利用trnL-trnF探讨文定果属的系统地位及其与若干近缘科属的系统发育关系*
2012-05-09赖燕玲王晓明陈国培王春波苏应娟廖文波
赖燕玲,王晓明,陈国培,王春波,苏应娟,廖文波
(1.深圳市公园管理中心,广东 深圳 518040;2.深圳市仙湖植物园,广东 深圳 518000;3.中山大学生命科学学院∥广东省植物资源重点实验室,广东 广州 510275)
文定果MuntingiacolaburaLinn.隶属于文定果属,为单型属,主要分布于热带南美洲、西印度群岛,是新热带地区的先锋树种。文定果果实甜香可食,色泽鲜艳,果肉柔软多汁,风味独特,又称南美假樱桃(英文名cherry tree或panama cherry)。此外,该树种还具有较高的观赏价值,其树冠造形美观,枝条伸展成平面状;果实卵状圆球形,浆果状;常全年开花结实,盛花期和果实成熟期常常与授粉昆虫以及传播该植物种子的鸟类活动相一致,被认为是一个重要的园林树种。因其种子常通过食果动物如鸟、蝙蝠、猴子等进行传播,是良好的“诱鸟树”。文定果目前在我国台湾及华南地区有零星栽培,海南地区较常见栽培。
文定果属MuntingiaLinn.自建立以来,首先是放在椴树科,后来得到许多植物学家的认同[1-4]。但也有不同的意见,例如形态特征上也与杜英科、大风子科很相似,或独立成科。Bentham & Hooker[1],Bocquillon[2],Szyszylowicz[3],Hutchinson[4],黄增泉等[5]及吴征镒[6],依据植物体整体特征,如发达的茎皮纤维,以及花结构,认为文定果属应置于椴树科;Shanmukha Rao[7]和 Singh & Dube[8]根据毛状体特征及个体发生过程也赞同放在椴树科。1982年,侯宽昭等[9]将文定果属置于杜英科,而Cronquist[10]、Huber[11]将文定果属置于大风子科。
但文定果属具1枚钻形托叶,腋生花序的特点又区别于杜英科及椴树科。花瓣在芽时具皱纹的特点又与半日花科相似。子房上位但具有环状花盘与椴树科、杜英科及大风子科亦相区别。
Metcalfe & Chalk[12]认为Dicraspidia属和文定果属的解剖结构与杜英科其它属不同,建议成立新科。根据以分子系统学证据为基础的新的APG系统,文定果属确被独立分出成为一个新科,即文定果科Muntingiaceae,仍隶属于锦葵目。Alverson等[13]利用rbcL和18S序列进行分子研究说明,文定果属与杜英科及大风子科亲缘较远。
另外,椴树科的两个单型属NeotessmanniaBurret及DicraspidiaStandl因与文定果属在形态上有诸多相似之处而被长期做为比较对象进行研究,但对于此3属的分类学处理结果却也不同:
1)Hutchinson[4]将文定果属置于Tilieae Bartl.族,将Neotessmannia和Dicraspidia置于Neotessmannieae Barret.族。
2)Takhtajan把Neotessmannia和Dicraspidia由Tiliodeae Arn.亚科中分出,成立新的亚科,即Neotessmannioideae Burret.。
3)Benn & Lemke[14]把这3个属同置于椴树科的Neotessmannieae族中。
