黄独 (D ioscorea bu lbifera L.)不同居群叶表皮微形态特征的比较观察
2011-12-31周义峰杭悦宇
彭 斌,周义峰,舒 璞,杭悦宇
〔江苏省·中国科学院植物研究所(南京中山植物园)江苏省植物迁地保护重点实验室,江苏 南京 210014〕
黄独 (DioscoreabulbiferaL.)隶属于薯蓣科(D ioscoreaceae)薯蓣属 (DioscoreaL.)基生翅组 (Sect.OpsophytonU line),广布于东南亚、大洋洲、非洲和美洲[1],属于古热带种 (白垩纪至第三纪),是基生翅组的祖先型[2]。基生翅组全世界共仅有 5~6种,广布于亚洲、非洲及南美洲。Prain等[3]认为:分布在中国的基生翅组包含有 1种 3变种,即黄独及其变种雷公薯 (D.bulbiferavar.deltoideaPrain etBurkill)(分布于中国广东)、大花黄独 (D.bulbiferavar.simbhaPrain et Burkill)(分布于中国云南腾冲 )和D. bulbiferavar.veraPrain etBurkill(分布于中国四川茂汶)。然而,《Flora of China》[4]一书正式收载的基生翅组仅包含黄独 1种,分布于中国黄河以南地区,仅将 Prain等提出的 3个变种作为文献提及,并没有认可这些变种。
近年来,国内外学者针对黄独种下类群的系统学进行了大量研究。Zhou等[5]将分布于中国云南的根茎断面白色的黄独变种定名为白金山药 (D.bulbiferavar.albotuberosaY.F.Zhou,Z.L.Xu et Y.Y. Hang),并认为在中国新分布的黄独变种有雷公薯和大花黄独等;另外,周义峰[6]认为有少花黄独 (D. bulbiferavar.pauciflorumY.F.Zhou et Y.Y.Hang)和枝花黄独 (D.bulbiferavar.ramiflorusY.F.Zhou et Y.Y.Hang)等新拟变种存在 (未正式发表)。