香根草在荆州的引种和有性繁殖习性研究
2012-11-10朱桂才长江大学园艺园林学院湖北荆州434025
姚 振,朱桂才,赵 江,王 超 (长江大学园艺园林学院,湖北 荆州 434025)
香根草在荆州的引种和有性繁殖习性研究
姚 振,朱桂才,赵 江,王 超 (长江大学园艺园林学院,湖北 荆州 434025)
为给香根草(VetiveriazizanioidesL.)生理生态研究和应用提供依据,进行了香根草多个生态型的引种、分蘖繁殖、开花结实习性调查、花粉萌发、实生苗繁殖以及越冬调查等试验,获得了香根草分蘖和有性繁殖的详细资料,探讨了香根草开花和结实的影响因素,分析了结实率低的原因。
香根草(VetiveriazizanioidesL.);分蘖繁殖;花粉萌发;有性繁殖
香根草(VetiveriazizanioidesL.)是禾本科岩兰草属多年生草本植物,1958年开始进入中国,20世纪80年代以后,在华南、华东、西南等地香根草被广泛用于治理水土流失。香根草被世界上100多个国家和地区列为理想的保持水土植物,在治理污染、土工工程防护、鱼畜饲料、菌草、工艺编织品开发利用、造纸和燃料使用等方面有巨大应用潜力[1]。
近年来,香根草生态工程受到国内外的广泛重视并得到迅速的推广和应用,国内的研究主要集中在以下几个方面:香根草对淹水、干旱的适应性研究,光合生理生态研究[2-3];香根草生态工程研究,优良生态型的筛选[4];香根草组织培养、种子繁殖等繁殖技术及子代植株生物学特性研究[5-6]等方面。
香根草的引种栽培会大大改善生态环境,阻止生境的进一步恶化,湖北省近年开始了引种栽培,利用香根草治理环境[7]。但是,香根草的引种推广首先需要研究其在引入地的适应性、繁殖,确定其是否易扩散等问题,所以研究香根草在引种地的无性繁殖、有性繁殖以及越冬的问题,对其在引入地发挥价值具有重要意义。本研究在湖北中部城市荆州进行了多个生态型的香根草的引种、分蘖繁殖、开花结实习性调查、花粉萌发、实生苗繁殖以及越冬调查等试验,获得了香根草无性和有性繁殖的详细资料。在此基础上,重点探讨了香根草开花和结实的影响因素,分析了香根草结实率低的原因,以期为香根草的生理生态研究、繁殖和应用提供详细的资料。