Bayer[15]认为以上3属不属于提及的任何一个科,认为在亲缘关系上与这些科亲缘较远,建议成立新科,即文定果科,该科相当于Benn & Lemke[14]的Neotessmannieae 族,包括文定果属MungtingiaLinn.1种,DicraspidiaStandl.1种及NeotessmanniaBurret 1种。在系统关系上,认为文定果属(科)与半日花科,龙脑香科,苞杯花科、沙莓科(或沙莓族)关系较近[16]。
关于杜英科、椴树科和梧桐科科间和属内的系统发育关系的讨论主要是基于形态学、细胞学和孢粉学及部分DNA序列。叶绿体基因组中的trnL-trnF基因间隔区因为属于非编码序列,进化速率快,目前已被广泛用于探讨系统发育关系。本文以杜英科、椴树科和梧桐科植物为材料,通过测定包括外类群在内的25个物种的trnL-trnF序列,并结合GenBank中已公布的3科44个代表种的trnL-trnF序列进行系统发育分析,初步研究了3科科间的系统关系,着重探讨了争议明显的滇桐属、文定果属、杜英属和猴欢喜属的系统位置,以期为它们的系统分类提供分子水平证据。
1 材料和方法
1.1 实验材料
本研究所用材料除文定果外,还包括杜英科、椴树科、梧桐科、大风子科与外类群蔷薇科Rosaceae共69种植物(表1),其中测定了25种植物的trnL-trnF序列,包括文定果属1种、杜英科3种、大风子科3种、椴树科4种、梧桐科11种、蔷薇科4种。另外,从GenBank获取44个物种的trnL-trnF序列,包括杜英科12种、大风子科26种和梧桐科6种。
1.2 实验方法
试剂和仪器:TaqDNA聚合酶、dNTP、100 bp DNA Ladder 购自加拿大Genda Technology 公司;PCR反应在Perkin Elmer Centus 公司2400型热循环仪上进行。
总DNA提取:采用改进的CTAB法。
PCR扩增反应 每100 μL PCR 反应体积中包括:50 mmol/L KCl; 10 mmol/L Tris-HCl; 1.5 mmol/L MgCl2;w=0.1% Triton X-100; dNTP 各 0.2 mmol; 模板用量为50~100 ng; 2.0 单位TaqDNA 聚合酶;引物各40 pmol。扩增引物采用Taberlet等的设计,引物1:5’-ATTTGAACTGGTGACACGAG-3’,引物2:5’-CGAAATCGGTAGACGCTACG-3’。引物由上海博雅生物技术有限公司合成。PCR反应程序为:94 ℃ 30 s,60 ℃ 30 s,72 ℃ 90 s,25个循环;末次循环72 ℃延伸7 min。PCR产物经琼脂糖凝胶电泳检测其相对分子质量。
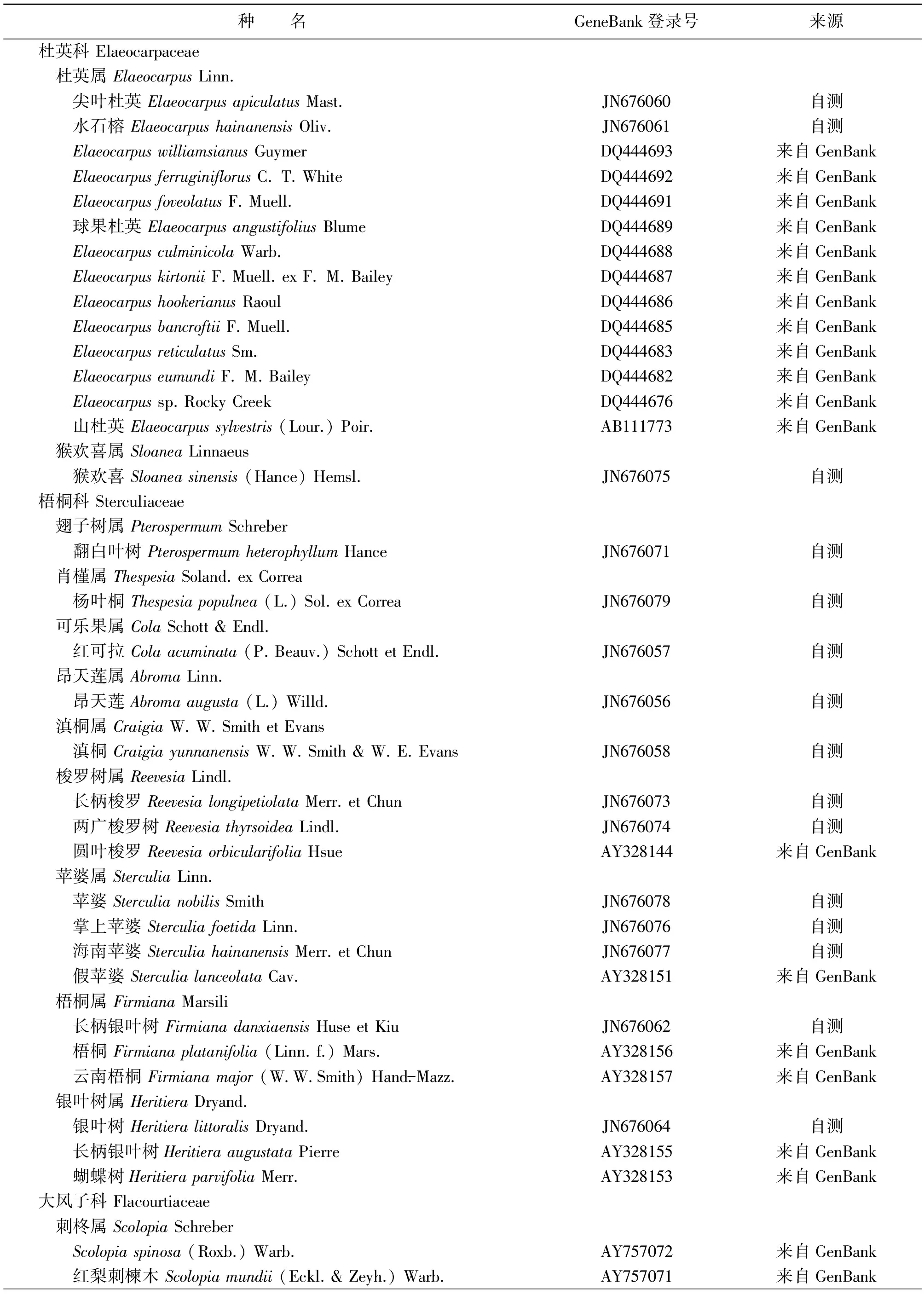
表1 植物材料来源和序列登录号
(续上表)
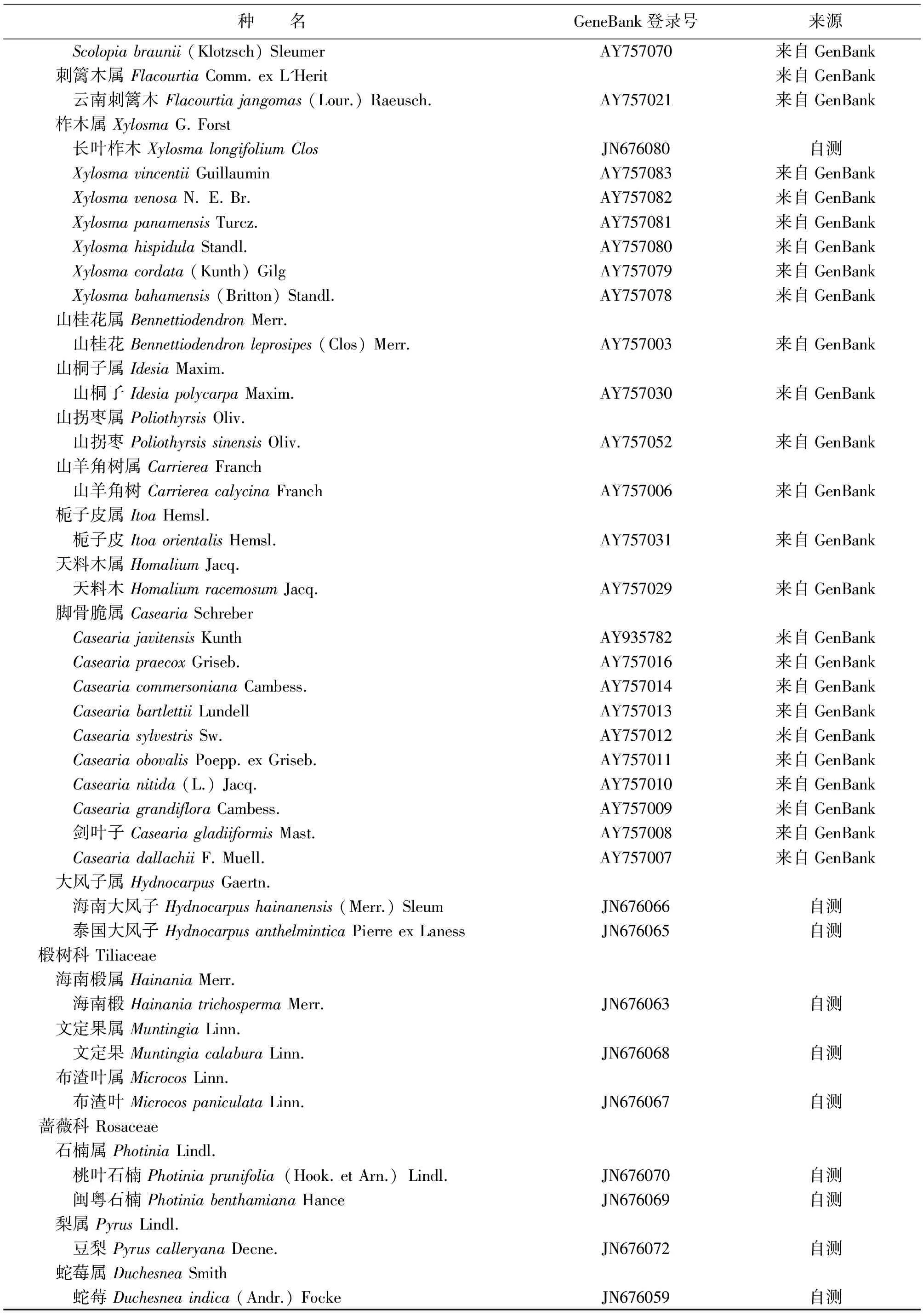
种 名GeneBank登录号来源 Scolopiabraunii(Klotzsch)SleumerAY757070来自GenBank 刺篱木属FlacourtiaComm.exL'Herit来自GenBank 云南刺篱木Flacourtiajangomas(Lour.)Raeusch.AY757021来自GenBank 柞木属XylosmaG.Forst 长叶柞木XylosmalongifoliumClosJN676080自测 XylosmavincentiiGuillauminAY757083来自GenBank XylosmavenosaN.E.Br.AY757082来自GenBank XylosmapanamensisTurcz.AY757081来自GenBank XylosmahispidulaStandl.AY757080来自GenBank Xylosmacordata(Kunth)GilgAY757079来自GenBank Xylosmabahamensis(Britton)Standl.AY757078来自GenBank 山桂花属BennettiodendronMerr. 