Terauchi等[7-8]以分布于非洲的黄独变种D. bulbiferavar.anthropo-phagorun(A.Chev.)Summ erh为材料,绘制了黄独叶绿体基因组图谱,并采用RFLP分子标记对来自非洲、亚洲和大洋洲的 6个黄独及其变种共 15个样品的亲缘关系进行了研究。郑玉红等[9]则利用 ISSR分子标记方法对国产黄独 14个居群的遗传多样性进行了研究。
形态学特征在薯蓣属的分类学研究中具有重要的作用[10],叶表皮结构特征 (如气孔、表皮毛及其他附属物的特征)被认为是较稳定的演化特征[11-16],在植物分类学或系统学研究方面具有重要意义。近年来,叶表皮结构特征在薯蓣属周生翅组 (Sect.EnantiophyllU line)、复 叶 组 〔Sect.Botryosicyos(Hochst.)U line〕及根状茎组 (Sect.StenophoraU line)等类群的系统学研究中得到了很好的应用[17-21],而有关黄独叶表皮形态特征方面的研究报道却较少,仅凌萍萍等[22]利用光学显微镜对黄独叶片表皮细胞及细胞壁的形态、腺体毛及非腺体毛的有无等进行了观察及简要的描述。
由于黄独的种下变异并不仅仅体现在遗传差异上,仅采用分子标记并不完全适用于黄独种下类群的区分,因此,寻找合适的形态特征对黄独的系统分类具有重要意义。作者以国产黄独的 11个野生居群及2个栽培居群为研究对象,并以 2个引自美国的栽培居群为外类群,应用扫描电子显微镜对叶表皮的微形态特征(包括气孔器、表皮毛和气孔周围表皮细胞的微形态特征)进行了观察,探讨了黄独及其种下变异类型的叶表皮微形态特征差异,并据此编制了 15个居群的分类检索表,以期为黄独及其种下类群的系统分类研究提供一定的理论依据。
1 材料和方法
1.1 材料
