这些复杂的成种背景和生境导致了很强的遗传分化[5,10],其种实性状也表现明显的变异[11]。笔者在前期对不同种群种实性状和种子萌发分析的基础上,进一步研究高山松不同种群在理论亲本种云南松生境下苗期生长表现,揭示不同杂交起源和遗传背景的高山松种群1年生苗木生长节律及其变异,为深入认识高山松同倍体杂交成种特点和种群进化机制提供基础。
1 试验地概况
试验地设在西南林业大学苗圃,海拔1 945 m,属北亚热带半湿润高原季风气候,年平均气温14.7℃、绝对最低温-9 ℃、绝对最高温32.5 ℃。年降水量700 ~1 100 mm,年平均相对湿度68.2%。
2 材料与方法
在高山松主要分布区内选择8 个代表性种群(表1)。每个种群均为成熟林分,在各种群内随机选取生长正常的成年树木20 株,各样株间距不小于100 m,每株树取树冠中部发育正常的成熟球果,所取球果均按种群混合,获得混合种子,用于播种和苗期观测研究。

表1 采样种群的地理位置
按照常规整地播种,完全随机区组排列,每个种群为1 个试验小区,每个试验小区60 粒,5 次重复,共40 个试验小区2 400 粒,种子播种株行距为5 cm×15 cm。种子萌发结束后(播种后第43 天)开始测定,按照系统抽样标记待测植株,每小区抽取5株,每个种群25 株,每月定期测量记录每样株的苗高与地径(0. 1 mm),直至苗木生长停止(播种后268 d)为止。
参照文献[12]、[13],以生长时间(t)为自变量,累积生长量(y)为因变量,利用各种群苗高、地径累积生长量的平均值与生长时间的关系对生长进程用“S”型生长曲线拟合,分析高山松1年生苗木的生长节律及其变异特征,所用的Logistic 数字表达式为:y=k/(1 +ea-bt),式中a、b、k 为待测参数,其中k 为生长量的理论极限值上限值或生长潜力,t 为生长时间(以播种日为0 算起),y 为生长量[14-16],参数a 与生长初始值及生长特征点出现的时间有关,b 为生长速率参数[17]。对该表达式进一步求导,可得该曲线瞬时最大斜率对应的时间点t0(速生点)以及曲线上瞬时斜率连续变化最快的两个时间点即生长拐点t1(增速点,即速生期起始时间)和t2(减速点,即速生期结束时间),其中t0=a/b、t1=(a -1.317)/b、t2=(a +1.317)/b[15-16,18],t1-t2为速生期持续时间[19],苗高和地径生长量的拟合模型经显著性检验以确定其相关系数。观测数据采用DPS7.05 进行方程拟合和参数估算,Excel 进行分析与绘图。
3 结果与分析
3.1 高山松苗木生长节律模型拟合
分别对高山松不同种群的苗高、地径生长动态进行拟合,其结果见表2和图1。
从表2可知,高山松苗高各种群拟合方程的相关系数为0.988 8 ~0.998 2,平均为0.995 7,地径各种群拟合方程的相关系数为0.967 3 ~0.997 6,平均为0.989 3,方程回归性检验均达极显著水平(p <0.01),说明利用Logistic 拟合曲线与实测值间的符合程度较高,用拟合方程的理论值来估测实际值具有较高的准确性。因此,应用Logistic 曲线拟合高山松1年生苗高和地径的生长节律效果较好。

表2 高山松不同种群苗木生长动态Logistics 模型拟合结果
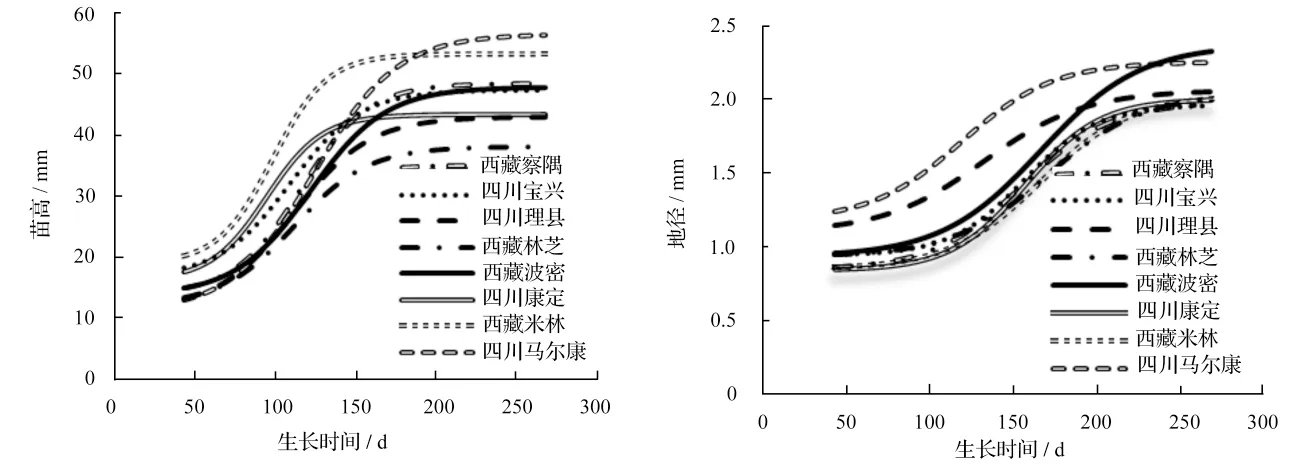
图1 高山松不同种群1年生苗苗高、地径生长Logistics 拟合曲线