山桂花Bennettiodendronleprosipes(Clos)Merr.AY757003来自GenBank 山桐子属IdesiaMaxim. 山桐子IdesiapolycarpaMaxim.AY757030来自GenBank 山拐枣属PoliothyrsisOliv. 山拐枣PoliothyrsissinensisOliv.AY757052来自GenBank 山羊角树属CarriereaFranch 山羊角树CarriereacalycinaFranchAY757006来自GenBank 栀子皮属ItoaHemsl. 栀子皮ItoaorientalisHemsl.AY757031来自GenBank 天料木属HomaliumJacq. 天料木HomaliumracemosumJacq.AY757029来自GenBank 脚骨脆属CaseariaSchreber CaseariajavitensisKunthAY935782来自GenBank CaseariapraecoxGriseb.AY757016来自GenBank CaseariacommersonianaCambess.AY757014来自GenBank CaseariabartlettiiLundellAY757013来自GenBank CaseariasylvestrisSw.AY757012来自GenBank CaseariaobovalisPoepp.exGriseb.AY757011来自GenBank Casearianitida(L.)Jacq.AY757010来自GenBank CaseariagrandifloraCambess.AY757009来自GenBank 剑叶子CaseariagladiiformisMast.AY757008来自GenBank CaseariadallachiiF.Muell.AY757007来自GenBank 大风子属HydnocarpusGaertn. 海南大风子Hydnocarpushainanensis(Merr.)SleumJN676066自测 泰国大风子HydnocarpusanthelminticaPierreexLanessJN676065自测椴树科Tiliaceae 海南椴属HainaniaMerr. 海南椴HainaniatrichospermaMerr.JN676063自测 文定果属MuntingiaLinn. 文定果MuntingiacalaburaLinn.JN676068自测 布渣叶属MicrocosLinn. 布渣叶MicrocospaniculataLinn.JN676067自测蔷薇科Rosaceae 石楠属PhotiniaLindl. 桃叶石楠Photiniaprunifolia(Hook.etArn.)Lindl.JN676070自测 闽粤石楠PhotiniabenthamianaHanceJN676069自测 梨属PyrusLindl. 豆梨PyruscalleryanaDecne.JN676072自测 蛇莓属DuchesneaSmith 蛇莓Duchesneaindica(Andr.)FockeJN676059自测