1 材料与方法
1.1 材料
试验材料来源于不同的气候区域的13个生态型,2005~2006年种植于长江大学园艺园林学院实习基地。其中,生态型Lilongwe和Zomba来源于马拉维,马拉维处于非洲东南内陆,大部分地区为热带干湿季气候区的高地气候。生态型Kandy来自斯里兰卡,斯里兰卡(亚洲岛国)属热带季风性气候。Malaysia、Sabak bernam来自马来西亚,马来西亚属于热带雨林海洋性气候。Capital、Sunshine、Hoffman来自美国,美国属于温带大陆性气候。Monto来自大洋洲的澳大利亚,澳大利亚与东南亚太平洋地区气候相近。Wild、Cultivated、A、C 4个生态型来自中国广东,广东绝大部分地区都属于亚热带季风气候。引入地荆州,属亚热带季风气候区,光能充足、热量丰富、无霜期长。年平均气温15.9~16.6℃,冬季一月是最冷月,最低温度曾低于-10℃。
从越冬后的母体植株上取单个分蘖进行繁殖,为了便于区别不同龄的分蘖丛,将单个分蘖形成的分蘖丛当年称为一年龄植株。一年龄植株经过1a的生长,越冬后继续萌发,第二年为二年龄植株,第三年称为三年龄植株,此后称为多年龄植株。
1.2 方法
1.2.1 分蘖繁殖
为了评估香根草分蘖繁殖的效果及其在引入地的适应性,记录所有植株3a的分蘖数、植株高度和基部周长。分蘖数指单个分蘖丛中所有分蘖的数目,植株高度指每株从基部到花序或最高叶片顶端的高度,基部周长指分蘖丛基部的周长。
1.2.2 有性繁殖习性
1)拔节与抽穗 A和C 2个生态型通过单个分蘖繁殖的植株,连续3a调查记录其拔节率和抽穗率。其他生态型的多年生植株连续3a调查记录抽穗率。拔节率=(拔节数/分蘖数)×100%,抽穗率=(抽穗数/拔节数)×100%,其中,拔节数指分蘖丛中出现节间伸长现象的分蘖数目,抽穗数指完成花序的形态分化的分蘖数目。