由图1可知,不同种群1年生苗苗高与地径的生长均呈现“慢-快-慢”的生长节律,但不同种群的速生期起始时间、结束时间和持续时间差异较大,相应的生长量差异也比较明显。西藏米林种群在各阶段苗高生长表现均较好;四川康定种群在前期生长较快,到中后期逐渐减慢,前期竞争力较强;四川马尔康种群前期生长较慢,但中后期加快,后期竞争力较强;西藏林芝种群在各阶段生长表现均较慢。四川马尔康和四川理县种群在各阶段地径生长均较好;西藏波密种群前期生长缓慢,而中后期生长逐渐加快;其它几个种群各阶段生长变化不大,表现为中、下水平。这种差异对不同种群在特定生境中苗木的建成具有不同的意义,生长比较快的,竞争力较强[20],能在特定生境中很快占有空间,从而有利于树木的生长。
3.2 高山松苗木生长阶段划分及其特点
根据不同种群的苗木生长Logistic 模型及其特点,按生长曲线上的拐点,划分速生期的起始时间和结束时间[15,18],从而划分3 个阶段:生长前期、速生期、生长后期,依此获得生长期速度最快的生长点t0(速生点)、速生期起始时间t1(增速点)和速生期结束时间t2(减速点)3 个主要的物候点,并求算出各个阶段的生长参数,结果见表3。从表3可知,不同种群的苗高和地径生长进程有所不同。苗高生长前期、速生期和生长后期3 个阶段的持续时间分别为28 ~48、45 ~68、110 ~147 d,地径分别40 ~89、58 ~79、61 ~113 d,前2 个阶段苗高生长比地径生长快,持续时间短。苗高生长的速生点为51 ~81 d,地径生长为76 ~127 d,这些结果说明不同种群苗木生长物候存在差异,且每个种群均表现为苗高较地径提早进入速生期,持续时间短,即苗高与地径生长间存在异速生长的现象。
除物候方面的差异外,不同种群各阶段苗高、地径生长量存在很大差异。从各阶段的生长速度来看,苗高日均生长量在3 个阶段分别为0.11 ~0.18、0.25 ~0.44、0.04 ~0.09 mm,以速生期的日均生长量最高,为另2 个生长阶段的3 ~6 倍;地径日均生长量在3 个阶段分别为0.003 ~0.004、0.007 ~0.011、0.002 ~0.004 mm,速生期的日均生长量为另2 个生长阶段的3 ~4 倍。较早进入速生期的是纬度较高的四川马尔康种群和四川理县种群,苗高和地径生长表现迟缓的是经度偏小的西藏林芝种群,这可能与不同种群的起源有关,四川马尔康和四川理县区域已证实是高山松的古老起源中心[21],起源中心复杂的遗传多样性可能导致较强的适应性,在新的环境条件下也能快速生长。
虽然各阶段的物候和生长存在差异,但不同种群苗木的苗高和地径生长均表现为速生期的生长量比例大。生长前期经历的时间较长,累积生长量不到总生长量的20%;速生期所经历的时间占整个生长期的1/4 左右,但生长量却占总生长量57.00%以上,苗高生长各种群所占比例为57.34% ~57.93%,地径生长各种群所占比例为57.29% ~58.47%,即在1/4 的生长期内完成了全年一半以上的生长量。因此,在符合“S”型生长曲线的林木个体年生长过程中,“速生期”是生长过程的关键阶段[22],速生期时间较短,但生长量较大[23]。不同种群苗高与地径的速生期起始时间、结束时间以及持续时间相差较大。除四川康定种群外,其它种群均表现为苗高生长速生期持续时间比地径生长的短。
3.3 高山松苗木生长拟合模型参数与生长量间的关系