2001年至 2009年,分别采集 11个野生居群和 4个栽培居群材料(表 1),其中的 2个栽培居群引种自美国北卡罗来纳 (经鉴定为黄独变种D.bulbiferavar.sativaPrain etBurkill),共 15个供试居群,凭证标本信息见表 1。
1.2 方法
分别采集各居群植株中部叶片,于叶片中脉处取4mm×4mm的小块,置于体积分数 95%乙醇中超声波振荡清洗 10m in,再用无水乙醇脱水,挥干后用双面胶将叶片小块平整黏贴于样品台上,置于 IB-3型离子溅射仪(日本 Eiko公司生产)中进行导电处理,导电膜厚 20 nm;然后置于 H itachi S-800型扫描电子显微镜(日本日立公司生产)下观察气孔器 (包括外拱盖内缘类型、旋转方向、长轴长度、密度及深度)、表皮毛 (包括主脉和二级脉表皮毛类型以及叶肉腺毛的长轴和短轴长度及腺毛密度)以及气孔周围表皮细胞(包括平周壁表面纹饰及扩散方式、平周壁上附属颗粒形状及垂周壁形状)的微形态特征;每个性状观察 5~10个视野并选取合适部位进行拍照,每份材料均重复 3次。相关术语均参照 Harris等[23]和Baranova[24]的描述。
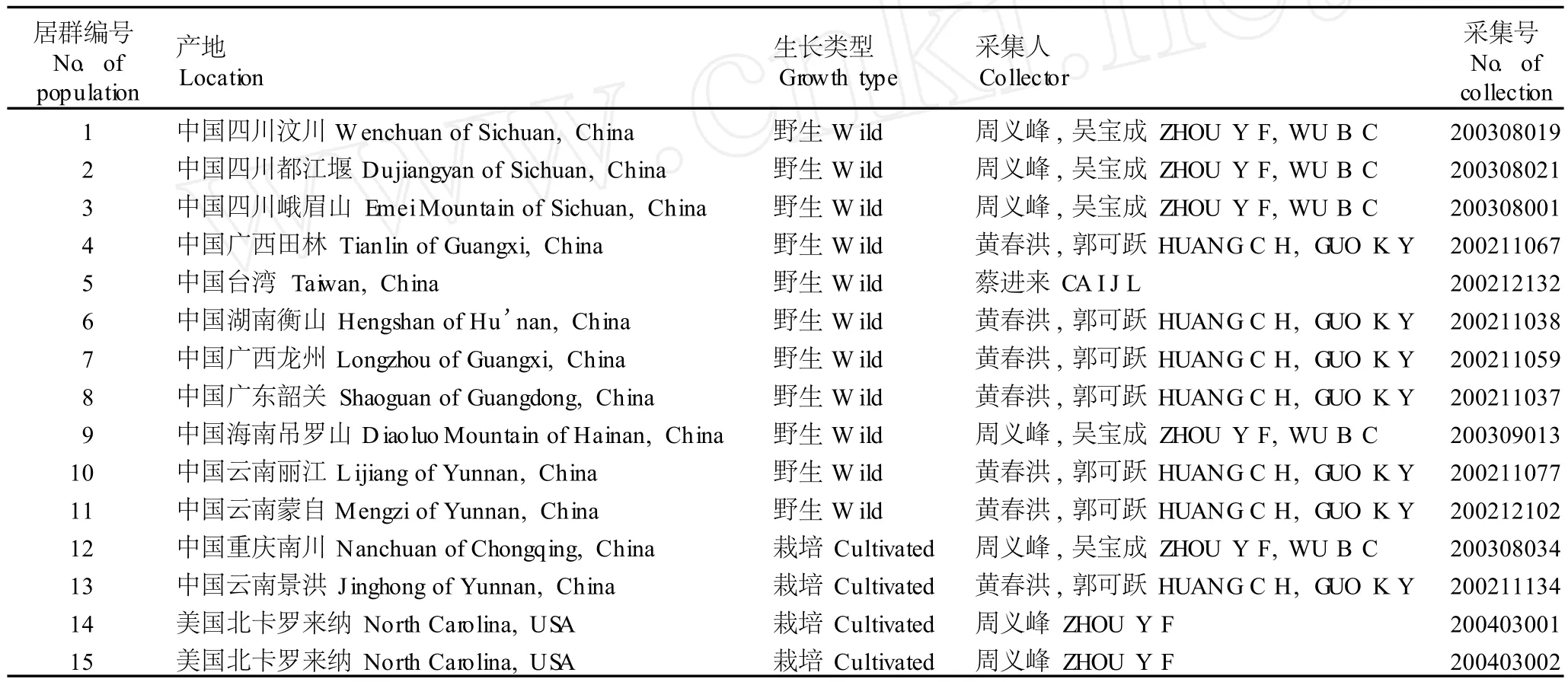
表 1 供试 15个黄独居群的凭证标本基本信息Tab le 1 Basic in form a tion of voucher specim en sof fifteen popu la tion s of D iosco rea bu lbifera L.for tested
2 结果和分析
2.1 黄独各居群叶表皮微形态特征
2.1.1 气孔器的微形态特征 供试的 15个黄独居群的气孔器均为不定型,副卫细胞多数为 3~5个且与周围细胞界线模糊。除云南景洪居群的气孔器外拱盖内缘为光滑型外 (图版Ⅰ-1),其余居群的气孔器外拱盖内缘均为浅波状(图版Ⅰ-2)。