江苏烟粉虱生物型鉴定与危害程度分析
2012-06-09谭永安肖留斌季英华柏立新
谭永安,肖留斌,孙 洋,季英华,柏立新
(江苏省农业科学院植物保护研究所,南京 210014)
烟粉虱Bemisia tabaci(Gennadius)为世界性、多寄主害虫,不仅可以取食危害多种寄主植物,而且能够传播110多种植物病毒,给多种作物造成了严重的经济损失 (Jones D,2003)。作为一个处于快速进化过程的复合种,烟粉虱遗传特性极为复杂,可分为多种不同的生物型,到目前为止,已经命名的生物型至少有26个 (Delatte Het al.,2005)。国内研究结果表明:B型烟粉虱在我国广泛分布 (罗晨等,2005);Q型烟粉虱自2005年首次在云南昆明发现以来 (褚栋等,2005),随后在江苏、浙江、武汉等地区发现其危害 (徐婧等,2006;饶琼等,2009;沈媛等,2011),有报道表明Q型烟粉虱已在局部地区大量发生并造成严重的经济损失 (徐婧等,2006);沈媛于2005~2009年对江苏地区烟粉虱生物型演替进行了较为系统的研究,发现Q型烟粉虱在江苏地区发生频率逐年升高,并由苏中和苏南地区向苏北地区扩散,显示出逐渐取代 B型烟粉虱的趋势 (沈媛等,2011)。但是,由于不同生物型烟粉虱在寄主范围、寄主危害程度均存在较大差异 (Nombela G et al.,2011;Bonato O et al.,2007),因此,需进一步研究明确不同生物型烟粉虱的地理分布特性与当地主要寄主危害程度之间的关系,明确重点防治作物与关键防治时期,以优化区域性烟粉虱监测防控策略。本研究在前人的工作基础上,为进一步探讨不同生物型烟粉虱在江苏省不同农区的发生现状与发展趋势,明确江苏省不同地域、主要受害作物上烟粉虱的生物型种类,探明不同生物型烟粉虱的地理分布与当地主要寄主上发生程度及受害程度之间的关系。作者于2010年烟粉虱发生危害高峰期在江苏省10个市、25个地点对当地主要受害寄主上烟粉虱的发生危害情况进行了抽样调查,应用危害指数对不同作物上烟粉虱的发生危害程度进行了比较,同时通过mtDNA COI分子标记技术鉴定各地主要受害寄主上烟粉虱生物型,并在此基础上构建系统进化树,以期从宏观上进一步把握不同生物型烟粉虱在江苏省的区域性遗传分布与主要寄主种类,从而指导防治。
1 材料与方法
1.1 寄主范围及发生程度调查
于2010年烟粉虱发生高峰期,分别在江苏省苏南 (无锡、苏州、南京)、苏中 (南通、扬州)和苏北 (徐州、盐城、宿迁、连云港、淮安)等多个地区调查主要寄主植物上烟粉虱的虫口密度与危害情况。各地主要寄主植物上烟粉虱的虫口密度与危害情况调查田块均采用5点取样法,每点调查5株,每株分别选上、中、下部叶片各2片,轻轻翻转叶片,记录成虫数。同时采集叶片带回实验室镜检,统计烟粉虱伪蛹、若虫及卵的数量。对叶片密集且较难区分上、中、下部叶片的寄主植物,则分别取上部嫩叶和中下部老叶各3片作为镜检材料。对寄主植物上烟粉虱若虫和伪蛹的数量以叶为单位进行统计,参照国内外有关烟粉虱发生程度的标准,将其划分为4个等级:每叶小于10头的为1级,记为“+”;10~30头的为2级,记为“++”;30~50头的为3级,记为“+++”;大于50头的为4级,记为“++++”。
1.2 生物型鉴定
1.2.1 样品采集
于2010年春、秋两季在江苏省13个地理区域采集烟粉虱成虫。采样时轻轻翻转寄主叶片,用吸虫管收集成虫,成虫放入75%的乙醇中,-20℃保存备用。
1.2.2 烟粉虱总DNA提取
单头烟粉虱成虫经ddH2O清洗后用滤纸吸干,参照罗晨等人的方法提取DNA(罗晨等,2002)。
1.2.3 烟粉虱COI基因的PCR扩增
采用COI的通用引物C1-J-2195(5'-TTG ATT TTT TGG TCA TCC AGA AGT-3')和L2-N-3014(5'-TCC AAT GCA CTA ATC TGC CAT ATT A-3')进行PCR扩增,PCR反应在eppendorf热循环仪中进行。PCR反应体系包含 DNA(20 ng/μL)3 μL,上游和下游引物 (10 μmol/L)各 1.5 μL,10 × Buffer(Mg2+)2.5 μL,dNTP(10 mmol/L)0.5 μL,Taq(Takara)(5 U/μL)0.5 μL,ddH2O 15.5 μL。PCR 条件为95℃预变性5 min;95℃变性50 s,52℃退火50 s,72℃延伸1 min,循环35次;最后72℃再延伸10 min。