为进一步讨论拟合的结果,对8 个高山松种群1年生苗木理论生长极值与实测值进行比较,结果表明:高山松1年生苗木苗高、地径理论生长极值k与实测值呈极显著的正相关关系,相关系数分别为0.995 9 和0.931 7。因此,可以用k 值来代替苗期的年生长量实测值,也可以用k 值作为苗木生长分析评价指标,这在其它的研究报道中也有类似的结果[22,24]。

表3 高山松1年生苗苗高、地径生长情况
高山松1年生苗木苗高、地径速生期时间短,但生长量所占比例高,8 个种群1年生苗高和地径速生期内生长量与理论生长极值k 间呈极显著的正相关关系,相关系数分别为0.999 8 和0.998 8,进一步说明,苗木生长速生期的生长量对整个苗木生长量有决定性作用,即速生期内生长量大,则整个生长期生长量大。
速生期进入的早晚及结束时间直接决定速生期持续时间的长短,一般要选择速生始点较早、速生终点较晚、速生持续期较长的有利于苗期年生长量的增加[24],从而对生长竞争有一定的影响。对8 个种群速生期的起始时间和结束时间进行拟合,相关系数为0.859 2 和0.819 1,存在明显的正相关关系,即开始时间较早的,同时也较早结束速生生长,说明高山松苗木的速生期起始时间相对比较稳定,速生期的生长量相对比例也相差不大,这从前面的分析中也可看出,尽管各种群的生长物候差异较大,但各种群苗高、地径生长在速生期的比例比较接近,为57% ~58%。
4 结论与讨论
高山松播种后,7 d 开始萌发,持续2 周后苗木基本出整齐,待生长基本稳定后开始测定,首次测定时间是播种后43 d,此时形成一定的根系和叶面积,具备吸收土壤养分和制造营养物质的功能,幼苗开始逐渐生长,但由于前期叶面积和根系都较小,真叶没完全发育,来源于子叶和幼嫩真叶的光合同化物远远不够维持较快的生长速度[25],生长较为缓慢;经过生长前期后,幼苗叶面积逐渐增大,根系逐渐发达,生物量不断积累,再加上苗木对外界条件的适应性增强,幼苗进入速生期;后期苗高、地径生长速度逐渐下降,直至生长停止。在测定的225 d 内,苗木生长呈现“慢—快—慢”的生长节律,表现为“S”型生长曲线,经拟合生长进程符合Logistic 增长,拟合效果较好。
根据高山松不同种群的苗木生长Logistic 模型及其特点,将1年生苗木生长节律划分为3 个阶段:生长前期、速生期和生长后期。不同种群各阶段的持续时间、生长速率及生长量均存在差异,总体表现为速生期持续时间占整个生长期的比例较小,约1/4,但此期间内生长量占总生长量的比例却较大(苗高和地径均在57%以上),即在1/4 的生长期内完成了全年一半以上的生长量;并表现出苗高生长速生期在前、地径生长速生期在后的异速生长现象,苗高生长比地径生长先进入速生期,有可能是因为苗木不同生长时期有不同生长中心[26],这也可能与高生长为争得上方光源以增加苗木之间的竞争有关[27]或优先投资高生长而抢占有限生存空间[25]。但也有研究报道这种异速生长现象地径生长早于苗高生长[24,27-28]或差异不大的[19]。此外,速生期持续时间绝大多数种群表现为苗高生长比地径生长短,速生期苗高的生长速度增加较快,而地径的相对缓慢。在泡桐、杨树等苗期生长研究中也发现苗高生长速生期的持续时间比地径生长短20 ~30 d[24,27-28]。