低熔点琼脂糖回收PCR产物:w=1.0 %琼脂糖凝胶1×TAE电泳2 h,在所需条带前填充低熔点琼脂糖,回收PCR产物。
克隆测序:回收所得PCR产物与pUCm-T 载体连接,转化导入由大肠杆菌DH-5α所制的感受态细胞中,采用蓝白斑筛选法获得克隆。应用PCR方法确定阳性克隆。所得阳性克隆在ABI 377 自动测序仪上测序。
1.3 数据分析
数据处理:序列排列用Clustal X 1.83软件自动完成,并进行手工调整。新测序列报送GenBank,登录号见表1。蔷薇科的四种植物桃叶石楠Photiniaprunifolia(Hook.et Arn.)Lindl.、闽粤石楠PhotiniabenthamianaHance、豆梨PyruscalleryanaDecne和蛇莓Duchesneazndica(Andy.) Focke作为外类群。
采用PAUP(Phylogenetic Analysis Using Parsimony)version 4.0进行最简约法(Maximum Parsimony Method)分析;使用PHYLIP(Phylogeny Inference Package) version 3.69 软件包完成最大似然法(Maximum Likelihood Method)分析;用bootstrap值检验树中各分支的置信度,自展数据集为1 000次。运用Treeview软件绘制树图。利用MrBayes(version 3.1.2)完成Bayesian推测(Bayesian Inference,BI),用Modeltest 3.06 软件进行模型选择。似然参数设置:DNA替换采用General Time Reversible (GTR)模型,位点变异速率取γ分布。系统发育模型的后验概率依据Mropolis-Hast ings-Green 算法通过4 条链(Markov Chain Monte Carlo,MCMC)运行15 000 000代估计。利用Tracer v1.4.1软件(Rambaut and Drummond,2009)检验系统发育模型运行程度,当所有参数的有效取样大小(ESS,efficient sampling size)值均大于500时,即可认为运算达到收敛。MCMC 分析以随机树起始,每100代保存1棵树,最初的37 500棵树被当作老化样本摒弃,剩余样本由Figtree v1.2.3 软件(Rambaut and Drummond,2009)[17]构建系统发育树。
2 研究结果
本研究共获得包括外类群在内的69种植物的叶绿体trnL-trnF序列,序列长度变化较大,介于230~429 bp。椴树科的G+C含量为31.14%,序列长度介于383~397 bp,序列最长的是布渣叶(Microcospaniculata),最短的为文定果Muntingiacalabura。梧桐科的G+C含量为31.73%,序列长度范围为369~429 bp,序列最长的是滇桐Craigiayunnanensis,最短的为苹婆Sterculianobilis和假苹婆Sterculialanceolata。杜英科的G+C含量为32.29%,序列长度介于236~306 bp,序列最长的是山杜英Elaeocarpussylvestris,最短的为Elaeocarpussp.'Rocky Creek'。大风子科的G+C含量为32.42%,序列长度介于230-388 bp,序列最长的是泰国大风子Hydnocarpusanthelmintica,最短的为山羊角树Carriereacalycina。