2)抽穗与开花 逐个记录每个花穗的穗展开时间、小花开始开放时间、开花结束时间,以及抽穗开花期间的温度及天气情况。
3)花粉离体萌发 生态型A开花期间收集花粉,进行离体萌发实验,设计培养温度和培养基2个处理。培养温度设置14、16、18、20、22℃5个水平。培养基包含4个水平,即A:蔗糖10g/L、Ca(NO3)2·4H2O 0.3g/L、H3BO30.1g/L、KNO30.1g/L、MgSO4·7H2O 0.2g/L;B:蔗糖10g/L、H3BO30.02g/L;C:蔗糖15g/L、H3BO30.02g/L;D:蔗糖20g/L、H3BO30.02g/L。
将培养基融化滴在载玻片上,冷却后将花粉用毛笔均匀刷在培养基上。将播种后的培养基放入培养箱中,每隔30min观察花粉萌发情况,2h后统计最终萌发花粉数,计算萌发率。
4)结实 逐个统计单穗结实的数量与空秕的数量,计算结实率。
5)实生苗 采收生态型A的种子,实验室催芽后,移栽幼苗种植于田间,记录实生苗生长情况。抽穗开花的生态型种植地周围不做干扰,次年调查种植地附近区域实生苗出现情况。
1.2.3 越冬调查
试验的第二年冬季,选择各生态型代表性植株,每隔2周于3个时间点(11月13日、12月1日、12月16日)调查计算绿叶比例,评估越冬情况。待地上部分全部枯黄后,离地50cm左右割去地上部分。第三年春季,记录返青情况。
2 结果与分析
2.1 分蘖繁殖的效果
A、C生态型经过3a的分蘖繁殖,株高平均值分别达到262.8cm和231.1cm。引入的其他生态型的第三年株高平均为241.9cm,其中生态型Wild最高株高可以达到360cm。从单棵分蘖苗到生长成丛,基部周长逐年上升。2个生态型第三年的分蘖数分别可以达到31.4株和33.1株,最高可以达到59株和61株。分蘖丛3a的基部周长最高可以达到66.88cm,2个生态型平均每年增加7.54cm和9.07cm。其他生态型在3a中也表现出基部周长的有限增加。
