水曲柳优树子代测定与家系选育技术1)
2013-03-05郭睿达曾凡锁詹亚光何之龙
郭睿达 曾凡锁 詹亚光 李 博 何之龙
(东北林业大学,哈尔滨,150040)
张桂芹 姚盛智
(黑龙江省帯岭林业科学研究所)
魏志刚
(林木遗传育种国家重点实验室(东北林业大学))
水曲柳(Fraxinus mandshurica)为木樨科白蜡属高大乔木,是东北林区的主要造林树种之一,是阔叶树中生长较快、适应性较强、经济价值较高的优良珍贵树种[1],属国家三级重点保护植物。水曲柳材质坚硬,纹理通直,富弹性,花纹美观;可供建筑、飞机、家具、室内装饰等广泛应用,是著名的军事用材和高级家具用材,也是制胶合板的良好原料[2-3]。随着人类对其的开发,水曲柳逐渐被广泛用作行道树和园林绿化树种,它成为具有较高景观价值的树种[4],由于近年来的过度采伐,导致水曲柳资源日益减少[5]。加强水曲柳的子代测定和优良家系选育研究,对于水曲柳优良资源的利用,促进人工林的良种化具有重要的推动作用。
生物量是评价森林生态系统结构与功能的重要指标;是关于生态系统单位面积上的生物有机体干物质量[6]。过去对于水曲柳的生物量性状的研究多是从宏观角度对一定区域的树木的蓄积量方面的研究,而且多数是从生物量与环境、生长条件等因素的关系方面来研究[7-11]。将水曲柳的生物量性状应用于子代测定和优树选择方面的研究未见报道。
生长量性状是树木家系选育最主要的性状之一,因为生长量性状反映了植株对特定环境的适应能力。但树木的生长量性状仅仅是作为比较直观的生理指标来反映其在特定环境下的外观生长状况。从碳汇角度看,生物量更能揭示碳储量的水平[12-14]。为了能够更有效地选育优良子代,深入地反映树木的生长发育能力,本试验将生物量性状和生长性状共同作为优良家系选择的依据来对水曲柳优树子代家系进行综合选择,旨在评价和选择优良家系,建立准确、高效的选育技术。
1 试验地概况
试验地位于黑龙江省伊春市的带岭区。帯岭地区位于北纬46°50' ~47°20'。带岭区地处中温带,属大陆性湿润季风气候。年气温变化较大,全年平均气温为1. 4 ℃左右;月平均最低气温为-19. 4℃;年最低气温天气在1月份,最低可达-40 ℃。月平均最高气温为20.9 ℃,年最高气温天气在7月份,最高可达37 ℃。降水期全年为130d 左右,多集中在7、8、9月份,占全年降水量的一半以上。帯岭地区土壤为山地暗棕色森林土,土壤全剖面呈暗棕色。
2 材料与方法
试验材料为水曲柳优树自由授粉子代的9年生幼树;共53 个家系。试验地中的水曲柳子代林共分5 个区组,各区组中的每个家系为一个小区(包括10 个单株,双行排列)。
对水曲柳子代测定林的所有家系的树高、胸径和地径进行实地测量,3 次重复,以当地种源作对照,并通过树高和胸径计算其材积。材积按平均试验形数法计算,公式为V = (h +3)g1.3fэ= (π/4)(d1.3)2(h+3)fэ,其中,h 为树高;d1.3为胸径;g1.3为树高1.3 m 处的横断面积;水曲柳的平均试验形数fэ为0.40[15]。