PCR产物于1.0%(g/mL)的琼脂糖凝胶上电泳检测 (120 V,0.5×TBE),溴化乙锭 (EB)染色后,在Bio-Rad Gel Doc EQ凝胶成像系统下观察并拍照记录结果。
1.2.4 序列测定
在电泳中检测到目标片段后,用TianGenDNA纯化试剂盒进行回收纯化,然后由上海生工生物工程技术服务有限公司测序,每采集点每种寄主上烟粉虱样品分别测5头。
1.2.5 序列分析
利用BLAST(NCBI网站)与国内外代表地区的烟粉虱COⅠ序列进行比对,通过Clustal X(序列相同时,则选取其中1条)和DNAstar软件对DNA序列进行比对及同源性分析,利用MEGA 5.1软件对序列进行系统发育分析,系统进化树的构建采用最大组成似然 (Maximum Composite Likelihood)模型,建立NJ(Neighbor-Joining)树,各个分支的bootstrap置信度用1000次自导复制检验。
2 结果与分析
2.1 不同地理种群烟粉虱样本的生物型检测及其系统发育分析
在苏南、苏中和苏北不同生态区域,设置了25个采集样点,共125个采集样本,采用mtDNACOI分子标记技术鉴定江苏不同地理种群烟粉虱的Q、B生物型,并在此基础上构建系统进化树。所有样品测序均得到800 bp以上的片段 (图1),分别从每个所测序列中截取720 bp的序列进行分析。通过分子生物学分析软件DNAMAN结合基因库序列比对分析发现,在所检测的125个烟粉虱种群COI基因序列中有118个相同 (序列编号为QJiangsu,简称Q-JS),其余7个COI基因序列一致 (序列编号:B-Jiangsu,简称 B-JS)。与已知序列比较 (表1),以温室白粉虱为外群,建立NJ树 (图2)。由构建的系统进化树可以看到,本试验所检测到的Q-JS与B-JS两个种群的烟粉虱与温室白粉虱的遗传距离较大,首先与温室白粉虱区别开来。Q-JS序列与已知Q型烟粉虱序列Q-Zhejiang、Q-USA和Q-Japan的同源性高达100%,显示Q-JS与已知的Q型烟粉虱种群聚为同一进化枝;B-JS与已知B型烟粉虱序列BPakistan、B-Japan、B-Beijing的同源性为100%,表明B-JS与已知的B型烟粉虱种群聚为同一进化枝。上述序列比对与同源性分析结果表明,江苏不同地理种群烟粉虱Q、B生物型同时存在。
图1 烟粉虱部分样品mtCOI DNA基因片段的PCR产物凝胶电泳Fig.1 PCR products of mtCOI DNA M,分子量参照;1-10,烟粉虱样品;M,Marker;1-10,Bemisia tabaci samples.
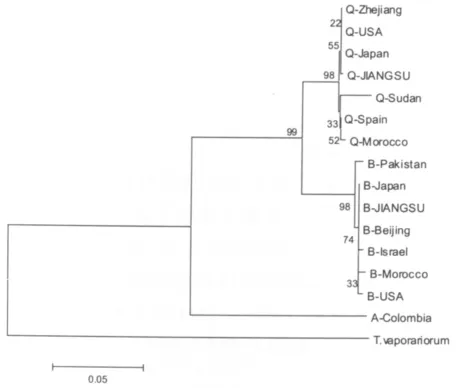
图2 基于COI基因序列构建的系统发育树Fig.2 Phylogenetic tree of B.tabaci populations based on COI sequences

表1 系统进化分析中引用的其他序列信息Table 1 COI sequences cited in this paper
2.2 不同生物型烟粉虱地理分布及主要寄主危害程度
为全面探究江苏不同生态农区烟粉虱的生物型分布与主要寄主受害之间的关系,2010年我们在苏南、苏中和苏北不同生态区域的25个采集样点中涉及了18种主要寄主作物,结合上述生物型检测结果,分析江苏不同地理种群烟粉虱的Q、B生物型分布及在主要寄主作物上的发生危害程度(表2)。序列分析检测结果表明,在所有的采集点与寄主作物上均检测到Q型烟粉虱。Q型烟粉虱的样点检出率为100%,但从样本的角度看,苏南、苏中和苏北的样本数分别为40、25、60,其Q型烟粉虱相应的样本检出率分别是97.50%、96.00%和91.67%。全省的Q型烟粉虱样本检出率为94.40%。从采集寄主上看,Q型烟粉虱在18个寄主上都发生为害,其中在茄科、葫芦科、锦葵科等经济作物上发生与危害程度最重,在十字花科、豆科作物上发生为害较轻,在菊科与苋科作物上发生危害最轻。仅在6个采集点、4种寄主、7个样本上检测到B型烟粉虱,样本检出率为5.6%。从表1可看出,Q型烟粉虱已成为江苏地区的优势生物型,在番茄、黄瓜、茄子、棉花、辣椒等经济作物上虫口密度大,植株受害重,应重视这类寄主作物上Q型烟粉虱的监测与防控。

表2 江苏不同地理种群烟粉虱的生物型分布与主要寄主发生危害程度Table 2 The distribution of the different biotype and occurrence level of B.tabaci in main host crops in Jiangsu