选取蔷薇科的4个物种作为外类群,基于椴树科、梧桐科、杜英科和大风子科26属65个物种的trnLtrn-trnF序列,采用Bayesian推测法构建了系统发育树(图1)。结果表明:
1)文定果属Muntingia独立为一个分支,与椴树科的海南椴属Hainania、布渣叶属Microcos以及梧桐科的类群组成的分支互为姊妹群,分支的后验概率为0.78。而杜英属和猴欢喜属位于杜英科,构成了多歧分支。应予分开。
2)椴树科、梧桐科、杜英科和大风子科均为单系群( monophyletic group)。
3)出现两个互为姊妹群的分支:① 梧桐科和椴树科构成一分支;② 杜英科和大风子科组成另一分支。除椴树科和梧桐科外,杜英科和大风子科分别为单系类群,后验概率分别为1.00和0.59。
4)梧桐科本身包括两个分支,其中一支包括滇桐属Craigia、翅子树属Pterospermum、肖槿属Thespesia、苹婆属Sterculia、可乐果属Cola、梧桐属Firmiana和银叶树属Heritiera,另一支为梭罗树属Reevesia,这两个分支互为姊妹群,后验概率为0.84。
比较最简约法(图2)、最大似然法(图3)和Bayesian推测给出的结果表明,除椴树科、杜英科各属的种间关系外,3种方法构建出的科属内拓扑结构相似,系统发育关系极为吻合,并具有较高的bootstrap支持率。在最简约法和最大似然法分析结果中,椴树科的海南椴属Hainania、布渣叶属Microcos所组成的分支和梧桐科昂天莲属Abroma互为姊妹群。最简约法和最大似然法能区分杜英科杜英属各种之间的系统发育关系。
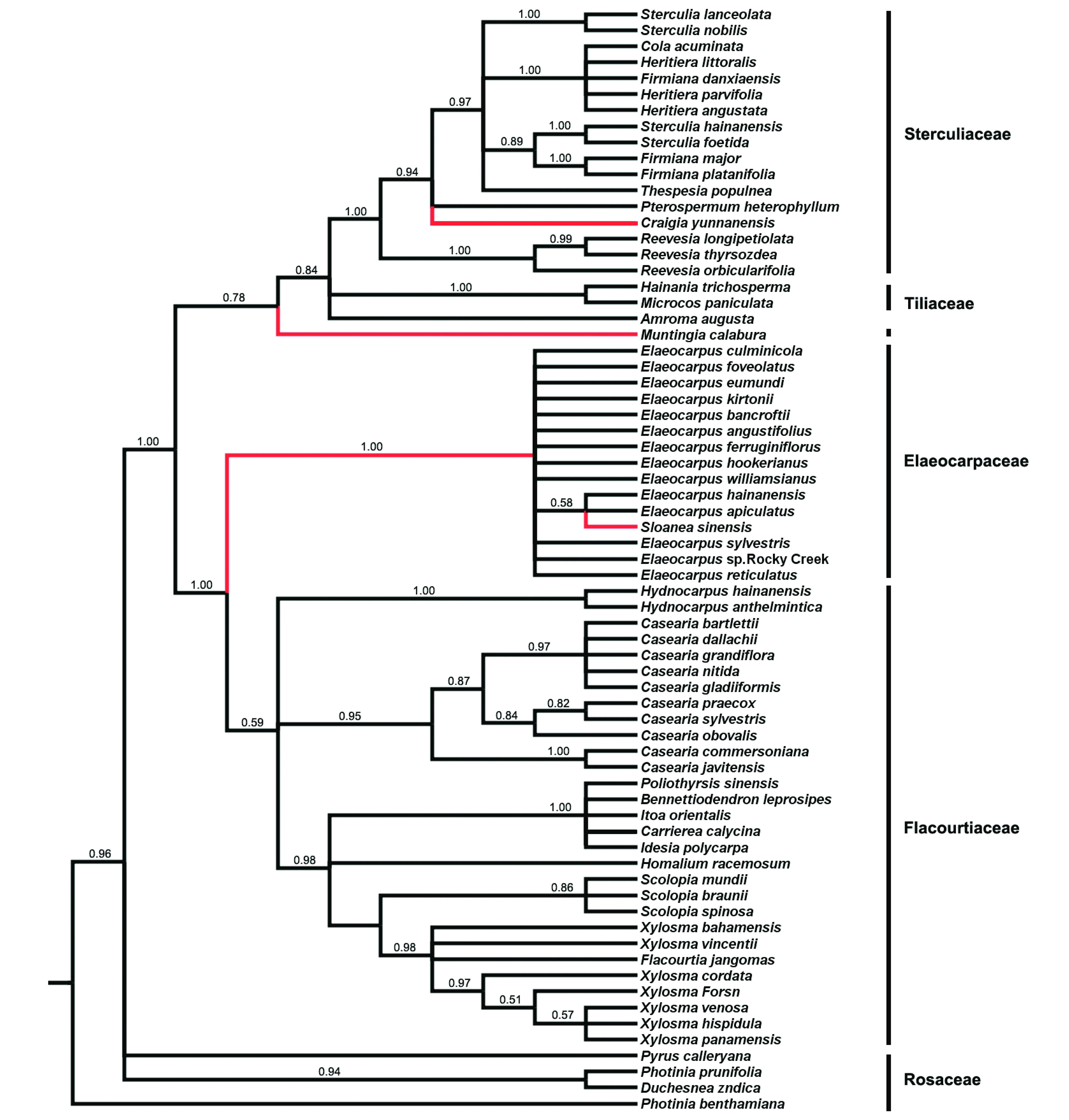
图1 基于trnL- trnF 序列数据利用贝叶斯推测法求得的50 %多数一致树(分支上数字为后验概率值)
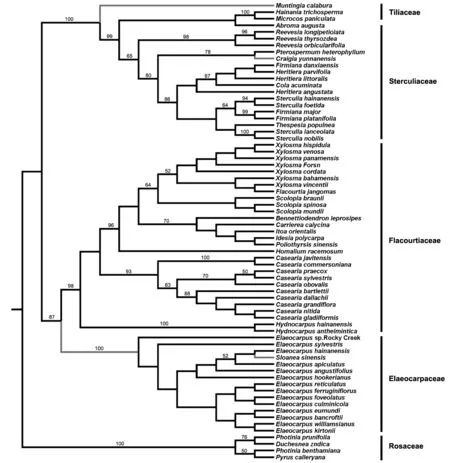
图2 最简约法分析得到的严格一致树
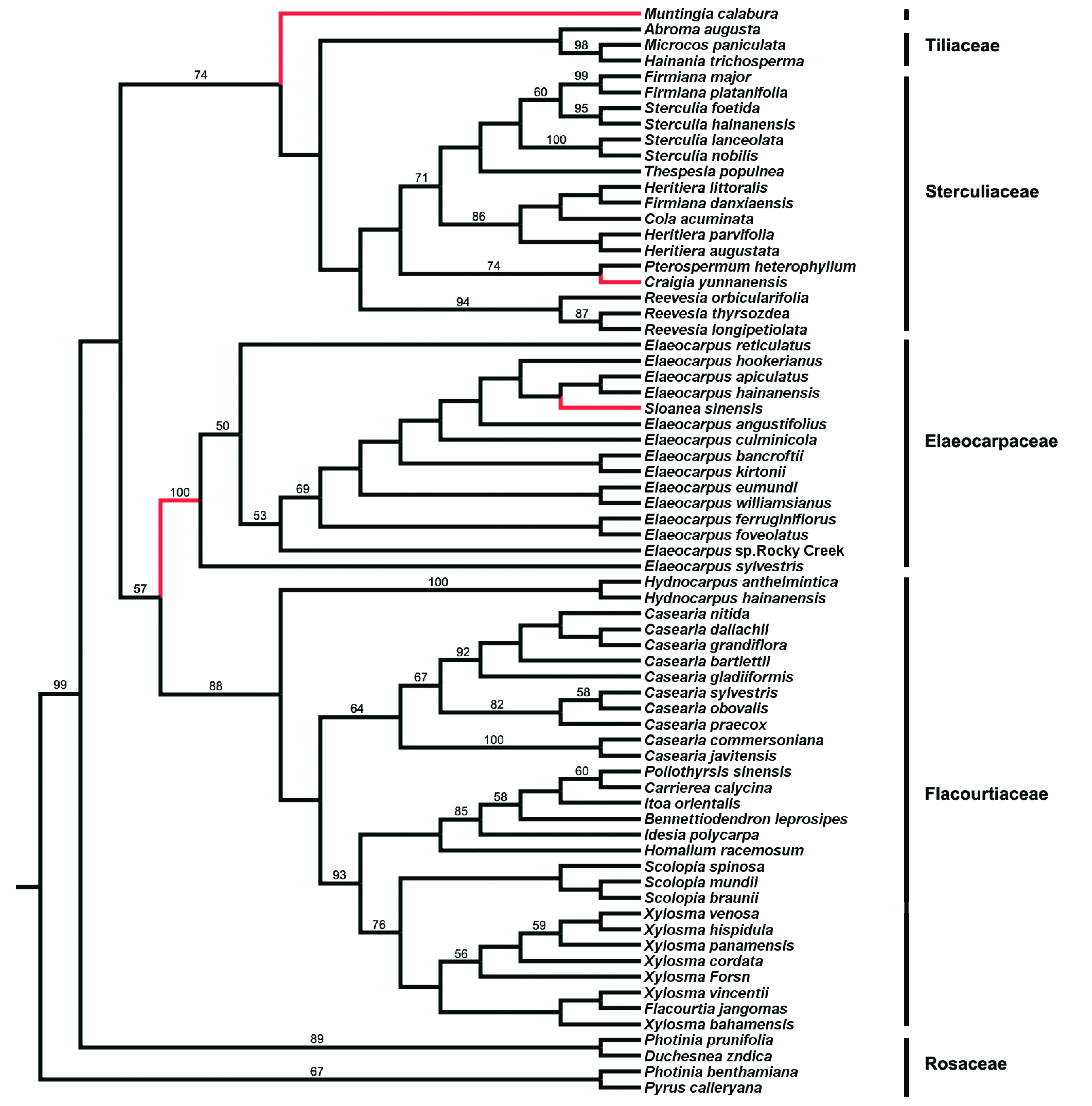
图3 由最大似然法求得的系统发育树
3 讨论与结论
椴树科Tiliaceae、杜英科Elaeocarpaceae和梧桐科Sterculiaceae是锦葵目Malvales的主要成员,一般认为3科关系较密切,但目前也存在争议[13]。椴树科是是被子植物中的一个大科[18],APG分类系统将其合并到锦葵科,成为椴树亚科。杜英科为常绿或半落叶木本,花粉形态学研究支持该科的成立[19]。梧桐科是一个多型科,形态特征极其多样[20]。在梧桐科成立之前,该科成员主要被归于椴树科和锦葵科[21]。梧桐科和椴树科植物形态上很相似,难以找出明显的划分界限,两科曾被合并。椴树科、梧桐科和杜英科之间的复杂关系,导致3科某些属的系统关系也存在明显的争议。
3.1 支持滇桐属Craigia置于梧桐科
滇桐属Craigia为第三纪孑遗属,植物体的生殖器官与花粉形态都比较特殊,长期以来颇受重视,但被认为是“地位不能肯定的属”[22]。该属成立于1921年,是Smith and Evans[23]根据开花阶段不同而建立的,由于其雄蕊形态和梧桐科的毛瓣族Lasiopetaleae和刺果藤族Byttnerieae相似,而置于梧桐科。徐祥浩[24]也赞同放在梧桐科。徐颂军等[25]根据其特殊的花和果形态建议成立滇桐亚科。龙活等[26]认为滇桐属花粉粒的结构和萌发孔的位置不同于椴树科,而与梧桐科相似。根据雄蕊和果实的形态特征,张宏达等[27]将其归于椴树科。分支分析也支持置于椴树科[28]。袁长春等[29],基于核糖体ITS的分析建议滇桐属归于椴树科。新近有几篇文献,也是将滇桐属归于锦葵科椴树亚科[30-32]。
