特早熟陆地棉的遗传效应及杂种优势分析
2012-04-17詹有俊孙建船庄生仁任福成
詹有俊,杨 涛,孙建船,庄生仁,任福成,张 璇,李 渊
国内外研究表明,棉花杂种优势利用是提高产量,改进纤维品质的有效途径之一。杂种棉具有较强的营养生长和生殖生长优势,表现在棉株生长势强,营养体生长快,植株高大,枝叶繁茂,铃多铃大,衣分提高,熟性提前等方面[1]。在甘肃河西走廊特早熟棉区开展杂交棉选育应用,是解决近年来棉花产量徘徊不前的技术关键。该棉区热量条件十分有限,≥10℃的有效积温3000℃-3600℃,无霜期140d-150d,春季气温回升慢,秋季气温下降快,早霜来的早,且强度大。棉花有效生长发育时期比较集中,植棉区土壤肥沃,灌溉便利,栽培管理精细,单产高,品质优。长期以来,形成了宽幅地膜覆盖、高密度种植、矮化促早的栽培模式。与其他棉区相比,生产上对品种的产量、早熟性要求较高。棉花杂种优势研究利用一直处于空白状态,加之用作研究分析的材料和试验环境条件不同,试验结果难以直接在该区应用。基于此种情况,我们对五个特早熟棉花品种进行完全双列杂交,分析其亲本及其F1代的农艺经济性状遗传表现。为因地制宜地开展杂种优势利用和杂交棉品种选育,进一步提高产量,增进纤维品质和增加植棉整体效益提供理论依据。
1 材料与方法
1.1 亲本材料
本试验选用五个特早熟陆地棉品种。依次为P1:酒棉1号,大铃品种;P2:酒棉8号,早熟性特好;P3:361,结铃性强;P4:酒棉17号,高衣分;P5:新陆早24号,纤维品质优良。
1.2 试验方法和性状调查
2010年用5个品种进行完全双列杂交。2011年在敦煌进行田间试验,采用随机区组设计,亲本、F1代各种植2行,行长9 m,重复三次。用1.45 m幅宽的地膜覆盖,一膜种植4行棉花,行距0.4 m,株距0.15 m,设计密度每公顷16.5万株。试验管理同当地大田。
棉花生长期间,每个小区连续选取有代表性的20株作为观察记载对象,调查主要农艺性状。收获前每小区选拾20个棉铃进行考种,测定铃重、衣分等。并按小区收获计产。各小区皮棉样送农业部棉花纤维检验监督测试中心进行纤维品质测定。
1.3 数据分析
采用朱军[2]提出的遗传模型分析方法和QGA Station软件进行分析。用AUP法预测遗传效应值,用Jackknife数值抽样技术,计算各项遗传参数的估测值,并用t测验对遗传参数进行显著性检验。用GRIFIFING法分析配合力和遗传参数估计值。
2 结果与分析
2.1 特早熟陆地棉亲本及其F1的主要农艺经济性状分析
表1分析结果表明,5个亲本品种所配制的F1的平均子棉产量均高于其亲本品种,增产幅度为5.93%-49.07%,平均增产25.02%;皮棉产量的变化趋势与子棉相同,其F1的增产幅度为6.43%-66.67%,平均增产30.71%,子、皮棉产量均表现出较强的杂种优势。亲本品种中,以酒棉17号所配制的杂种F1的平均子、皮棉产量均表现最高,分别比其增产32.63%、35.97%;以亲本361所配制的杂种F1的平均子、皮棉产量均居第二位,分别比其增产5.93%、6.43%;其余三个亲本所配制的杂种F1的平均子、皮棉产量虽比其增幅较大,但其亲本产量相对较低,F1的绝对产量也不高。说明选取对杂种一代贡献大的亲本品种是选配杂交组合的关键所在。

表1 五个品种完全双列杂交的F1代主要农艺性状平均值与其亲本的增减比率 (单位:%)