水曲柳生物量性状的调查:通过对上述生长性状数据的分析,对所有参试家系(包括对照)按照材积生长量水平从高到低进行排序。根据排序的结果,分别从高、中、低生长量选出3 个家系,加上当地对照共10 个家系。每个家系选取3 个单株,共30株进行树干解析。分别按1.3 m 和整米的树干高度截取圆盘、分类采集不同部位的样品,样品包括树干去皮圆盘、主根圆盘、主干茎皮、叶、枝和侧根(包括直径(D)D <2 mm、2 mm≤D <5 mm、D≥5 mm 的根)。烘干后,进行干质量的测定;求得每个单株的每个部位的含水率,以整株及每个部位的总鲜质量乘以相应的含水率,得到每个单株及其各个部位的生物量。每个家系的3 次重复的平均值作为该家系的生物量值。
水曲柳子代生物量与生长量之间的回归方程的建立:利用所选的10 个家系30 个单株的生物量和生长量调查结果,建立以生长量为自变量、生物量为因变量的回归方程。
通过SPSS 软件进行方差和各性状间的相关性分析与计算,进一步估算下列遗传参数:
各性状遗传变异系数:CV,f=(σf/X)×100%,式中,CV,f为遗传变异系数;σf为遗传标准差;X 为总体平均值。
各性状遗传增益:ΔG =(R/X)×100%,R 为选择响应,R=Sh2或R=iσph2,式中:S 为选择差;X 为总体平均值;h2为遗传力;i 为选择强度;σp为表型标准差。其中,h2=(式中,σ2g为遗传方差;σ2e为环境方差。S =x -X,x 为入选家系的总体平均值)[16-18]。
3 结果与分析
3.1 水曲柳子代林生长量性状变异分析
3.1.1 水曲柳优树子代家系生长性状调查结果
54 个家系(包括当地对照)的生长量调查结果表明,家系间树高的变异幅度为1.21 ~3.5 m,平均值为2.39 m;胸径的变异幅度为0.94 ~2.98 cm,平均值为1.78 cm;地径的变异幅度为2.18 ~5.83 cm,平均值为3.52 cm;单株材积的变异幅度为1.16 ~18.1 cm3,平均值为5.95 cm3。平均值分别超出对照种源的树高(2.19 m)9.13%、胸径(1.66 cm)7.23%、地径(3.36 cm)4.76%和材积(5.23 cm3)13.77%。方差分析结果(表1)表明,参试的54 个水曲柳家系(包括当地对照)间的树高、胸径、地径和材积差异均达到极显著水平。说明水曲柳子代家系间在生长性状上存在广泛的变异。
3.1.2 各生长性状间的相关性分析
各生长性状的相关性分析结果见表2。从表2中可看出,树高、胸径、地径和材积4 个生长性状之间呈正相关,且每两个生长性状之间的相关系数均在0.820 以上,其中材积与其他生长性状之间的相关系数均在0.900 以上,尤其是材积与胸径的相关系数,达到了0.982。这说明参试家系的各生长性状间具有很高的相关性。
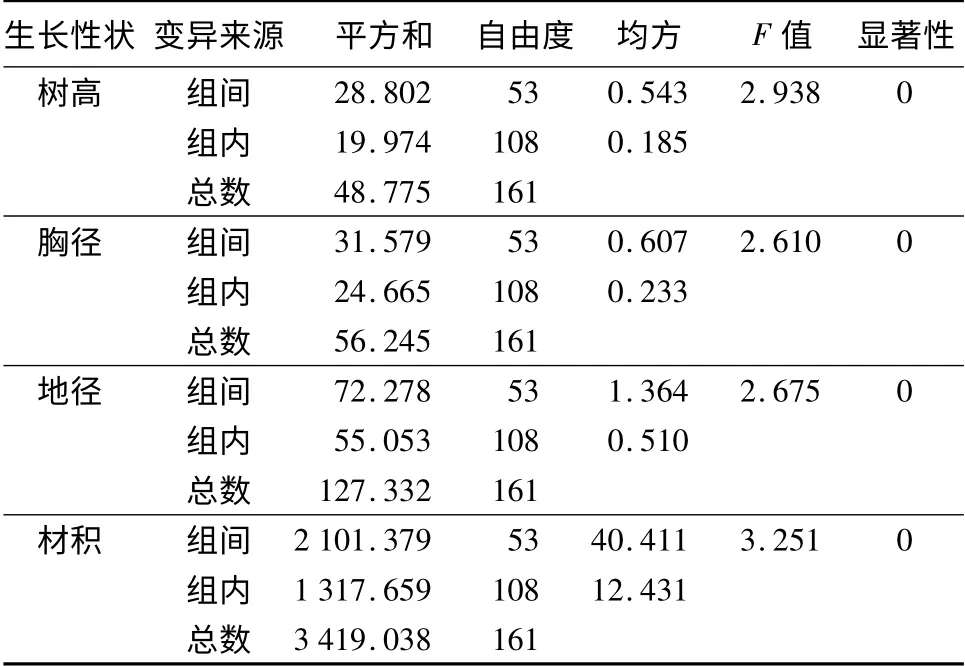
表1 生长性状方差分析

表2 水曲柳4 个生长性状之间的相关系数
3.1.3 遗传参数分析
计算了树高、胸径、地径、材积4 个生长量指标的家系遗传力和家系变异系数,并按照20%的入选率估算遗传增益。结果见表3。如表3所示,材积的家系遗传力最大,为69.24%;树高的家系遗传力为65.93%;胸径的家系遗传力为61.61%;地径的家系遗传力为62.61%。各生长性状的家系遗传力的变动范围在61.61% ~69.24%,说明参试家系的各生长性状受中度以上的遗传控制。材积的遗传变异系数为38.98%,遗传增益为23.45%,均高于其他生长性状的遗传变异系数和遗传增益。说明用材积作为优良家系选择的依据,其选择潜力大于树高、胸径和地径。遗传增益由大到小的顺序为材积、胸径、地径、树高,这4 个指标的遗传增益排序结果与遗传变异系数排序一致,证实水曲柳家系在生长性状方面具有丰富的遗传变异,具有较大的家系选择潜力和潜在的遗传增益。