在新版的《Flora of China》中,滇桐属为椴树科中一个独立的属。在本文研究中,trnL-trnF的Bayesian推测表明,滇桐属与梧桐科6个属构成一个单系群,后验概率为0.84。最简约法和最大似然法的结果显示,滇桐与梧桐科翻白叶树Pterospermumheterophyllum互为姊妹群,bootstrap的支持率分别为78%和74%。表明滇桐属与梧桐科的关系非常近,这与基于花果的形态分类和孢粉学研究结果一致。因此,支持将滇桐属归于梧桐科的观点。
3.2 支持杜英属Elaeocarpus和猴欢喜属Sloanea置于杜英科
杜英属Elaeocarpus和猴欢喜属Sloanea也是两个有争议的属。Candolle[33]将猴欢喜属置于椴树科,而不是杜英科。Hutchinson[4]也是将杜英属和猴欢喜属放在椴树科,分别作为两个族。Engler和Cronquist系统将这两个属移至杜英科[10,34]。
杜英属Elaeocarpus和猴欢喜属Sloanea是杜英科的第一和第二大属,分别包括大约200种和120种。猴欢喜属是杜英科惟一一个东亚北美间断分布的属。Hutchinson系统、Engler系统、Cronquist系统对这两个属的处理方式截然不同,分歧点是置于椴树科还是杜英科。
本文研究结果,虽然trnL-trnF序列无法分辨属内种间的关系,但最简约法、最大似然法和Bayesian推测的结果显示,这两个属是杜英科的主要成员,应归于杜英科,这与Engler系统和Cronquist系统相一致。目前《Flora of China》也采用Engler系统和Cronquist系统处理方式,将杜英属和猴欢喜属归于杜英科。
3.3 支持文定果属Muntingia独立为文定果科
文定果属Muntingia是另一个分类地位存在较大争议的属。Burret[35]、Edlin[36]、Hutchinson[4]、Cronquist[10]、Takhtajan[37-38]以及Benn和Lemke[14]把文定果属放到椴树科。Engler系统将其置于杜英科[34]。Metcalfe and Chalk[12]、Robyns[39]、Smith[40]、Brizicky[41]、Gasson[42]认为文定果叶具粘液腔、花萼镊合状、子房多室且花粉粒大小与杜英科的一些属较相似,应放入杜英科更合适。
由于杜英科、椴树科以及大风子科部分物种的关系密切,文定果属甚至被建议置于大风子科[10-11]。
最近,Bayer等[15]根据形态学特征和分子数据成立了文定果科Muntingiaceae,隶属于锦葵目,包含文定果属Muntingia、Neotessmannia和Discraspidia,其毛被特征、叶形态、花位置、镊合状的花萼、花瓣形状、下位的子房均不同于锦葵目中其它科。这个结论也得到了纹孔研究结果的支持[43]。
本研究的最简约法、最大似然法和Bayesian推测的结果发现,文定果构成一个独立于梧桐科和椴树科的分支,bootstrap 的支持率和后验概率分别为100%、74%和0.78;也表明其与梧桐科和椴树科的系统发育关系密切,支持其独立成科的观点。