从株高、分蘖数和分蘖丛基部周长分析,引入的所有生态型表现了非常好的适应性,并没有因为与原产区环境差异而表现出不适应。香根草的分蘖属于疏蘖型,不会从栽植点向四周扩散,易于控制,生产中可以利用分蘖苗进行繁殖。
2.2 香根草一年龄、二年龄和多年龄植株有性繁殖特性的差别
图1是生态型A一年龄、二年龄和多年龄植株的拔节率和抽穗率的比较。由图1可以看出,拔节率在3a中出现波动。抽穗率一年龄和二年龄的植株大致相同,多年龄的抽穗率提高约2倍,抽穗率表现为升高的趋势。图2是生态型C一年龄、二年龄和多年龄植株的拔节率和抽穗率的比较。由图2可以看出,生态型C在3a中也表现了拔节率的波动,但是抽穗率变化不大。
引入的2个生态型分蘖繁殖的幼苗在当年便可以完成营养积累,转入生殖生长而抽穗开花。但是,2个生态型的抽穗率差别较大,但均在30%以下,只有一部分分蘖植株可以完成抽穗的过程。与生态型C的分蘖苗一直保持很低的抽穗率所不同的是,生态型A的分蘖苗抽穗率在第二年和第三年逐渐增加。
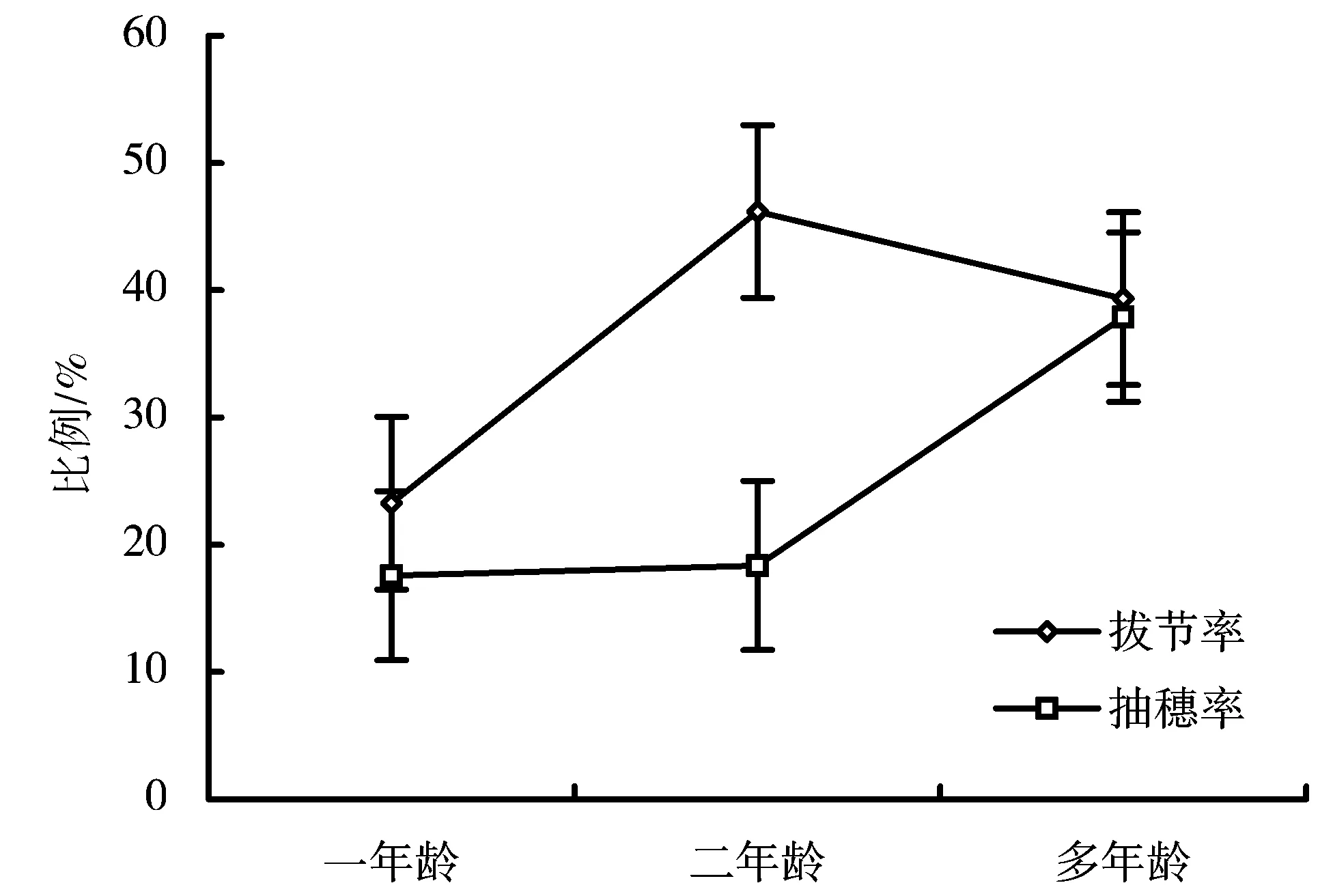
图1 生态型A不同龄植株的拔节率和抽穗率
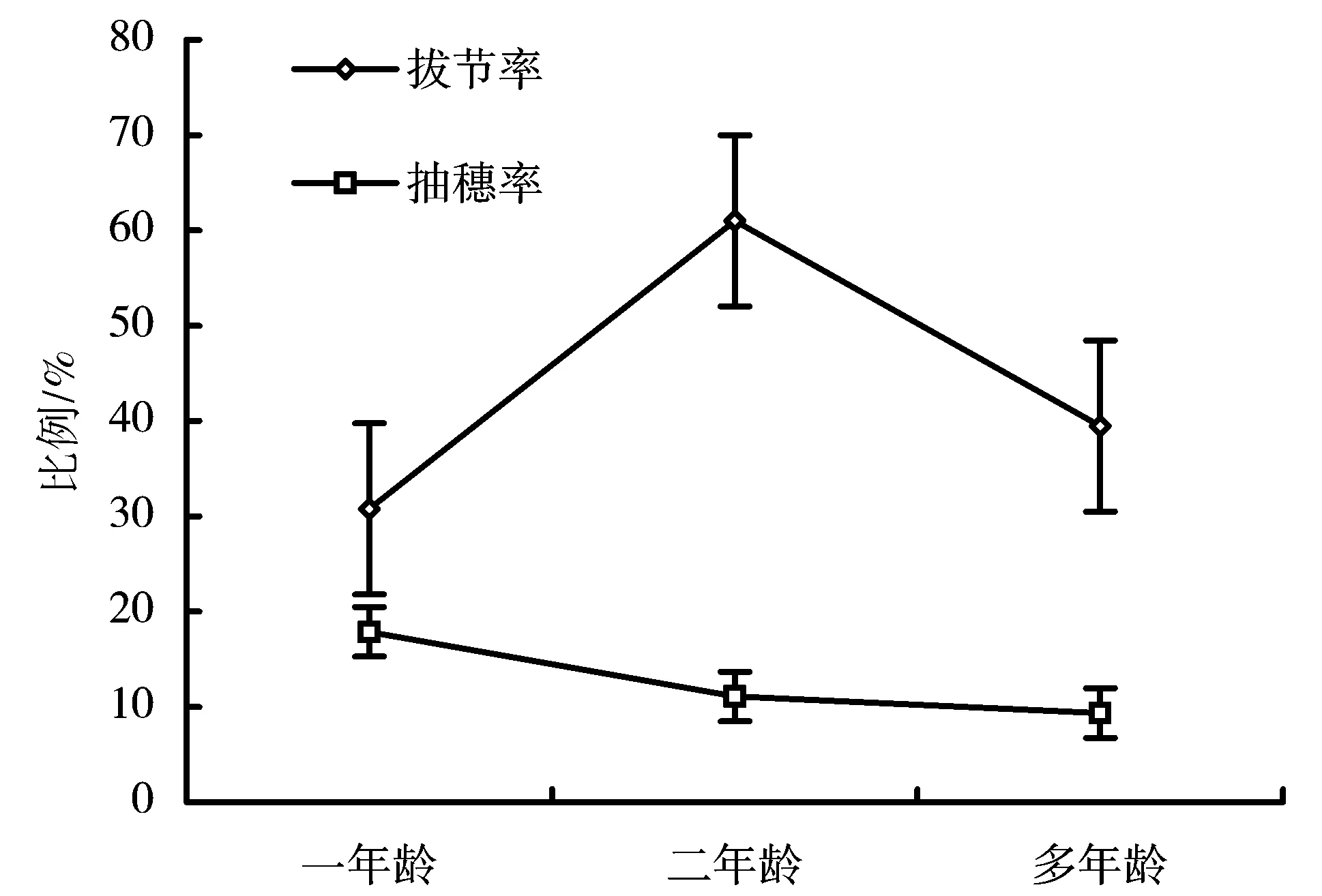
图2 生态型C不同龄植株的拔节率和抽穗率
图3是其他11个生态型3a抽穗率比较。从图3可以看出各生态型经过引种后连续几年呈现出不同的生长状况。抽穗率表现为上升、下降或者波动,并且这种趋势与引种来源地的关系不明显。来源于广东的4个生态型都有较高的抽穗率,其中生态型wild的抽穗率最高可以达到65.7%。生态型Kandy、Hoffman、Capital、Sunshine、Sabak bernam、Lilongwe抽穗率一直保持在很低的水平,甚至没有抽穗,这些生态型在整个试验期间都没有结实。
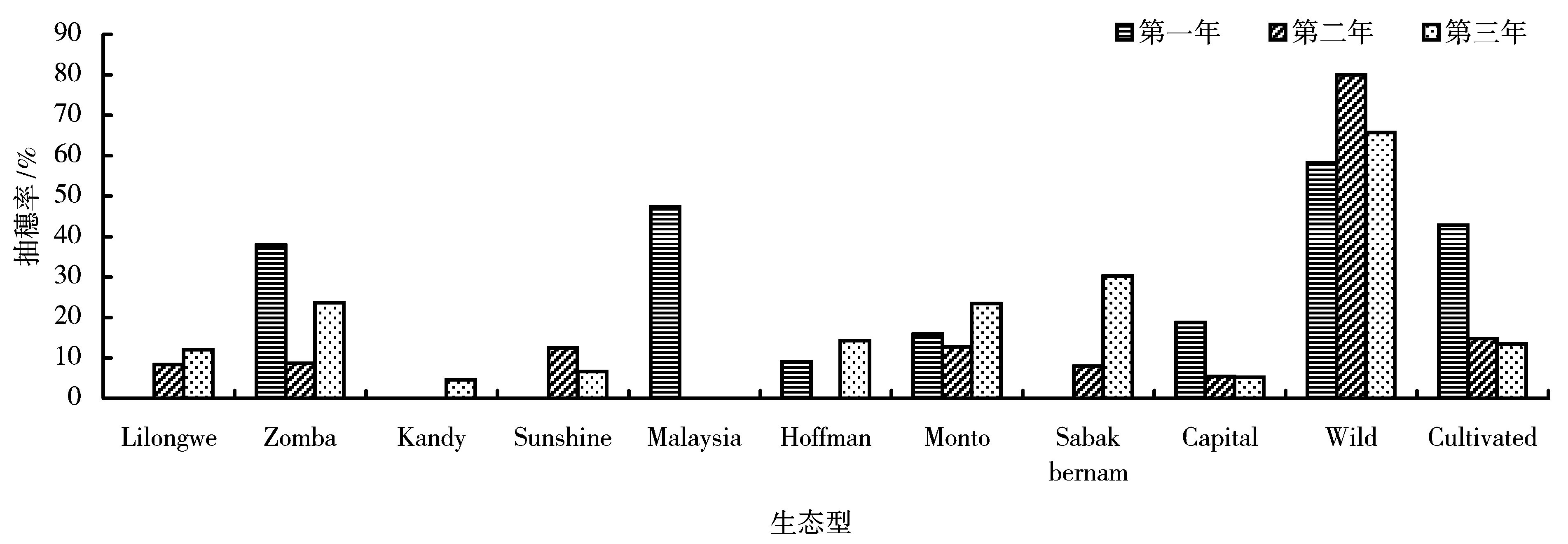
图3 11个生态型3a的抽穗率
2.3 温度、培养基对花粉离体萌发的影响
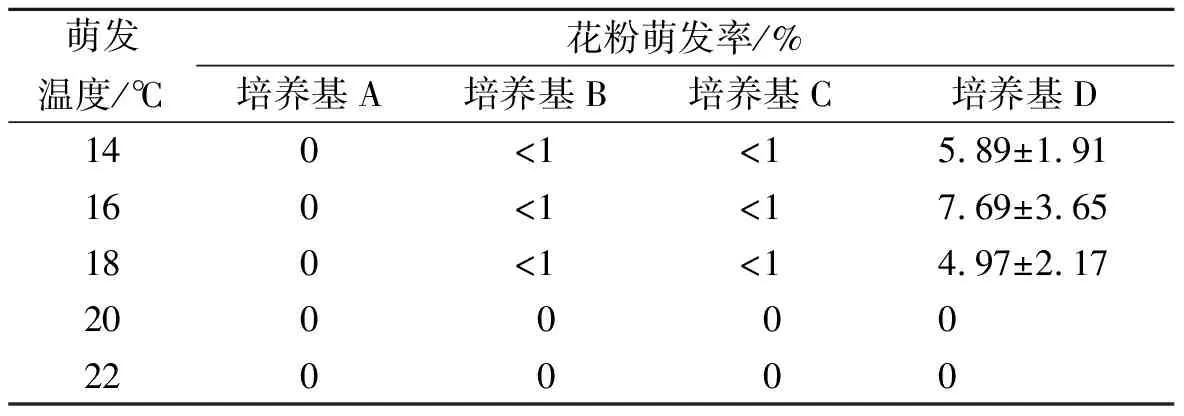