速生期起始时间与结束时间相关性分析表明,苗高、地径生长的速生期起始时间和结束时间呈显著正相关,即进入速生期的时间早,完成速生的时间也早,迅速的生长能优先抢占有利空间,具有更强的竞争力,保证种群的生长、更新。本研究较早进入速生期的是纬度较高的四川马尔康种群和四川理县种群,这2 个种群分布于东北部紧接油松分布的种群,该分布区域已证实是高山松的古老起源中心[21],且该区域可能是一个避难所,古老的多态性得以保存[29-30],遗传组成复杂,多样性丰富,在新生境下体现出适应性较强,能快速地完成速生生长,但在有限的资源供给下,高生长较早进入速生期,优先投资高生长而抢占有限生存空间[24],满足苗高生长后,地径生长随之进入速生期,当苗高生长完成速生期生长后变缓时,地径生长进入速生期,此时无苗高生长的竞争,地径生长延长速生期持续时间,从而获得更多的生长量。不同种群苗木光合特性、水分利用效率等有所不同,且受产地气象因子的影响。因此,本研究需进一步从生理、气象等方面探讨,以揭示造成这种差异和相关性的深层次原因。
[1] 中国科学院中国植物志编辑委员会. 中国植物志:第7 卷[M].北京:科学出版社,1978:253 -255.
[2] 吴中伦.中国松属的分类与分布[J]. 植物分类学报,1956,5(3):131 -167.
[3] Wang X R,Szmidt A E. Evolutionary analysis of Pinus densata(Masters),a putative Tertiary hybrid. 2. A study using speciesspecific chloroplast DNA markers[J]. Theoretical and Applied Genetics,1990,80:641 -647.
[4] Wang X R,Szmidt A E. Hybridization and chloroplast DNA variation in a Pinus species complex from Asia[J]. Evolution,1994,48:1020 -1031.
[5] Wang X R,Szmidt A E,Savolainen O. Genetic composition and diploid hybrid speciation of a high mountain pine,Pinus densata,native to the Tibetan plateau[J]. Genetics,2001,159:337 -346.
[6] Song B H,Wang X Q,Wang X R,et al. Maternal lineages of Pinus densata,a diploid hybrid[J]. Molecular Ecology,2002,11(6):1057 -1063.
[7] Song B H,Wang X Q,Wang X R,et al. Cytoplasmic composition in Pinus densata and population establishment of the diploid hybrid pine[J]. Molecular Ecology,2003,12(11):2995 -3001.
[8] Liu Z L,Zhang D,Hong D Y,et al. Chromosomal localization of 5S and 18S-5.8S-25S ribosomal DNA sites in five Asian pines using fluoresence in situ hybridization[J]. Theoretical and Applied Genetics,2003,106(2):198 -204.