表 2结果显示:云南蒙自居群的气孔器旋转方向为不定向 (即左旋和右旋同时存在,见图版Ⅰ-3),广西田林和龙州、云南景洪和丽江 4个居群的气孔器旋转方向为左旋(图版Ⅰ-4),其余居群的气孔器旋转方向均为右旋(图版Ⅰ-5);15个居群气孔长轴长度的变化幅度为18.59~31.93μm,其中四川汶川居群的气孔长轴长度最短,云南蒙自居群的气孔长轴长度最长;15个居群的气孔密度变化幅度为 86~356 mm-2,其中四川峨眉山居群和引种自美国北卡罗来纳的编号 14的栽培居群的气孔密度最小,而海南吊罗山居群的气孔密度最大;各居群间的气孔深度也有明显的差异。
2.1.2 表皮毛的微形态特征 黄独叶片的表皮毛均存在于下表皮,主要有乳突和多细胞头单细胞柄腺毛2种类型。乳突主要分布于主脉 (图版Ⅰ-6),而腺毛则主要分布于二级脉 (图版Ⅰ-7)及叶肉 (图版Ⅰ-8)。表 3结果显示:云南景洪、蒙自和丽江 3个居群叶片同时具有乳突和腺毛 2种类型的表皮毛;海南吊罗山和广西龙州 2个居群叶片的主脉和二级脉没有表皮毛,引种自美国北卡罗来纳的编号 15的栽培居群叶片的主脉和二级脉也没有表皮毛;广西田林、台湾、广东韶关和重庆南川 4个居群的叶片有乳突但没有腺毛,其余居群叶片的主脉和二级脉没有乳突仅有腺毛。
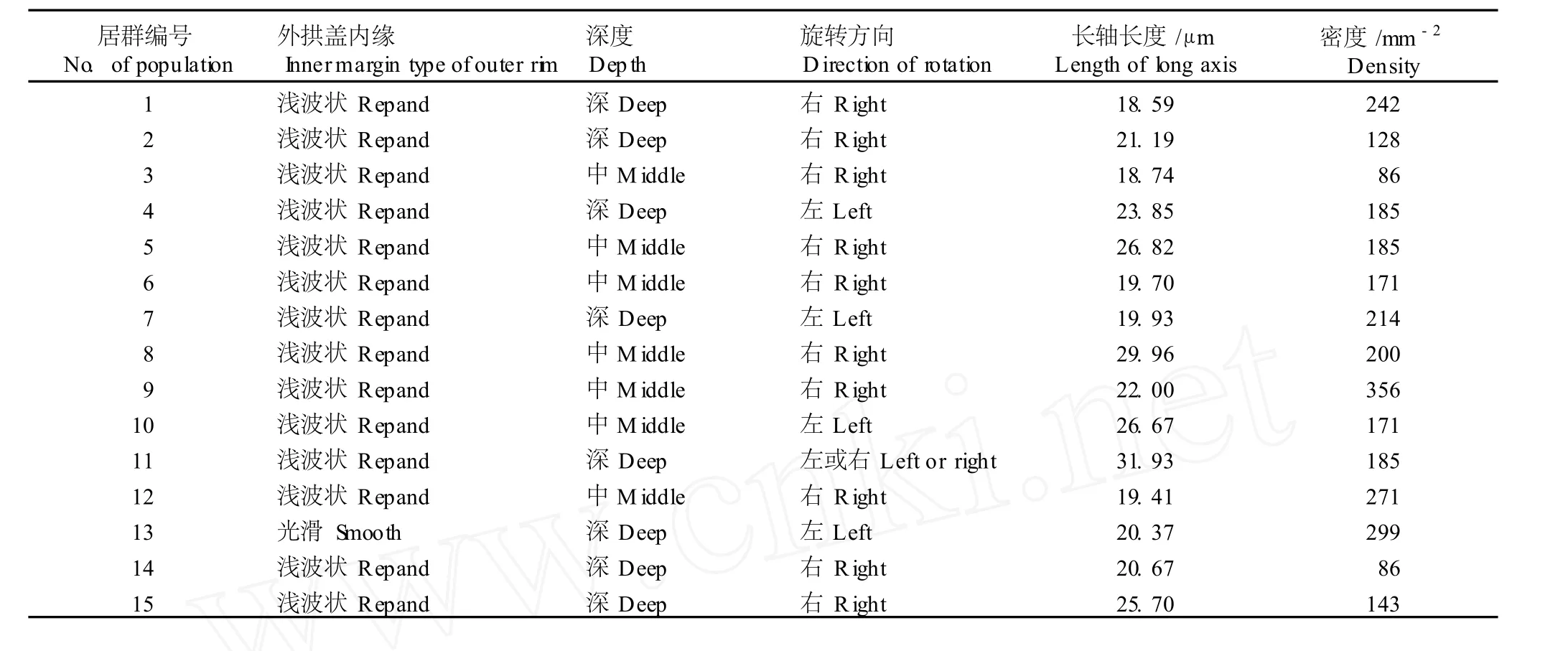
表 2 15个黄独居群气孔器微形态特征比较Tab le 2 Com par ison ofm icro-m orpholog ica l characters of stom a ta lappara tusof D iosco rea bu lbifera L.from fifteen popu la tion s
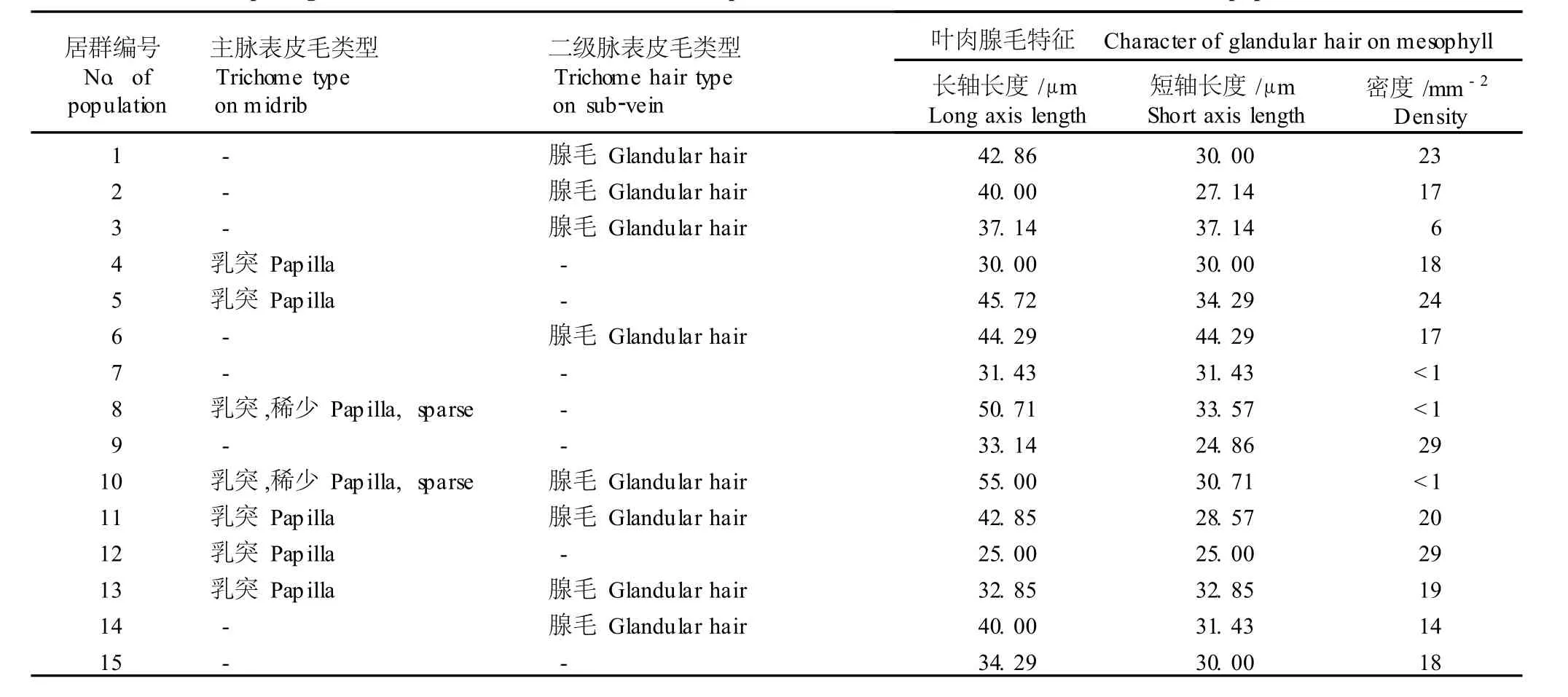
表 3 15个黄独居群叶片各部位表皮毛的微形态特征1)Tab le 3 M icro-m orpho log ica l characters of tr ichom e on d ifferen t par ts of D iosco rea bu lbifera L.lea f from fifteen popu la tions1)