表1 温度和培养基条件对花粉萌发率的影响
香根草花粉在不同温度和培养基条件下的萌发率见表1。由表1可以看出,花粉在不同温度下的萌发率具有很大差异,在14~18℃下萌发数最高,20℃和22℃时花粉不萌发。在富含无机盐的A培养基下,14~22℃时花粉均不萌发。不同糖浓度(B、C、D培养基)下,萌发率在糖浓度达20%后萌发率才有所提高,但花粉萌发率仍还是很低,均小于10%。
2.4 气象条件对花序展开和开花的影响
通过逐个记录所有抽穗分蘖的几个重要时间(穗展开时间、小花开放时间和开花结束时间),研究温度条件对抽穗和开花的影响。为便于在图4和图5中将时间数据与期间的温度变化相对应,研究中引入T值,T为试验期间的时间刻度,即以9月1日的T值为1,9月2日的T值为2,然后类推至整个试验阶段。
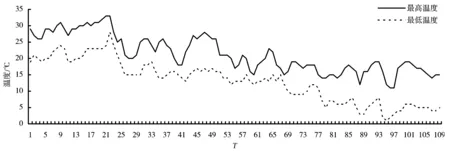
注:9月1日的T值为1,9月2日的T值为2,余类推,图5、图6同。T=25~29、55~68时为连续阴雨天气。
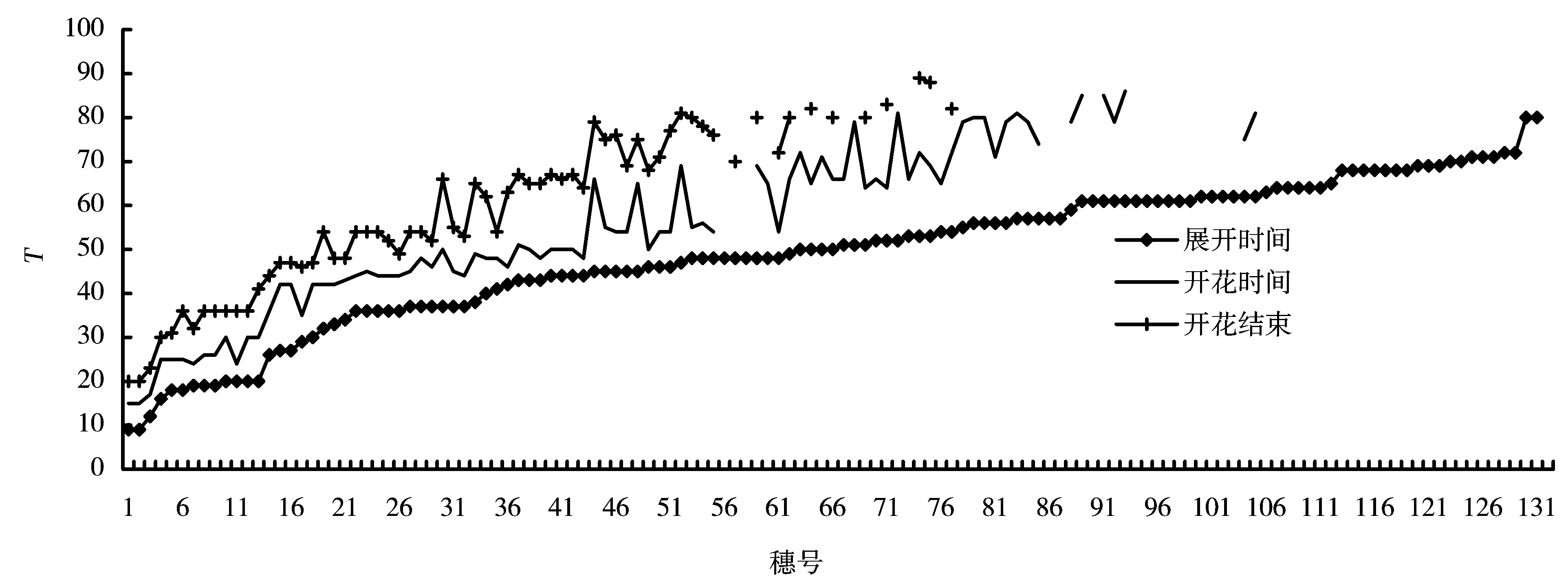
图5 香根草抽穗开花在时间上的分布
生态型A、C和Wild的抽穗率都能保持在很高的水平,但是并不是每一个抽穗分蘖的花序都能顺利地展开。从图5可以看出,第一个抽穗展开从9月9日(T=9)开始,11月19日(T=80)最后一个花穗展开,前后间隔71d。在11月19日后香根草的抽穗不再展开,但是可以观察到花序的形态分化已经完成。花穗的展开主要集中在10月5日(T=35)到11月9日(T=70),前期和后期只占很小的比例。根据图4给出的温度曲线,对应的花序展开的最低温度为12℃,温度低于12℃时花序不再展开。
圆锥状的花序展开后小花逐渐开放,共有约50个穗能够完成抽穗和开花的整个过程,一个花序上的小花开始开放到整个花序开花结束,平均时间为11.6d。前期的花穗开放最短时间为5d,后期的花穗最多可以持续25d。从图5的曲线可以看出,后期多数分蘖的抽穗只能完成穗的展开或部分花的展开,不能完成开花的过程。