3 结论与讨论
烟粉虱在世界范围广泛分布、寄主植物多样化,生物型和地理分化明显。因此,烟粉虱生物型的监测与鉴定工作既是烟粉虱综合治理的基础,也直接影响到烟粉虱防控策略的优化。已有研究发现本世纪初烟粉虱在我国的传播以B型烟粉虱取代白粉虱和当地烟粉虱为主要趋势 (吴杏霞,2002;臧连生,2005;幐希,2009),此后Q型逐步取代B型,并在我国大部分地区成为优势生物型 (徐婧等,2006;饶琼等,2009)。另外在多个地区也发现非B、Q生物型的存在,如在广西南宁的南瓜寄主上发现非B型烟粉虱 (广西型)(邱宝利等,2003);在山东、河北和浙江发现2个非B/Q型 (褚栋等,2005),其中浙江非B/Q型可能是中国本地种 (臧连生等,2005)。在江苏省,沈媛等从2005~2009年对江苏省烟粉虱生物型进行调查时发现B型和Q型烟粉虱都有分布,同时Q型烟粉虱有从江苏的苏南和苏中向苏北扩散的趋势,2009年Q型烟粉虱在苏北的发生分布频率由2005年的 47.42%上升为 77.78% (沈媛等,2011)。2010年我们采集了江苏省绝大部分地区烟粉虱种群,涉及18种主要寄主,包括棉花、蔬菜和花卉等,其结果可进一步充实不同生物型烟粉虱在江苏地区的遗传分布特性。根据mtDNA COI基因序列分析结果,Q型烟粉虱在江苏省为主要生物型烟粉虱,占所测总样本94.40%。同时试验结果还表明,Q型烟粉虱在苏南、苏中和苏北的比例相差不大,均大于90%,其比例高于沈媛等2009年的检测结果,说明Q型烟粉虱从苏南、苏中扩散到苏北的频率在进一步加快,并有取代B型烟粉虱的趋势。Q型烟粉虱的竞争取代机制是否也存在非对称互作交配现象尚需进一步研究(Liu et al.,2007)。
通过系统聚类树分析发现,江苏地区的Q型烟粉虱与我国浙江、美国和日本的Q型烟粉虱聚为一枝,B型烟粉虱与巴基斯坦、日本和我国北京的B型烟粉虱聚为一枝。从对江苏地区烟粉虱的寄主危害程度的调查发现,Q型烟粉虱在葫芦科、茄科、锦葵科等寄主植物上虫量较高,危害也较重,已经成为危害蔬菜、棉花等安全生产的重要致灾因子。特别是由于设施农业在江苏省大范围推广,有利于Q型烟粉虱危害的寄主、越冬的场所逐年增加,同时Q型烟粉虱对多种新型烟碱类杀虫剂的抗药性高于B型烟粉虱 (Rauch N and Nauen R,2003),因此亟待加强设施农业中Q型烟粉虱的防控。
References)
Bonato O,Lurette A,Vidal C,2007.Modelling temperature-dependent bionomics of Bemisia tabaci(Q biotype).Physiological Entomology,32:50-55.
Chu D,Zhang YJ,Cong B,Xu BY,Wu QJ,Zhu GR,2005.Sequence analysis of mtDNA COI gene and molecular phylogeny of different geographical populations of Bemisia tabaci(Gennadius).Scientia Agricultura Sinica,38(1):76-85.[褚栋,张友军,丛斌,徐宝云,吴青君,徐朱国仁,2005.烟粉虱不同地理种群的mtDNA COI基因序列分析及其系统发育.中国农业科学,38(1):76-85]
Chu D,Zhang YJ,Cong B,Xu BY,Wu QJ,2005.Identification for Yunnan Q -biotype Bemisia tabaci population.Chinese Bulletin of Entomology,(42)1:54-56.[褚栋,张友军,丛斌,徐宝云,吴青君,2005.云南Q型烟粉虱种群的鉴定.昆虫知识,(42)1:54 -56]
Delatte H,Reynaud B,Granier M,2005.A new silverleaf-inducing biotype Ms of Bemisia tabaci(Homoptera:Aleyrodidae)indigenous to the islands of the south- west Indian ocean.Bull.Electrochem.Res.,95(1):29 -35.