15个供试居群叶片的叶肉部位均具有腺毛,但各居群叶肉腺毛的特征有一定的差异(表 3)。15个供试居群叶肉腺毛长轴和短轴的长度变化幅度分别为25.00~55.00和 24.86~44.29μm;大部分居群腺毛密度(图版Ⅰ-9)的变化幅度为 6~29mm-2,其中四川峨眉山居群叶肉腺毛密度最小,重庆南川和海南吊罗山居群叶肉腺毛密度最大;此外,因广西龙州、广东韶关和云南丽江居群叶肉腺毛极少,故没有统计这 3个居群叶肉腺毛的密度。
2.1.3 气孔周围表皮细胞的微形态特征 黄独不同居群叶片气孔周围表皮细胞的形态特征明显不同(表 4)。15个居群叶片气孔周围表皮细胞垂周壁形状有平直 (图版Ⅰ-10)、平直隆起 (图版Ⅰ-11)、平直脊状隆起(图版Ⅰ-12)、弯曲 (图版Ⅱ-1)、弯曲脊状隆起 (图版Ⅱ-2)和弯曲拱状隆起 (图版Ⅱ-3)等 6种形态类型,各居群无特殊规律。
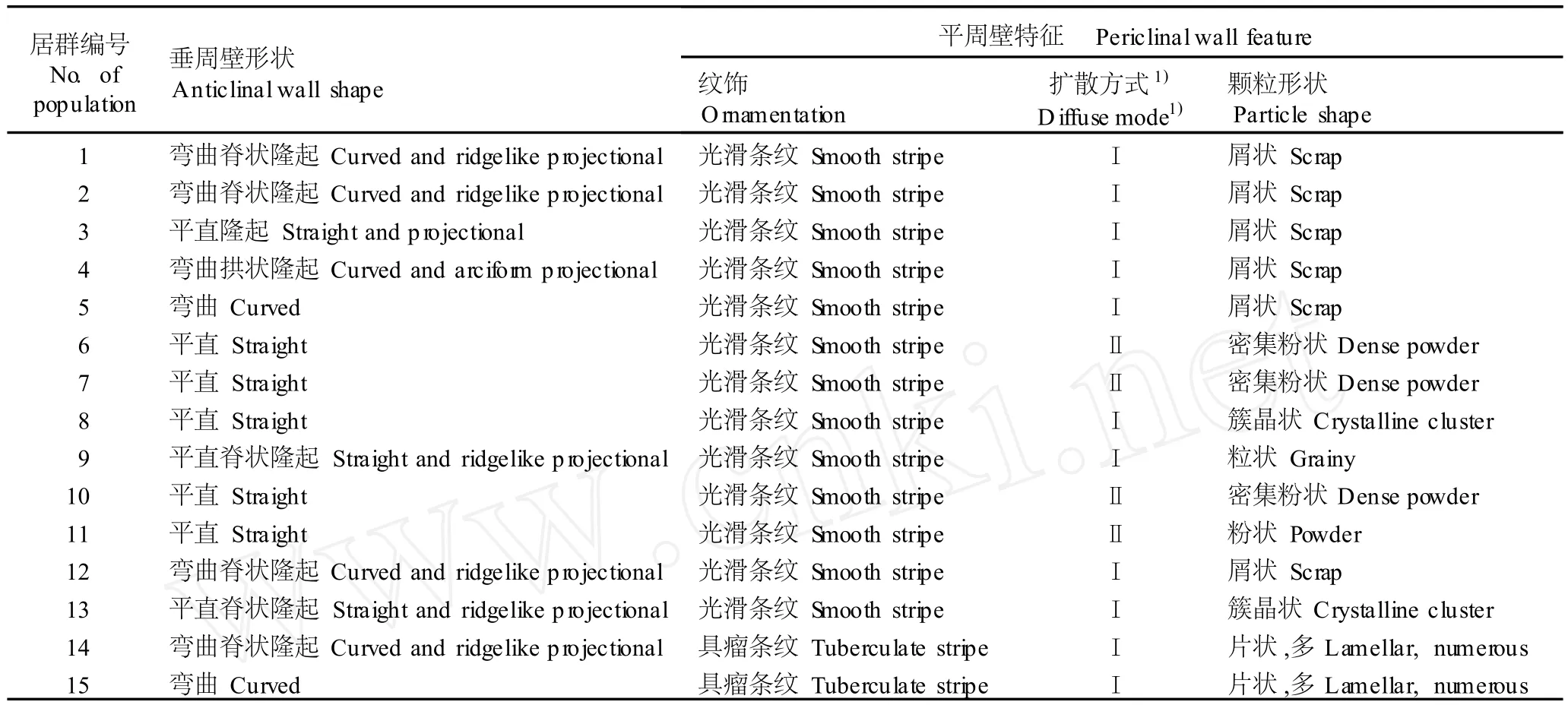
表 4 15个黄独居群叶片气孔周围表皮细胞的微形态特征Tab le 4 M icro-m orpho log ica l characters of ep iderm is cellsaround stom a ta on lea f of D iosco rea bu lb ifera L.from fifteen popu la tion s
15个居群叶片气孔周围平周壁表面纹饰有具瘤条纹和光滑条纹 2种形态,其中,来自美国北卡罗来纳的 2个居群为具瘤条纹 (图版Ⅱ-4),其余各居群均为光滑条纹(图版Ⅱ-5,6)。