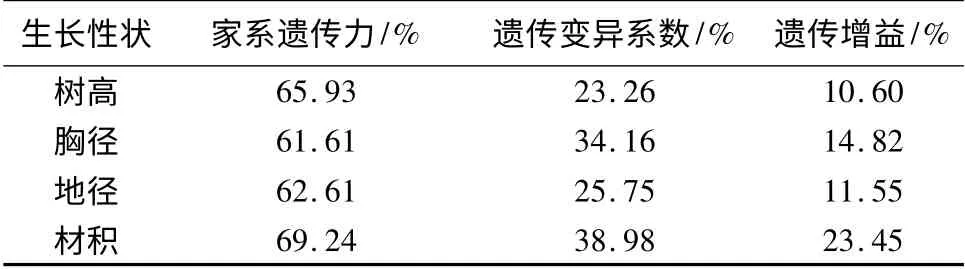
表3 各生长性状的遗传参数
3.1.4 根据生长性状进行家系选择
水曲柳生长初期以高生长为主,材积生长量又是反映树木生长量的综合指标,上述的各生长性状的相关性分析结果又表明各生长性状之间具有很高的正相关性。所以本试验用树高和材积2 个指标对水曲柳子代家系进行综合选择,将树高和材积2 个指标均优良的家系作为生长性状优良家系。
为便于优良家系的选择,将树高和材积高于所有家系的总平均值和高于对照种源的家系数目按照高出的百分比分别列出(表4、表5)。从表4、表5中可看出,材积高于总体平均值的家系有21 个,其中高出总平均值10%以上的有20 个;材积高于当地对照种源的家系有23 个,其中高出当地对照种源10%以上的有21 个。树高高于总体平均值的家系有21 个,其中高出总体平均值10%以上的家系有16 个;树高高于当地对照种源的家系有33 个,其中高出对照种源10%以上的家系有20 个。

表4 材积高于总体平均值和对照种源的家系数按分别高出平均值和对照的百分比的分布情况

表5 树高高于总体平均值和对照种源的家系数按分别高出平均值和对照的百分比的分布情况
按材积从大到小进行优树家系排序,其顺序为14 号、70 号、41 号、39 号、76 号、10 号、35 号、53 号、47 号、2 号、57 号、52 号、54 号、26 号、50 号、12 号、7号、27 号、59 号、21 号、42 号、55 号、对照、20 号、40号、6 号、1 号、17 号、56 号、63 号、3 号、46 号、74 号、28 号、77 号、75 号、45 号、71 号、38 号、62 号、73 号、64 号、36 号、29 号、13 号、68 号、65 号、61 号、44 号、72 号、8 号、60 号、51 号、69 号。
按20%的选择强度,将材积生长量高于总体平均值和对照10%以上的前10 位家系初步选为材积性状优良家系,前10 位家系为:70 号、14 号、39 号、10 号、41 号、76 号、35 号、53 号、47 号和2 号。
按照树高从大到小进行家系排序,其顺序为:14号、39 号、70 号、10 号、47 号、35 号、53 号、41 号、54号、57 号、50 号、76 号、59 号、2 号、7 号、55 号、12号、26 号、52 号、42 号、27 号、56 号、20 号、40 号、46号、63 号、6 号、1 号、21 号、3 号、38 号、64 号、74 号、对照、17 号、62 号、77 号、75 号、45 号、29 号、73 号、71 号、72 号、36 号、61 号、13 号、65 号、44 号、8 号、28 号、68 号、60 号、51 号、69 号。
同样按照20%的选择强度,初步选出的树高生长量高于总体平均值和对照10%以上的前10 位家系为:14 号、39 号、70 号、10 号、47 号、35 号、53 号、41 号、54 号和57 号。