小花开放从9月15日(T=15)开始,一直到11月29日(T=90)结束,前后间隔75d。小花开放开花主要集中在9月25日到10月20日,后期只有少数开花,而且一个花序中只有很少量的小花能够开放。根据图4给出的温度曲线,温度低于14℃小花就不再开放。
在T为25~29、55~68时试验记录为连续阴雨天气,这段时间的展开花穗不能开花。
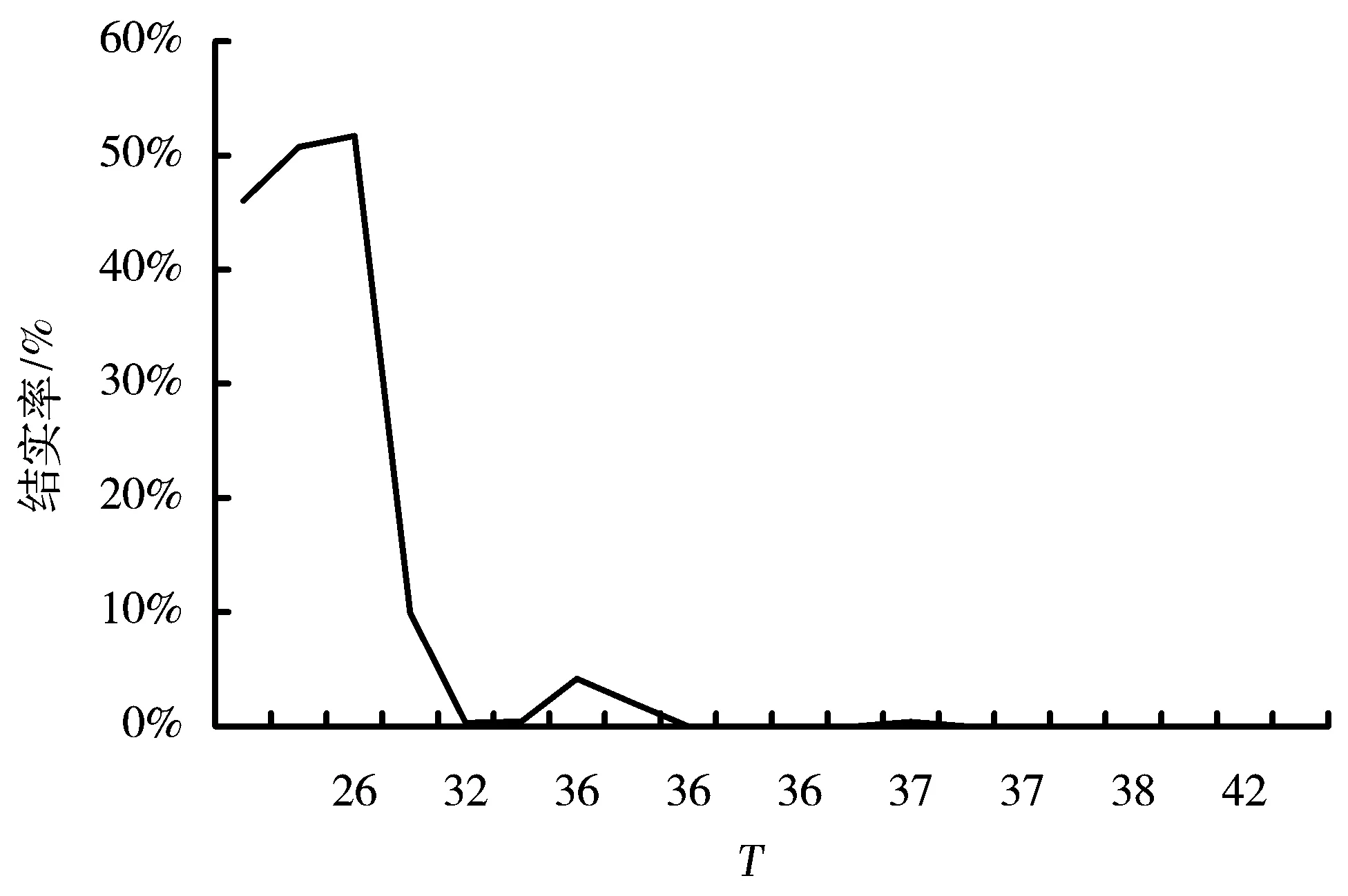
图6 抽穗时间对香根草结实率的影响
2.5 抽穗时间与结实率的关系
从图6可以看出,香根草结实主要集中在很短的时间内,抽穗时间T=42(10月11日)为临界点,以后香根草不再结实。结实的高峰期抽穗时间T在16~32之间,单穗最高结实率为51.7%,此时的最高温度在25℃左右,最低温度在15℃左右。抽穗时间T的最小值为9,9月9日到9月15日的少量抽穗由于脱落比较严重,无法统计,虽然可以观察到结实但结实率不高。
2.6 实生苗
生态型A采收的种子在实验室条件下育苗,幼苗移栽到大田。移栽的实生苗当年平均分蘖数为21.3株,第二年平均分蘖数为42.8株,这些分蘖丛的生长与通过分蘖繁殖的分蘖丛无明显差异。生态型A在开花结实的过程中,逐步有一些种子散落在四周。在试验的第三年,在距离香根草生态型A植株1m左右的位置发现了多颗实生苗。
2.7 香根草分蘖丛的越冬情况
从图7可以看出,Lilongwe、Zomba、Hoffman、Sabak bernam、Capital、A、C以及Kandy在其他生态型地上部分基本枯萎的情况下,绿叶比例仍然较高。生态型Malaysia和Cultivated地上部分枯萎的速度最快,抗寒性较差。各个生态型表现了抗寒性上的差异,各个生态型第二年均能正常萌发新的分蘖。
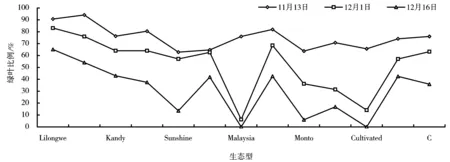
图7 不同生态型3个不同时间点的绿叶比例
3 讨论与结论