从产量构成的直接因子单株结铃数、单铃重和衣分来看。5个品种所配制的F1平均单株结铃数的杂种优势强,平均为16.47%;单铃重和衣分的杂种优势较弱,分别为4.30%和4.10%。由此说明,单株结铃数增加是杂种产量增加的主要因子。5个亲本所配制的F1的平均单株结铃数,除结铃性强的亲本361外,其余亲本所配制的F1均明显增多,增幅为5.73%-39.86%。在平均单铃重上,除大铃亲本酒棉1号外,其余亲本所配制的F1均表现增重,增幅为2.52%-9.52%。在平均衣分上,各个亲本所配制的F1均表现出不同程度的杂种优势,增幅为0.59%-11.58%。
与早熟有关的性状中,平均生育期、铃期除特早熟亲本酒棉8号外,其余4个亲本所配制的F1代的平均值均比其亲本品种有所缩短,生育期缩短4d-6d,铃期缩短3d-6d,4个亲体平均杂种优势分别为3.71%和5.59%。平均霜前花率除特早熟亲本酒棉8号外,其余4个亲本所配制的F1代平均值均比其亲本品种高,平均杂种优势为7.96%。平均始蕾期、始花期除前期发育快的酒棉1号外,其余4个亲本品种所配制的F1均比其亲本有所缩短,平均杂种优势分别为7.03%和2.04%。以上分析说明,杂种F1代普遍早熟于其亲本,体现在生育期、铃期、始蕾期、始花期普遍缩短,霜前花率普遍增加。但杂种F1代果枝始节和果枝高度明显增加,这与棉株生长势增强有关。
5个品种所配制的F1代平均纤维品质与其亲本相比,杂种优势不甚明显。F1代的平均整齐度指数和反射率值比其亲本略有增加。其余六个品质性状F1代与其亲本相比有增有减。马克隆值除亲本酒棉1号外,其余4个亲本所配制的F1代的平均值都增加。纤维上半部平均长度、断裂比强度、纺纱均匀指数等品质指标表现趋势一致,优质亲本新陆早24号和361,其F1代的杂种优势值均为负值,相差相对较大;其余亲本品质性状较为接近,其F1代平均值与之相差不大。
2.2 特早熟陆地棉主要农艺经济性状的配合力和遗传效应分析
配合力的高低是选择杂交亲本的重要依据。研究分析表明(表2):所有产量性状的一般配合力和特殊配合力均达极显著水平,正反交组合只有单铃重、百粒重、子指和衣指达显著水平,其余均不显著,说明子、皮棉产量、结铃数、衣分等多数产量性状是受核基因控制的。早熟性状的一般配合力和特殊配合力均达极显著水平。正反交效应除始蕾期外,其余性状达(极)显著水平。说明早熟性不完全受核基因作用,还与选用杂交的父母本次序有关。纤维五项品质指标一般配合力和特殊配合力均达(极)显著水平,正反交效应均不显著,说明这五项品质性状也是受核基因控制的。
遗传分析表明,产量构成因素中,除单铃重外,子、皮棉产量、结铃数、衣分的显性效应较强,表现出较强的杂种优势。反映早熟的生育期、霜前花率、始花期、铃期等性状,以加性效应为主,显性效应也相当显著,杂种F1代熟性提前。纤维上半部平均长度、断裂比强度、纺纱均匀指数等主要品质性状以加性效应为主,显性效应较弱,杂种优势不太明显。
对亲本品种的一般配合力进一步分析得出(表3),产量性状中,酒棉17号和361的子、皮棉产量、结铃数、衣分的配合力均表现较高,但单铃重配合力较低。品质性状中,新陆早24号和361的上半部平均长度、断裂比强度、纺纱均匀指数的配合力均较高,马克隆值和伸长率的配合力较低。早熟性状中,酒棉8号和酒棉1号的霜前花率、生育期、果枝始节、始蕾期、始花期的配合力均较高。