综合树高和材积这2 个生长性状,高出各自总体平均值和对照种源10%以上的前10 位家系为70号、14 号、39 号、10 号、41 号、35 号、53 号和47 号。其中14 号家系的树高和材积均最高,其树高为3.5 m,胸径为2.98 cm,地径为5.39 cm,材积为18.1 cm3。树高高出参试家系总体平均值46.44%,高出对照种源59.82%;其材积高出所有参试家系的总体平均值达到204.62%,高出对照种源246.08%。入选家系中树高相对最低的41 号家系,其树高也高出总体平均值20.08%,高出对照种源31.05%;入选家系中材积相对最低的47 号家系,其材积高出总体平均值54.14%,高出对照75.22%,因此,按树高和材积生长量综合选出这些家系显示出明显的生长优势。
3.2 水曲柳生物量性状变异分析
3.2.1 不同部位生物量及整株生物量之间的相关性分析
根据上述的生长性状分析结果,生长量低于总体平均值和对照10%以上且排序在后10 位的家系作为生长性状较差的家系;其余排序在中间位置的33 个家系作为中等水平家系。分别选取优、中、差3个等级各3 个家系(70 号、39 号、10 号、52 号、36号、21 号、13 号、60 号、68 号),每个家系3 株样木以当地种源3 株样木作对照,对共计30 株样木进行树干解析。取标准样烘干后称质量,获得含水率,进而计算出各部位的干质量生物量。进一步分析整株生物量与各部位生物量之间的相关性(表6)。相关性分析结果表明,整株生物量和其他部位的生物量之间呈正相关,且相关性系数比较高。整株生物量和叶生物量之间的相关系数达到0.932,和主干生物量之间的相关系数达到0.977,和根生物量之间的相关系数达到0.974;整株生物量和枝生物量之间的相关系数相对最小,也达到了0.763。这说明整株生物量和各部位的生物量之间存在很高的正相关性。所以,可以仅用整株生物量来作为衡量参试家系生物量水平的指标。
3.2.2 回归方程的建立
根据所选的10 个家系(包括对照种源)共30个单株的生长量和单株生物量,分别以树高(H)、胸径(D1.3)、地径(D0)、D20H、D21.3H、D21.3D20H 为 自 变量,与单株生物量进行回归分析。在回归分析中选用线性函数、对数函数、倒数函数、二次函数、三次函数、复合函数、幂函数多种回归模型[19-21];在每一种自变量与单株生物量所建立的各种模型的回归方程中,选择相关系数R2最高,回归方程与回归系数均通过0.01 的显著性检验的方程作为备选回归方程。从各个自变量与单株生物量所建立的各种回归模型的回归方程中筛选出各备选回归方程(表7)。从表7中可看出,在筛选出的单株生物量与各个自变量的回归方程中,自变量D21.3D20H 与单株生物量的拟合效果最好,相关系数R2最高(R2=0.934),F 值最大,经检验达到了极显著水平(P <0.001),被选为最优的生物量回归模型,其函数式为W =3.456D21.3D20H-0. 001(D21.3D20H)2+ 642. 15(W:单株生物量)。

表6 不同部位生物量及整株生物量之间的相关系数

表7 初步筛选出的生长量与单株生物量的回归方程
3.2.3 水曲柳优树子代家系的生物量
根据建立的生物量与生长量之间相关关系的回归方程,求得了其他家系的单株生物量。各家系以选取的3 个单株生物量的平均值作为该家系的生物量水平。所有参试家系的单株生物量的变异幅度为456.52 ~3 337.34 g;参试家系单株生物量的总体平均值为1 109.33 g;当地对照种源的单株生物量为1 208.11 g。方差分析结果(表8)表明,所有参试家系的单株生物量的差异极显著,说明水曲柳子代家系间在生物量性状上存在明显变异。