Jones D,2003.Plant viruses transmitted by whiteflies.European Journal of Plant Pathology,109:195-219.
Liu SS,De Barro PJ,Xu J,2007.Asymmetric mating:interactions drive widespreadinvasion and displacement in a whitefly.Science,318:1769-1772.
Luo C,Yao Y,Wang RJ,Yan FM,Hu DX,Zhang ZL,2002.The use of mitochondrial cytochrome oxidaseⅠ(mtCOI)gene sequences for the identification of biotypes of Bemisia tabaci(Gennadius)in China.Acta Entomologica Sinica,45(6):759-763.[罗晨,姚远,王戎疆,阎凤鸣,胡敦孝,张芝利,2002.利用mtDNA COI基因序列鉴定我国烟粉虱的生物型.昆虫学报,45(6):759 -763]
Nombela G,Beitia F,Muniz M,2001.A differential interaction study of Bemisia tabaci Q-biotype on commercial tomato varieties with or without the Mi resistance gene,and comparative host responses with the B-biotype.Entomologia Experimentalis et Applicata,3:339-344.
Qiu BL,Ren SX,Wen SY,Nasser S.Mandour,2003.Biotype identification of the populations of Bemisia tabaci(Homoptera:Aleyrodidae)in China using RAPD -PCR.Acta Entomologica Sinica,46(5):605-608.[邱宝利,任顺祥,温硕洋,Nasser S.Mandour,2003.利用RAPD-PCR方法鉴定我国烟粉虱的生物型.昆虫学报,46(5):605-608]
Rao Q,Luo HG,Wang XQ,Zhou GZ,Zhang HY,Luo C,2009.Damage and biotypes identification of Bemisia tabaci(Gennadius)in Wuhan Area.Journal of Huazhong Agricultural University,28(5):535-539.[饶琼,罗汉钢,汪细桥,周国珍,张宏宇,罗晨,2009.武汉地区烟粉虱的危害及其生物型鉴定.华中农业大学学报,28(5):535-539]
Rauch N,Nauen R,2003.Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci(Hemiptera:Aleyrodidae).Arch.Insect Biochem.Physiol.,54(4):165-176.
Shen Y,Du YZ,Ren SX,Qiu BL,2011.Preliminary study of succession of Bemisia tabaci biotypes in Jiangsu Province,China.Chinese Journal of Applied Entomology,48(1):16 -21.[沈媛,杜予州,任顺祥,邱宝利,2011.江苏地区烟粉虱生物型演替研究初报.应用昆虫学报,48(1):16-21]
Teng X,2009.Distribution status of Bemisia tabaci Q-biotype in China and the phylogenetic analysis on mtCOI sequence.Chinese Academy of Agricultural Sciences Master Dissertation.[幐希,2009.Q型烟粉虱在中国的发生现状及基于mtCOI序列的系统发育分析.中国农业科学院硕士学位论文]
Wu XX,2002.The occurrence,distribution and molecular identification of B biotype Bemisia tabaci in China.Chinese Agricultural University Ph.D Dissertation.[吴杏霞,2002.我国B型烟粉虱的发生、分布以及分子鉴定.中国农业大学博士学位论文]
XU Q,Wang WL,Liu SS,2006.The damage of Q biotyoe Bemisia tabaci(Gennadius)in Zhejiang Province.Plant Protection,4:121.[徐婧,王文丽,刘树生,2006.Q型烟粉虱在浙江局部地区大量发生危害.植物保护,4:121]
Zang LS,2005.The competitive replacement and behavioral mechanisms form B biotype betwen non-B biotype Bemisia tabaci in Zhejiang.Zhejiang University Ph.D Dissertation.[臧连生,2005.B型对浙江本地非B型烟粉虱的竞争取代及其行为机制.浙江大学博士学位论文]
Zang LS,Liu SS,Liu YQ,Chen WQ,2005.A comparative study on the morphology and biology between the B biotype and a non-B biotype of Bemisia tabaci from Zhejiang,China.Acta Entomologica Sinica,48(5):742-748.[臧连生,刘树生,刘银泉,陈伟强,2005.浙江B型与一非B型 (China-ZHJ-1)烟粉虱形态物学和生物学特性的比较研究.昆虫学报,48(5):742-748]