2.3 特早熟陆地棉主要农艺经济性状的杂种优势分析
遗传分析表明(表4),产量直接构成因素中,F1代的基因型值均达(极)显著水平,群体平均优势预测值均为正值,且

表2 五个品种完全双列杂交的F1代农艺经济性状配合力的方差分析及遗传参数估计值
注:+、*和**分别表示达到0.10、0.05和0.01显著水平,下同。达极显著水平,群体超亲优势预测值除铃重为负值外,其余四个性状均具有正向超亲优势,皮棉产量达到了0.10的显著水平。与早熟有关的性状,F1代的群体平均优势和超亲优势趋势一致,始蕾期、始花期、铃期和生育期的优势预测值均为负值,霜前花率为正值。说明蕾、花、铃发育天数缩短,霜前花率增加,以上各性状均表现早熟。纤维品质性状中,F1代的基因型、群体平均优势和超亲优势预测值多数未达显著水平,多数性状具有微弱的群体平均优势,没有群体超亲优势。

表3 五个亲本品种农艺经济性状的一般配合力效应值
对10个正反交组合的遗传分析表明(表5),子棉产量、皮棉产量、结铃数、单铃重和衣分等产量性状,群体平均优势的优势组合数均较多,优势组合率占80%-100%。群体超亲优势的优势组合率占50%-100%,以子棉产量和衣分的优势组合数居多,优势组合率分别为90%和100%。纤维品质方面(表6),上半部平均长度、断裂比强度和马克隆值的群体平均优势的优势组合率分别为70%、40%和80%。群体超亲优势除四个组合的马克隆值具有正向优势外,其余组合均没有超亲优势。早熟性状的各个杂交组合均趋向于早熟,表现在生育期没有群体平均优势和超亲优势,霜前花率均具有群体平均优势和超亲优势。

表4 五个品种完全双列杂交的平均表现型值和基因型及杂种优势预测值
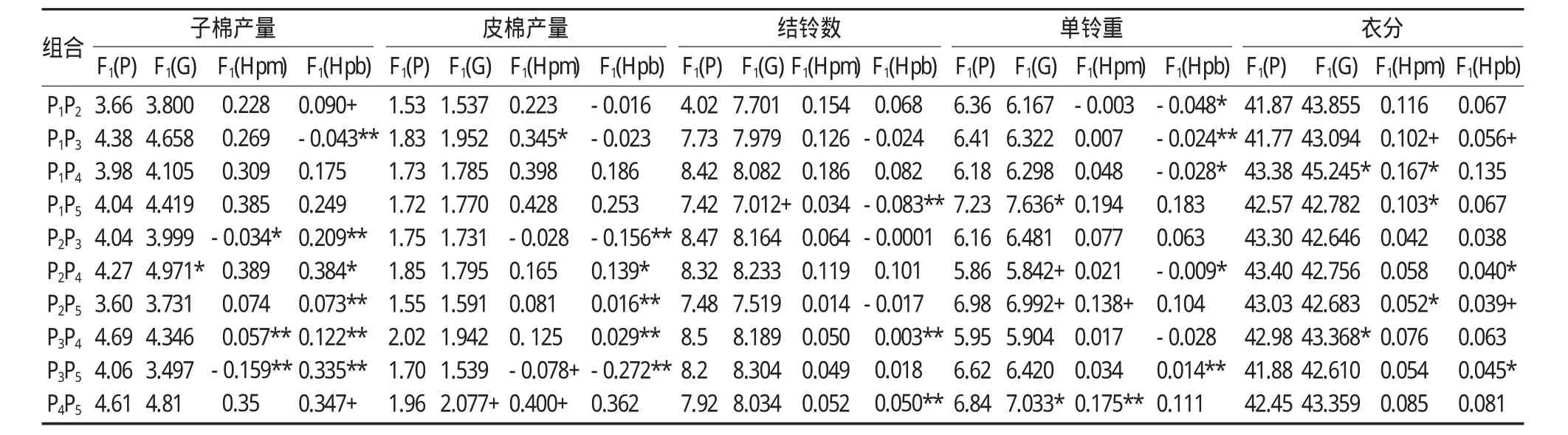
表5 五个品种完全双列杂交组合产量性状的表现型值和基因型及杂种优势预测值

表6 五个品种完全双列杂交组合早熟性、品质性状的表现型值和基因型及杂种优势预测值
3 结论与讨论
3.1 结论
双列杂交研究表明,特早熟陆地棉的产量、早熟、纤维品质性状的一般配合力和特殊配合力方差均达(极)显著水平。遗传效应中,产量性状以显性效应为主,早熟性和品质性状以加性效应为主。杂种优势分析得出,产量及其构成因素优势明显,产量各主要性状具有极显著的群体平均优势,群体超亲优势除单铃重外,均有优势。早熟性状杂种优势比较明显,各性状趋势一致,F1代均表现早熟,蕾、花、铃发育天数缩短,霜前花率增加。纤维品质杂种优势不明显。组合P3P4综合表现好,子棉、皮棉产量均居于10个正反组合的首位,且子棉产量的群体平均优势和超亲优势均达极显著水平,皮棉产量的群体超亲优势达极显著水平,熟性适中,品质优良。亲本酒棉17号的一般配合力好,其所配制的高优势产量组合多,新陆早24号次之。
3.2 讨论
陆地棉品种间杂交种的农艺经济性状表现主要取决于双亲的一般配合力,亲本品种性状表现与其一般配合力呈正相关关系。朱军等[3]、张正圣等[4]研究得出,当性状的一般配合力方差大于特殊配合力方差时,可以只根据亲本的一般配合力表现预测F1的性状表现。本研究所选用的特早熟陆地棉品种间杂交,其产量、早熟性和纤维品质性状的一般配合力均达到极显著水平,且大于相应的特殊配合力值。由此可以根据亲本配合力表现筛选合适的杂交亲本,进而筛选出强优势的杂交组合和积极有效地进行棉花杂交育种。