气孔周围平周壁条纹的扩散方式也有 2种形式:一种为不环绕气孔且四方扩散(图版Ⅱ-5),气孔两端的条纹与气孔长轴平行,两侧的条纹与气孔长轴垂直,引种自美国北卡罗来纳的 2个居群以及产自广东韶关、云南景洪、四川峨眉山、四川汶川、四川都江堰、重庆南川、广西田林、台湾和海南吊罗山的居群均属于该类型;另一种为环绕气孔 2~3周后扩散,但扩散方向不定 (图版Ⅱ-6),产自广西龙州、云南丽江和蒙自及湖南衡山的居群属于这种形态类型。
15个黄独居群叶片表皮细胞平周壁上的颗粒形状有簇晶状、屑状、密集粉状、粉状、粒状和片状 6种形态(图版Ⅱ-7~12)。其中,湖南衡山、广西龙州及云南丽江 3个居群为密集粉状,云南蒙自居群为粉状,海南吊罗山居群为粒状,引种自美国北卡罗来纳的 2个居群为片状,广东韶关和云南景洪 2个居群为簇晶状,其余 6个居群均为屑状。
从上述的观察和分析结果可见:黄独不同居群叶表皮微形态特征的多态性较高。地理位置相近的居群相似性较高。如产自四川的 3个黄独野生居群在叶表皮毛特征、表皮细胞颗粒形状及平周壁表面纹饰、气孔器旋转方向等特征上存在高度相似性,仅在叶肉腺毛密度、气孔深度和密度等方面有一定的差异;引自美国北卡罗来纳的 2个居群也高度相似,仅在二级脉有无腺毛这一特征上存在差异。气孔周围表皮细胞平周壁上的颗粒类型在各居群间有着较大的、稳定的差异。
2.2 基于叶表皮微形态特征的 15个黄独居群的分类检索表
根据叶表皮微形态特征 (主要包括气孔器、表皮毛和表皮细胞的微形态特征),编制出供试 15个黄独居群的分类检索表。

基于叶表皮微形态特征的 15个黄独居群的检索表
3 讨 论
目前,相关研究者对黄独的分类问题还存在一定的争议。《Flora of China》[4]正式收载的黄独仅 1种;而 Prain等[3]则认为:除黄独外,广东、云南腾冲及四川茂汶地区还分布有黄独的 3个变种;周义峰等[5-6]认为:中国不但分布有白金山药、雷公薯和大花黄独等变种,还分布有少花黄独和枝花黄独等新拟变种。因此,对黄独的种下分类研究需要借助于多学科的证据。
