泰山虫草子实体形态特征与 rDNA变异位点分析
2011-01-24曲志才苏延友
王 波,曲志才*,苏延友
(1.曲阜师范大学生命科学学院,山东曲阜 273165;2.泰山医学院药学院,山东泰安 271000)
虫草属 (Cordyceps)是一类虫生真菌,隶属麦角菌科 (Clavicipitaceae),广布于全世界。一般认为该属拥有 400多种[1],我国现有记载 139个名称[2],其中九州虫草 (Cordyceps kyushuensis)在蒙山[3]和泰山均有发现,而泰山虫草 (Cordyceps taishanensis)[4]仅见于泰山南麓。目前对两株虫草的研究主要集中在有效成分及人工培养方面,其形态学、分类地位以及分子生物学方面的研究还鲜见报道。
关于虫草的分类方法主要有两种,一是 Kobayasi分类系统,该系统主要侧重于虫草子座、可孕部分、子囊、子囊壳、子囊孢子等形态特征的描述,尤其是子囊上部、子囊孢子和次生孢子的形态及大小。但虫草子囊孢子发育存在阶段性,因此给依据形态特征鉴定虫草带来了困难[5]。探讨用分子生物学技术测定虫草不同个体发育阶段及虫草无性型和有性型的遗传统一性,在虫草分类中具有明显的针对性。Sung等通过对虫草属 91个种类的 nrSSU,nr LSU,β -tubulin,EF-1α,RPB1andRPB2,m tATP67个位点进行多基因分析,重建了虫草属的系统发育关系[6],为现有的分类系统提供了重要的分子生物学信息。
核糖体DNA是细胞中最古老的分子之一,被广泛地应用于真菌系统发育和进化类型的分析。真核生物的核糖体 DNA以串联重复方式出现在染色体基因位点上,每个序列单位含有编码 18S,5.8S和28S的 rRNA,两个内转录间隔区 (ITS1和 ITS2)以及外转录间隔区 (ETS)。编码 18S,5.8S和 28S的序列选择压力大,为高度保守区段,进化较缓慢,可以用于亲缘关系较远的有机体;ITS为进化较快的中度保守序列,其保守性基本上表现为种内相对一致,种间差异比较明显,并且具有各重复单元同步进化的特点,因此可以用于比较关系较近的种或亚种[7]。本文首次对我省两株虫草的核糖体 DNA的18S,28S、ITS序列进行测序,通过对特定区段变异位点的比较分析,并结合形态学上的指标,以期从理论上探讨两株虫草分类上的关系,为虫草资源的合理开发利用奠定基础。
1 材料与方法
1.1 材料
供试材料虫草 T1和 T2均采自泰山南麓槐、栎树阔叶林下,海拔 700~900 m。其中虫草 T1经山东大学微生物研究所张长铠教授和山东省农科院谢思密研究员初步鉴定为九州虫草(Cordyceps kyushuensis)。
1.2 虫草形态观察
冷冻切片显微观察:取虫草子座可孕部分置蒸馏水中浸泡使其充分吸胀,采用包埋法制备横切薄片,置光学显微镜下观察。子座扫描电镜观察:虫草可孕部分自然风干后,在冷场发射扫描电镜 (JS M-6700F)下观察其超微结构。
1.3 虫草基因组 DNA的提取
采用 CTAB法提取总 DNA[8],略加修改。烘干菌丝体在 TE缓冲液 (10 mM Tris-HCl,1 mM EDTA,pH8.0)中浸泡 12 h。称取 30 mg菌丝体加10%PVP,液氮中研磨,加入 750μL提取缓冲液(0.25 M NaCl,0.25M Tris-HCl,50 mM EDTA,2%β-巯基乙醇),置冰浴 30 min。7 000 g,4℃离心10 min。取上清液,加 750μL含 VitC干粉末的 2.5×CTAB,65℃水浴 2 h,每隔 15 min颠倒混匀 1次。10 000 g离心 10 min(下同),取上清液,加等体积酚 /氯仿 /异戊醇 (25∶24∶1),离心后取水相加等体积氯仿 /异戊醇 (24∶1),再离心,取水相加 1/20体积 5 M NaCl和 0.8倍体积异丙醇,4℃离心,70%乙醇洗涤沉淀。以 TE溶解沉淀,经 RNase A处理,最后溶于 20μL TE中,20℃保存备用。
1.4 PCR引物设计及扩增反应体系
参照 White等人[9]所设计的 ITS区、18S、28S rDNA的引物序列 (表1)。扩增反应体系组成如下:超纯水 17.35μL,10×Buffer(含 Mg2+)2.5μL,2.5 mmol/L dNTP 2.0μL,50μmol/L引物各 1μL,Taq DNA聚合酶 0.15μL,模板 1.0μL,总体积 25μL。
扩增程序:94℃预变性 5 min;94℃变性 30 s,退火 1 min(复性温度见表1),72℃延伸 1.5 min,35个循环,72℃再延伸 10 min。
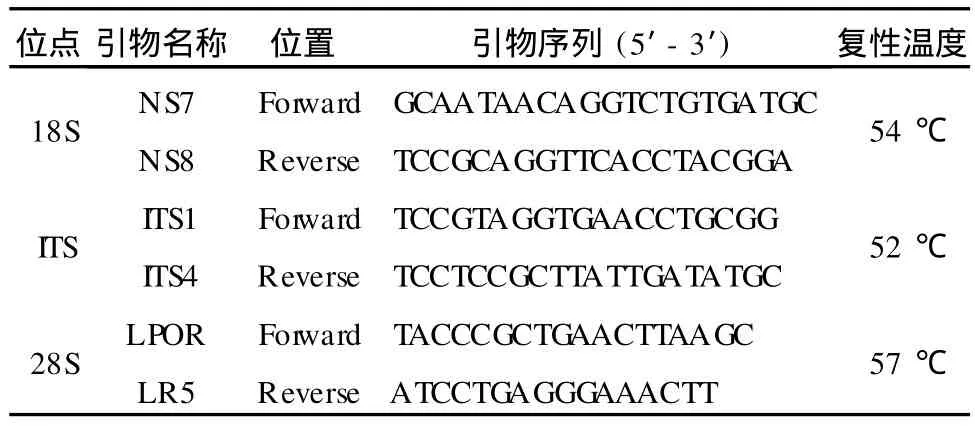
表1 PCR引物的核苷酸序列及复性温度
1.5 特异扩增片段的回收和克隆及测序分析
PCR产物经琼脂糖凝胶 (1.5%质量浓度)电泳分离,302 nm紫外灯观察,割取目的 DNA条带。以DNA胶回收试剂盒 (大连宝生物)回收特异条带。按 pMD18-T载体试剂盒说明进行连接反应,转化大肠杆菌DH5α感受态细胞,继而进行蓝白斑筛选。挑取单个白斑菌落,37℃过夜培养。经菌落 PCR进一步鉴定阳性克隆后,送上海基康生物技术公司测序。
采用 ClustalX对测定序列进行排序,序列间插入适当的空位,以达到最大的同源性;应用 DNASP4进行数据分析。
2 结果与分析
2.1 虫草 T2子实体形态特征
虫草子座着生于豆天蛾幼虫体上,虫体土黄褐色,表面覆盖白色菌丝,由这些菌丝形成菌索。子座由虫体头部发出,呈圆柱形、椭圆形或棍棒形,橙红色,高 2~8 cm,直径 2~6 mm,单生,直立不分枝,前部膨大而粗糙,由很多子囊壳形成的微小颗粒(图1)。