表8 生物量性状方差分析
3.2.4 遗传参数分析
通过计算得出,生物量性状的家系遗传力为71.65%,家系变异系数为38.51%;根据20%的入选率,得出遗传增益为21.06%。结果表明,生物量性状的家系遗传力、家系变异系数和遗传增益均比较大。说明水曲柳优树子代家系间生物量受较高强度的遗传控制,且家系间生物量具有丰富的变异,证明以生物量性状作为良种选育指标,选育潜力比较大。
3.2.5 根据生物量性状进行优良家系选择
生物量高于总体平均值和对照种源的家系与总体平均值和对照种源的比较情况如表9所示。根据单株生物量进行家系比较,结果表明,单株生物量高于总体平均值的家系数有18 个,其中高出总体平均值10%以上的有11 个家系。单株生物量高于对照种源的家系数有11 个,其中高出对照种源10%以上的有10 个家系。将参试的54 个家系(包括对照)按照单株生物量从大到小的顺序进行了排序,其顺序为:70 号、14 号、39 号、10 号、41 号、76 号、52号、35 号、53 号、47 号、2 号、对照、54 号、57 号、7号、59 号、27 号、50 号、26 号、12 号、55 号、21 号、42号、20 号、13 号、3 号、40 号、46 号、1 号、6 号、17 号、56 号、36 号、77 号、74 号、28 号、75 号、71 号、45 号、63 号、64 号、62 号、38 号、29 号、73 号、72 号、61 号、60 号、44 号、65 号、8 号、68 号、51 号、69 号。同样按20%选择强度,将生物量排序在前10 位,且单株生物量高于所有参试家系(包括对照)的生物量总体平均值和对照种源的家系选为生物量性状优良的家系。根据以上结果,70 号、14 号、39 号、10 号、41号、76 号、52 号、35 号、53 号、47 号家系入选。其中70 号家系的单株生物量最大,其单株生物量为3 337.34 g。生物量最大的70 号家系的生物量高出所有参试家系总体平均值200.84%,高出对照种源176.24%;位于第10 位的47 号家系也高出总体平均值28.25%,高出对照种源17.76%。

表9 生物量高于总体平均值和对照种源的家系数按分别高出平均值和对照的百分比的分布情况
3.3 以生长量、生物量为指标进行优良家系的综合选择
本研究中,综合生长性状和生物量性状选出了9 个生长性状和生物量性状均优良的家系,这些家系为70 号、14 号、39 号、10 号、41 号、76 号、35 号、53 号和47 号。取材用于生物量测定的9 个水曲柳优树子代家系中,多数家系在分别按照生长量水平和生物量水平的排序中,排序位置浮动不大。其中生长量水平优良的70 号、39 号和10 号家系,也同样入选为生物量水平优良的家系;生长量水平较差的60 号和68 号家系,其生物量水平也较差。为了深入探讨以生长量和生物量为指标进行优良家系综合选择的方法,进一步分析了生长量性状与单株生物量之间的相关性。相关性分析结果表明,单株生物量与树高的相关系数为0.829,与胸径的相关系数为0.897,与地径的相关系数为0.918,与材积的相关系数为0.951。单株生物量与各生长性状之间的相关系数均在0.820 以上;尤其与地径和材积之间的相关系数均在0.900 以上。从而得出,单株生物量与各生长性状之间呈正相关,且相关系数较高。说明生长性状与生物量性状之间有较高的相关性,可以用生长量性状代替生物量性状进行家系选择。