[9] Liu Z L,Zhang D,Wang X Q,et al. Intragenomic and interspecific 5S rDNA sequence variation in five Asian pines (Pinus)[J].American Journal of Botany,2003,90(1):17 -24.
[10] Ma Xiaofei,Szmidt A E,Wang Xiaoru. Genetic structure and evolutionary history of a diploid hybrid pine Pinus densata inferred from the nucleotide variation at seven gene loci[J]. Molecular Biology and Evolution,2006,23:807 -816.
[11] 毛建丰,李悦,刘玉军,等. 高山松种实性状与生殖适应性[J].植物生态学报,2007,31(2):291 -299.
[12] 郑益兴,冯永刚,彭兴民,等. 印楝1年生苗木生长节律与数量分级标准[J]. 南京林业大学学报:自然科学版,2008,32(3):25 -30.
[13] 彭方仁,黄宝龙.四川大叶樟一年生播种苗年生长规律的初步研究[J].南京林业大学学报,1989,13(4):1 -8.
[14] 莫惠栋.Logistic 方程及其应用[J]. 江苏农学院学报,1983,4(2):53 -57.
[15] 张连翔,刘学增.逻辑斯蒂曲线上的两个重要特征点的分析及其应用[J].河北林学院学报,1992,7(2):15 -158.
[16] 李秋元,孟德顺.Logistic 曲线的性质及其在植物生长分析中的应用[J].西北林学院学报,1993,8(3):81 -86.
[17] 魏巍,李根前,李甜江,等. 不同灌水强度下中国沙棘苗木生长过程[J].林业科学研究,2012,25(3):317 -324.
[18] 赖文胜.长序榆一年生播种苗的年生长规律[J].南京林业大学学报:自然科学版,2001,25(4):57 -60.
[19] 洑香香,方升佐,汪红卫,等.青檀一年生播种苗的年生长规律[J].南京林业大学学报:自然科学版,2001,25(6):11-14.
[20] 赵丹宁,熊耀国,宋露露,等. 泡桐无性系苗期年生长动态分析[J].林业科学研究,1993,6(1):39 -45.
[21] Wang Baosheng,Mao Jianfeng,Gao Jie,et al. Colonization of the Tibetan Plateau by the homoploid hybrid pine Pinus densata[J]. Molecular Ecology,2011,20(18):3796 -3811.
[22] 麻文俊,王军辉,张守攻,等. 楸树无性系苗期年生长参数的分析[J].东北林业大学学报,2010,38(1):4 -8.
[23] 杜坤,王军辉,贾子瑞,等. 不同种源砂生槐种子及幼苗生长变异研究[J].西北林学院学报,2012,27(4):144 -148.
[24] 傅大立,杨绍彬,丛培生,等. 泡桐苗期年生长参数的分析研究[J].林业科学研究,2001,14(3):301 -306.
[25] 吴国欣,王凌晖,俞建妹,等. 降香黄檀幼苗年生长节律研究[J].浙江林业科技,2010(3):56 -60.
[26] 骆绪美,苗婷婷,方建民,等. 乌桕1年生播种苗生长规律及育苗技术[J].林业科技开发,2010,24(5):114 -116.
[27] 姜成平,孙家京,隋日光,等.L35 等黑杨新无性系苗期生长规律研究[J].山东林业科技,2006(4):23 -24.
[28] 庞世龙,黄志玲,郝海坤,等. 大叶栎苗期生长模型及分析[J].林业科技开发,2011,25(5):36 -39.
[29] Lehmkuhl F. Extent and spatial distribution of Pleistocene glaciations in eastern Tibet[J]. Quaternary International,1998,45/46:123 -134.
[30] Frenzel B,Bräuning A,Adamczyk S. On the problem of possible last-glacial forest-refuge-areas within the deep valleys of eastern Tibet[J]. Erdkunde,2003,57(3):182 -198.