陆地棉产量、早熟性和纤维品质等主要性状均是由多基因控制的数量性状,同时还受其他因素影响[5]。宋美珍等[6]研究表明,短季棉产量及其直接构成因素均以显性效应遗传为主,其次是加性效应。生育期、铃期、始花期等早熟性状以加性效应为主,其次为显性效应。本研究表明,子棉产量、皮棉产量、结铃数和衣分等产量性状具有极显著的显性和加性效应,且显性效应值大于加性效应值,这与上述研究结果相一致。单铃重以加性效应为主,显性次之,研究结果与宋美珍等不相一致,与吴振衡等[7]结果一致。早熟性状受环境因素影响较小,主要受基因的遗传效应作用,生育期、霜前花率、始花期、铃期等以加性效应为主,显性次之,这与宋美珍等[6]的研究相同。纤维品质性状受环境因素影响较小,基因的遗传效应起着主要作用,且以加性效应为主,这与前人[5-10]研究相同。
作物杂种优势利用的终极目标是根据杂交组合的基因型和环境表现,筛选出适合当地特定生产条件的高产高效、强优势杂交组合。邢朝柱等[11]研究转基因抗虫杂交棉在不同环境条件下的性状表现,得出陆地棉品质性状间杂种优势不明显,受环境因素影响较小。产量杂种优势明显,产量各性状均表现极显著水平正向优势,以皮棉产量优势最大,子棉产量其次。本研究的结果表明,特早熟陆地棉纤维品质的杂种优势不甚明显,环境因素对其影响较小,主要决定于用作杂交的亲本的纤维品质性状表现。子棉产量、皮棉产量杂种优势表现与上述结论一致。产量构成因素中,单株结铃数优势最强,单铃重次之,衣分的优势率最低。而且环境条件对产量及其构成因素各性状具有极显著水平的影响效应,这与该棉区采用高密度、矮化促早栽培措施有关,棉株个体生长发育空间有限,棉株杂种优势效应不能得以充分发挥。
致谢:此研究得到浙江大学朱军教授的指教和大力帮助,深表谢意!
[1] 张天真,靖深蓉,金林,等.杂交棉选育的理论与实践[M].北京.科学出版社,1998.
[2] 朱军.遗传模型分析方法[M].中国农业出版社,1997.
[3] 朱军,季道藩.陆地棉产量性状的双列分析[J].浙江农大学报,1987,13(3):280-287.
[4] 张正圣,李先碧,刘大军,等.陆地棉高品质系的杂种优势利用研究[J].棉花学报,2002,14(5):264-268.
[5] 潘家驹.棉花育种学[M].北京.中国农业出版社,1998:273-295.
[6] 宋美珍,喻树迅,范术丽,等.短季棉主要农艺性状的遗传分析[J].棉花学报,2005,17(2):94-98.
[7] 吴振衡,刘定俊,莫惠栋,等.陆地棉数量性状的遗传分析[J].遗传学报,1985,12(5):334-349.
[8] 肖光顺,李保成,谢宗铭,等.早熟陆地棉主要纤维品质性状的遗传效应及杂种优势研究[J].作物杂志,2010(6):62-66.
[9] 袁有禄,张天真,郭旺珍,等.棉花高品质纤维性状的主基因与多基因遗传分析[J].遗传学报,2002,19(9):827-734.
[10] 袁有禄,张天真,郭旺珍.棉花纤维品质性状的遗传稳定性分析[J].棉花学报,2002,14(2):67-70.
[11] 邢朝柱,喻树迅,郭立平,等.不同生态环境下陆地棉转基因抗虫杂交棉遗传效应及杂种优势分析[J].中国农业科学,2007,40(5):1056-1063.