4 结论与讨论
对于生物量指标的测定,本试验采用了回归方程法。按参试的54 个家系优中差三级取样木建立生长量和生物量的回归方程,推算获得的生物量值经检验,多数家系的排序与生长量排序位置变动不大,说明所建立的回归方程能较为有效地进行生物量变异分析。建议当参试家系比较多,进行全部树木解析困难的情况下,采用回归方程法用于生物量测定是一种可行的方法。
本试验选择出的8 个生长性状优良的家系,全部被选为生物量性状优良的家系,进一步证明了本试验中生长性状与生物量性状之间有很高的相关性。在通过相关性分析的结果证明生长性状与生物量性状之间相关性高的情况下,可以用生长性状代替生物量性状来进行水曲柳优树的子代测定和家系选择工作,而选择的结果可作为这两种性状综合选择的结果。这样可以减少工作量和减少对树木的采伐。
对于幼龄期的树木,在树高、胸径、地径和材积这4 个生长性状中,胸径和地径比较容易造成测量的误差。而树高的测量不易造成误差。材积是能够综合体现树木生长量水平的性状。所以综合评价水曲柳幼龄树木的生长量水平时,应以树高和材积为主。
基于生长性状的子代测定和优树选择方面的研究前人曾做过很多。早在20 世纪六七十年代就有人做过这方面的研究,如南京林产工业学院树木育种组在1967—1975年进行了基于生长性状的杉木优树子代测定的研究[22]。近年来,林木子代测定和优树选择方面的研究日趋成熟和深入,开发了综合多种生理指标进行子代测定和多性状综合选择的研究,如郝明灼[23]综合生长性状和生物量性状进行了关于银杏杂交种的子代苗期测定和多性状综合评价的研究。但关于水曲柳的子代测定和优树选择方面的研究都是仅仅将生长量性状(树高、胸径、地径、材积)作为优树选择的依据,根据多种性状对水曲柳家系、种源等进行综合选择的研究未见报道[24-26]。而且关于多性状综合选择方面的研究多是以苗期的树木为研究对象,缺乏文中探讨的幼龄以及成年树木为对象的多性状综合选择的研究[27-29]。
本试验的参试家系数量较多,所以遗传参数分析结果能比较客观地反映水曲柳优树子代林的情况。生长性状和生物量性状的遗传变异系数均比较大,树高这一性状的遗传变异系数虽相对小,其遗传变异系数也达到了23.26%。说明水曲柳优树子代的生长性状和生物量性状都具有较大的遗传改良潜力。材积和生物量的遗传增益均比较大,分别达到了23.45%和21.06%,具有较大的选育潜力。本试验是从53 个参试家系中选出10 个优良家系,入选率约为20%;为了提高遗传增益,可降低入选率提高选择强度。本试验所建立的生长量和生物量的综合选择技术及其选出的优良家系,将为今后进一步的优良家系选育、良种扩大培育奠定很好的基础。
[1] 王金国,李尚友,朱传富,等. 水曲柳优树子代测定试验初报[J].林业科技,1999,24(5):1 -4.
[2] 尹立辉,孙亚峰.水曲柳研究进展和展望[J]. 长春大学学报,2006,16(3):72 -75.
[3] 荆涛,张林玉.水曲柳的生长规律[J]. 内蒙古农业大学学报,2004,25(1):36 -39.
[4] 吴增荣,秦勇强. 水曲柳引种育苗试验初报[J]. 浙江林业科技,1996,16(4):18 -20.
[5] 林士杰,张忠辉,谢朋,等.中国水曲柳基因资源的保护与利用[J].中国农学通报,2009,25(24):158 -162.