香根草可分为有性繁殖(能正常开花结实,如印度北部的一个野生种以及广东湛江发现的野生种)和无性繁殖(不开花或开花不结实,靠营养器官繁殖)[6,8]2大类,本研究选用的生态型包含了这2种类型。所有生态型都能在引入地正常生长,可以正常利用分蘖繁殖的方法,分蘖丛保持在原种植的范围内而不扩散。但是,这些生态型在开花结实习性上有很大的区别,来源国内的几个生态型能够有效地开花结实,而生态型Kandy、Hoffman、Capital、Sunshine、Sabak bernam、Lilongwe抽穗率一直保持在很低的水平,基本没有有效的抽穗结实。试验已经证实了散落种子可以萌发实生苗,这一点对于香根草的利用提出了要求,生产应用中最好选用没有有效抽穗的生态型。
研究结果表明,分蘖繁殖的幼苗当年抽穗开花会有延迟,2~3a后开花结实和多年生的分蘖丛相同。出现这种情况的原因可能是植株在分蘖繁殖后需要营养积累,在合适的情况下转入生殖生长。如果香根草在引入地作为牧草使用,反复的收割可能会抑制开花。
研究发现香根草花粉萌发率极低,并且离体萌发温度在14~16℃之间,这与其他禾本科植物花粉的萌发温度具有很大的差别。水稻、小麦、玉米花粉的离体最适萌发温度在28~30℃之间[9-11],显著高于实验测得的香根草的花粉离体萌发温度。本研究只对香根草花粉离体萌发的条件进行了初步摸索,如果将温度提高到28~30℃,或是提供其他的营养条件,花粉的萌发率是否提高,有待进一步研究。
研究发现,在引入地能正常结实的香根草生态型,结实率很低。有效结实穗集中在约2周内,这些穗的结实率高,而其他时间的穗基本不能结实。香根草抽穗开花集中在9月至10月,此时温度偏高,适合香根草花粉萌发的时间段很短,而花粉萌发对温度的要求较为苛刻,两者的矛盾可能导致了传粉和受精困难,进而导致了结实率低。同时,抽穗后期的低温造成有效的开花穗少,大量的抽穗都不能结实。可以推测,抽穗前期的高温和后期的低温都会抑制香根草结实。引入地秋冬季长时间的阴雨天气比较常见,这些天气影响了香根草抽穗的展开及花的开放,这也是香根草大量的抽穗结实率不高的重要原因。