3.4 文定果科、椴树科、梧桐科和杜英科的系统关系
Cronquist系统、Takhtajan系统和Thorne系统等一致认为椴树科、梧桐科和杜英科的关系密切,属于锦葵目(Malvales)。但是在基于rbcL、atpB和18S rDNA建立的APG分类系统中,椴树科和梧桐科被合并到锦葵科中,成立了广义锦葵科,置于锦葵目中。本研究由于缺乏锦葵科的样品,无法探讨椴树科、梧桐科和锦葵科的关系,但trnL-trnF序列分析证实椴树科、梧桐科和杜英科的有较近的亲缘关系。从最简约法、最大似然法和Bayesian推测结果可以看出,椴树科和梧桐科的关系非常密切,两科并没有构成两个独立的单系群,而是形成复系群。椴树科和梧桐科这种复杂的关系也存在于ITS序列的研究中[29]。另外,基于trnL-trnF结果,明确支持杜英科为一个独立的科,这与孢粉学的研究结果一致。
从毛状体结构及个体发生来看,文定果科与椴树科相似[8],但其具星状毛、球状头型的腺状毛状体结构在椴树科、杜英科、大风子内未发现。特别是粘液腔或粘液道、叶、托叶、毛被特征、花序形态、花蜜腺、下位子房具有沟状下延的柱头、胚珠数目、倒生的胎座、花粉粒小、胚发育、纤维状的外珠被等方面均不同于椴树科[12,41,44-45]。
甚至,Cronquist[10]认为文定果花药短而宽、纵裂,不同于杜英科,而更像大风子科的某些属,因此将其放入大风子科。但William等[46]认为分子系统学数据表明文定果属与杜英科、大风子科、椴树科等亲缘关系较远,而与龙脑香类等类群亲缘关系较近。因此,文定果科的系统地位还有待于更多的深入研究,特别应与包括椴树科、梧桐科、杜英科、大风子科、龙脑香科、锦葵科在内若干属进行系统发育比较研究。
参考文献:
[1]BENTHAM G,HOOKER J D.Genera plantarum[M].London:[s.n.],1862,1(1):236.
[2]BOCQUILLON H.Memoire sur le groupe des Tiliacees[J].Adansonia,1866,7: 17-64.
[3]SZYSZYLOWICZ I .Zur Systematik der Tiliaceen[J].Bot Jahrb Syst,1885,6 : 427-457.
[4]HUTCHINSON J.The genera of flowering plants (Angiospermae) (vol.2 )[M].Oxford: Clarendon Press,1967.
[5]黄增泉.植物分类学-台湾维管束植物科志[M].台北: 南天书局,1993.
[6]吴征镒,孙航,周浙昆,等.中国植物区系中的特有性及其起源和分化[J].云南植物研究,2005,27(6): 577-604.
[7]SHANMUKHA RAO S R.Trichome ontogenesis in some Tiliaceae[J].Beitr Biol Pflanzen,1990,65: 363-375.
[8]SINGH H B,DUBE V P.Taxonomic significance of foliar epidermal features ofMuntingiaLinn.(Tiliaceae)[J].J Pl Anat Morphol,1993,6: 123-128.
[9]侯宽昭.中国种子植物科属词典[M].北京:科学出版社,1982.