[6] 程堂仁,冯菁,马钦彦,等.小陇山油松林乔木层生物量相容性线性模型[J].生态学杂志,2008,27(3):317 -322.
[7] Poorter H,Nagel O. The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water:a quantitative review[J]. Australian Journal of Plant Physiology,2000,27(12):595 -607.
[8] Son Yowhan,Hwang Jaehong. Fine root biomass,production and turnover in a fertilized Larix leptolepis plantation in central Korea[J]. Ecological Research,2003,18(3):339 -346.
[9] 田宇明,王庆成.初植密度对10年生水曲柳人工林生物量及根系的影响[J].林业科学,2011,47(7):102 -107.
[10] 王向荣,孙海龙,余鑫,等. 坡向和坡位对水曲柳中龄林生长的影响[J].山西农业大学学报,2011,31(1):30 -34.
[11] Bond-Lamberty B,Wang C,Gower S T. Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba[J]. Canadian Journal of Forest Research,2002,32(8):1441 -1450.
[12] 王秀云,孙玉军,马炜.不同密度长白落叶松林生物量与碳储量分布特征[J].福建林学院学报,2011,31(3):221 -226.
[13] Homann P S,Remillard S M,Harmon M E,et al. Carbon storage in coarse and fine fractions of pacific northwest old-growth forest soils[J]. Soil Science Society of America Journal,2004,68(6):2023 -2030.
[14] Wang Jiashe. The analysis and assessment on the biomass and carbon sequestration of Chinese fir plantation ecosystem[J]. Journal of Fujian Forestry Science and Technic,2008,35(2):1-4.
[15] 陈晓阳,沈熙环. 林木育种学[M]. 北京:高等教育出版社,2005.
[16] 邓继峰,张含国,张磊,等.杂种落叶松F2 代自由授粉家系纸浆材遗传变异及多性状联合选择[J]. 林业科学,2011,47(5):31 -39.
[17] 张超,朱航勇,邓继峰,等.日本落叶松碳储量家系遗传变异及优良家系选择研究[J]. 中南林业科技大学学报,2011,31(10):53 -62.
[18] 陈岳武. 树木良种选育方法[M]. 北京:中国林业出版社,1984.
[19] 胥辉.一种生物量模型构建的新方法[J].西北农林科技大学学报:自然科学版,2001,29(3):35 -40.
[20] 郭永清,郎南军,杨旭,等. 膏桐人工林单木生物量回归模型研究[J].浙江林业科技,2009,29(6):35 -37.
[21] Case B,Hall R J. Assessing prediction errors of generalized tree biomass and volume equations for the boreal forest region of west-central Canada[J]. Can J For Res,2008,38(4):878 -889.
[22] 福建省洋口林场,南京林产工业学院树木育种组. 杉木优树后代测定及其遗传力的估计[J]. 遗传学报,1977,4(2):152-158.
[23] 郝明灼. 银杏杂交育种的初步研究[D]. 南京:南京林业大学,2004.
[24] 崔宝国,朴楚炳,王洪君,等. 水曲柳天然优良林分选择方法和标准的探讨[J].林业科技,1992,17(5):1 -3.
[25] Bird J A,Torn M S. Fine roots vs. needles:A comparison of 13C and 15N dynamics in a ponderosa pine forest soil[J]. Biogeochemistry,2006,79:361 -382.
[26] Fournier R A,Luther J E,Guindon L,et al. Mapping aboveground tree biomass at the stand level from inventory information:test cases in Newfound and Quebec[J]. Canadian Journal of Forest Research,2003,33(10):1846 -1863.
[27] 黄崇辉,杨朝辉,陈文平,等. 尾叶桉家系遗传力评估和选择研究[J].广东林业科技,2008,24(6):41 -45.
[28] Niklas K J,Enquist B J. Canonical rules for plant organ biomass partitioning and annual allocation[J]. American Journal of Botany,2002,89(5):812 -819.
[29] Maria J G,José L L,José C R. Does selecting for improved growth affect wood quality of Pinus pinaster in Portugal[J]. Forest Ecology and Management,2009,258(2):115 -121.