15个供试黄独居群的外部形态特征有明显差异,主要区别特征如下:湖南衡山、广西龙州、云南丽江和云南蒙自 4个居群植株的雄花有穗状花序 3~6个,生于叶腋,其中 1枝突化为花序枝;广东韶关居群和云南景洪栽培居群植株的零余子少而小,雌雄花序少而短;引种自美国北卡罗来纳的 2个栽培居群的零余子大,表面白色而光滑;海南吊罗山居群植株的叶基平截或三角状深心形,果实较短;其余居群形态特征与黄独原变种基本相同,即:零余子紫棕色、表面有圆形斑点,雄花花序穗状、下垂、常数个丛生于叶腋,叶片宽卵状心形或卵状心形[1]。
依据植株的形态特征可以将供试的 15个黄独居群分为 5种形态类型。其中,广东韶关和云南景洪居群为一类,特征为零余子少而小,雌雄花序少而短;湖南衡山、广西龙州、云南丽江和云南蒙自居群为一类,特征为雄花有穗状花序 3~6个、生于叶腋,其中 1枝突化为花序枝。这 2个类型的黄独分别被周义峰等[6]拟定为黄独变种,拟定名为少花黄独和枝花黄独。海南吊罗山黄独植株的叶基平截或三角状深心形,果实较短,周义峰等[5-6]认为应该为中国分布的变种雷公薯。15个居群植株叶片表皮细胞平周壁上的颗粒有簇晶状、屑状、密集粉状和粉状、粒状、片状等6种形态,分别对应于少花黄独 (拟)、黄独原变种、枝花黄独(拟)、雷公薯和引种自美国北卡罗来纳的变种D.bulbiferavar.sativa。因此,叶表皮微形态特征揭示了这些黄独种下类群存在的可能性,可为黄独的种下分类提供科学的依据。
黄独 15个居群植株叶片的气孔器、表皮毛和表皮细胞微形态特征的综合分析结果表明:地理分布相近的居群叶表皮微形态特征相似程度较高,一定程度上反映出环境的饰变作用。根据叶表皮微形态特征的多样性可对供试的 15个黄独居群加以区分,表明叶表皮微形态特征对黄独种下类群的划分具有一定的帮助。但这些形态特征是否稳定?是否还具有更为稳定且特异的形态特征?这些疑问则有待进一步深入研究。
[1]中国科学院中国植物志编辑委员会.中国植物志:第十六卷第一分册[M].北京:科学出版社,1985:88.