[10]CRONQUIST A.An integrated system of classification of flowering plants[M].New York: Columbia University Press,1981.
[11]HUBER H.Angiospermen: Leitfaden durch die Ordnungen und Familien der Bedektsamer[M].New York: Stuttgart.1991.
[12]METCALFE C R,CHALK L.Anatomy of the dicotyledons.vol.2[M].Oxford: Clarendon Press,1950.
[13]ALVERSON W S,WHITLOCK B A,NYFFELER R,et al.Phylogeny of the core Malvales: evidence fromndhF sequence data[J].American Journal of Botany,1999,86(10): 1474-1486.
[14]BENN S J,LEMKE D E.Taxonomy of Neotessmannieae (Tiliaceae)[J].Amer J Bot,1991,78 (suppl):166-167.
[15]BAYER C,CHASE W M,FAY F M.Muntingiaceae,a new family of Dicotyledons with malvalean affinities[J].Taxon,1998,47(1): 37-42.
[16]张宏达.种子植物系统学[M].北京:科学出版社,2004.
[17]RAMBAUT A,DRUMMOND A J.FigTree v1.2.3[EB].http: // tree.bio.ed.ac.uk/ software/ figtree/[2009-10-23].
[18]PHILLIPS O,MILLER J S.Global patterns of plant diversity: Alwyn H.Gentry’s forest transect data set[M.USA: Missouri Botanical Garden,St.Louis,2002.
[19]唐亚,吴征镒.国产杜英科花粉形态的研究[J].云南植物研究,1990,12(4): 397-403.
[20]WHITLOCK B A,BAYER C,BAUM D A.Phylogenetic relationships and floral evolution of the Byttnerioideae ("Sterculiaceae" or Malvaceae s.l.)based on sequences of the chloroplast gene,ndhF[J].Systematic Botany,2001,26(2): 420-437.
[21]BAYER C,FAY M F,de BRUIJN A Y,et al.Support for an expanded family concept of Malvaceae within a recircumscribed order Malvales: a combined analysis of plastidatpBandrbcLDNA sequences[J].Botanical Joumal of the Linnean society,1999,129: 267-303.
[22]ERDTMAN G.Pollen Morphology and Plant Taxonomy,Angiosperms[M].USA: The Chronica Botanica Co,1952.
[23]SMITH M A,EVANS W E.Craigia,a new genus of Sterculiaceae[J].Trans Bot Soc Edinburgh,1921,28: 69-71.
[24]徐祥浩.中国梧桐科植物的整理[J].植物分类学报,1977,15 ( 1) : 73- 84.
[25]徐颂军,徐祥浩.梧桐科一些属的分类位置探讨[J].热带亚热带植物学报,2000,8 (1):11-16.
[26]龙活,何丽卿,徐祥浩.中国梧桐科植物花粉类型的研究[J].华南农业大学学报,1989,10(1):23-32.
[27]张宏达,缪汝槐.椴树科岘木亚科的系统分类[J].中山大学学报:自然科学版,1978,17(3) :19- 26.
[28]诸葛仁.滇桐属系统位置的分支分析[J].云南植物研究,1989,11 (1) : 17- 23.
[29]袁长春,施苏华,钟扬,等.用核糖体DNA的ITS序列探讨滇桐属的系统学位置[J].中山大学学报:自然科学版,2002,41(6): 73-77.
[33]CANDOLLE A P.De Prodromus Systematis Naturalis Regni Vegetabilis (Vol.3 )[M].[s.n.],1824: 503-520.
[34]ENGLER A.,PRANTL K.Die Naturlichen Pflanzenfamilien[M].Leipzig : Verlag von Wilhelm Englmann,1895:1-8.
[35]BURRET M.Beiträge zur Kenntnis der Tiliaceen[J].Notizbl Bot Gart Berlin- Dahlem,1926,9: 592-797.
[36]EDLIN H L.A critical revision of certain taxonomic groups of the Malvales[J].New Phytologist,1935,34: 1-20,122-143.
[37]TAKHTAJAN A.Systema Magnoliophytorum[M].Editoria Nauka,Leningrad (St.Petersburg),1987.
[38]TAKHTAJAN A.Diversity and Classification of Flowering Plants[M].New York:Columbia University Press,1997.
[39]ROBYNS A.Family 114.Tiliaceae[J].Ann Missouri Bot Gard,1964,51: 1-35.
[40]SMITH C E.Family 113.Elaeocarpaceae.[J].Ann Missouri Bot Gard,1965,52: 487-495.
[41]BRIZICKY G K.The genera of Tiliaceae and Elaeocarpaceae in the southeastern United States[J].J Arnold Arbor,1965,46: 286-307.
[42]GASSON P.Wood anatomy of the Elaeocarpaceae[M]// DONALDSON L,et al,eds.Recent Advances in Wood Anatomy.Rotorua,1996,47-71.
[43]JANSEN S,BAAS P,SMETS E.Vestured pits: their occurrence and systematic importance in eudicots[J].Taxon,2001,50:135-167.
[44]CORNER E J H.The Seeds of Dicotyledons[M].Cambridge: Cambridge University Press,1976.
[45]BAYER C,BAYER C.Zur infloreszenz morphologie der Malvales[J].Dissertationes Botanicae,1994,212:1-280.
[46]WILLIAM G V M,TALLON J L,QUILTY J W,et al.Absence of an isotope effect in the pseudogap in yba2Cu4O8as determined by high-resolution89YNMR[J].Phys Rev Lett,1998,80,377-380.