致谢:感谢中山大学绿地中心提供试验材料。
[1]程 洪,李 斌.香根草的技术应用[J].江西科学,1998,16(3):204-210.
[2]刘金祥,王铭铭.淹水胁迫对香根草生长及光合生理的影响[J].草业科学,2005,22(7):71-73.
[3]刘 艳,黄乔乔,马博英,等.高温干旱胁迫下香根草光合特性等生理指标的变化[J].林业科学研究,2006,19(5):638-642.
[4]夏汉平,刘世忠.香根草优良生态型筛选研究[J].草业学报,2003,12(2):97-105.
[5]马镇荣,刘 卫,王昌虎,等.香根草体细胞胚胎发生的细胞学特征与形成条件[J].生态学报,2003,23(7):1290-1296.
[6]刘金祥,李文送,李红燕.种子繁殖香根草植株的生物学特性及病虫害初报[J].草业科学,2005,22(4):108-111.
[7]李爱华,赵 虎,伍仪章,等.香根草的引种及抗逆性栽培试验[J].湖北林业科技,2006,(5):19-22.
[8]何 敏,严力蛟,陈启银,等.香根草研究综述[A].徐礼煜.香根草研究与展望[C].北京:中国农业科技出版社,1998:147-152.
[9]陈士强,王 忠,刘满希,等.水稻花粉萌发及花粉管生长动态[J].中国水稻科学,2007,21(5):513-517.
[10]朱展望,张改生,牛 娜.小麦花粉的离体萌发研究[J].麦类作物学报,2007,27(1):12-15.
[11]王艳哲,崔彦宏,张丽华,等.玉米花粉活力测定方法的比较研究[J].玉米科学,2010,18(3):173-176.
10.3969/j.issn.1673-1409(S).2012.04.003
Q949.71+4.2
A
1673-1409(2012)04-S009-06
2012-02-22
长江大学科研启动基金项目(2005018)。
姚 振(1980-),男,湖北荆州人,博士生,讲师,研究方向为植物分子生物学与植物学。