[2]吴征镒,路安民,汤彦承,等.中国被子植物科属综论[M].北京:科学出版社,2004:212-217.
[3]Prain D,Burkill IH.An accountof the genusDioscoreain the East. Part I.The specieswhich tw ine to the left[J].Annalsof the Royal Bo tanic Garden(Calcutta),1936,14:1-20.
[4]D ing Z Z,GilbertM G.D ioscoreaceae[M]∥W u Z Y,Raven PH. Flora of China:Vol.24.Beijing:Science Press,2000:286-287.
[5]Zhou Y F,Xu Z L,Hang Y Y,et al.Dioscoreabulbiferavar.albotuberosa(Dioscoreaceae),anewvarietyfromYunnan,China [J].Novon,2008,18(4):555-557.
[6]周义峰.中国薯蓣属黄独分类及遗传多样性研究[D].南京:江苏省·中国科学院植物研究所,2006:22-44.
[7]TerauchiR,Terachi T,Tsunewaki K.Physicalmap of chlorop last DNA of aerial yam,DioscoreabulbiferaL.[J].Theoretical and App lied Genetics,1989,78:1-10.
[8]Terauchi R,Terachi T,Tsunewaki K.Intraspecific variation of chlorop last DNA inDioscoreabulbiferaL.[J].Theoretical and App lied Genetics,1991,81:461-470.
[9]郑玉红,夏 冰,杭悦宇,等.黄独遗传多样性研究[J].西北植物学报,2006,26(10):2011-2017.
[10]杭悦宇,徐珞珊,史德荣,等.中国薯蓣属植物地下茎淀粉粒形态特征及其分类学意义[J].植物资源与环境学报,2006, 15(4):1-8.
[11]Irvine FR.W oody Plantsof Ghana[M].London:Oxford University Press,1961:26-28.
[12]M etcalfe C R,Chalk L.Anatom y of D icotyledons[M].2nd ed. London:C larendon Press,1979:97-162.
[13]杨传友,苗 良.苹果叶片气孔的研究[J].山东农业大学学报:自然科学版,1998,29(1):8-14.
[14]韦存虚,谢佩松,周卫东,等.麦冬、土麦冬和阔叶土麦冬叶表皮形态结构的观察 [J].植物资源与环境学报,2008,17 (4):9-15.
[15]高召兰,李法曾.山东广义苦荬菜属(菊科)叶表皮微形态的研究[J].广西植物,2007,27(3):435-439.
[16]高德民,樊守金.姜品种叶片形态学研究[J].热带亚热带植物学报,2007,15(4):324-332.
[17]贾成森,徐增莱,程树兵,等.山药不同种质资源的叶下表皮微形态研究[J].广西植物,2009,29(5):607-613.
[18]蒋道松,周朴华,梁曾恩妮,等.盾叶薯蓣四倍体与二倍体叶表皮及气孔器显微结构的比较[J].广西植物,2008,28(4): 427-430,450.
[19]杭悦宇.中药萆薢的原植物、鉴定及质量评价研究[D].南京:中国药科大学生药学院,2007:48-55.
[20]郑玉红.中国薯蓣属周生翅组的系统学研究[D].南京:南京农业大学生命科学学院,2006:11-82.
[21]史德荣.中国薯蓣属复叶类群的系统学研究[D].南京:江苏省·中国科学院植物研究所,2006:22-23.
[22]凌萍萍,吴竹君,秦慧贞.薯蓣属(DioscoreaL.)叶表皮气孔类型在分类上的意义[M]∥南京中山植物园研究论文集编辑组.南京中山植物园研究论文集:1981.南京:江苏科学技术出版社,1982:9-16.
[23]Harris JG,HarrisM W.图解植物学词典[M].王宇飞,赵良成,冯广平,等,译.北京:科学出版社,2001:277-302.
[24]Baranova M.Princip les of comparative stom atographic studies of flowering p lants[J].The BotanicalReview,1992,58(1):49-99.