图1 虫草 T2子座及豆天蛾幼虫虫体
子实体可孕部分横切薄片,显微镜下观察,子囊壳呈卵圆形 500~700μm ×200~300μm,在子座内呈辐射状排列于四周,内生 (图2)。该结果与安秀荣等[10]描述的情况非常相似。通过扫描电镜观察可以看出,子实体的子囊壳均以半埋方式着生于子座上,尖端部突出于子座表面,似“山丘”状,正面高低不平 (图3)。

图2 虫草 T2子实体光镜横切片观察

图3 虫草 T2子实体外部形态扫描电镜观察
2.2 虫草基因组 DNA的电泳检测
提取的虫草 T1和 T2基因组 DNA,经琼脂糖凝胶电泳检测,可以清晰地看到基因组 DNA大小约为23 kb(图4),且带型完整、明亮,可以作为 PCR扩增的模板。
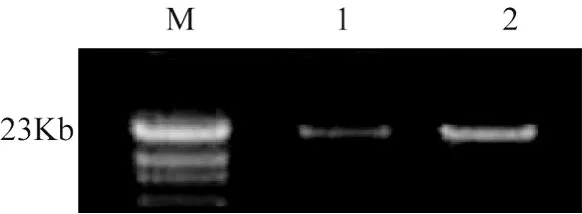
图4 虫草核基因组DNA电泳图谱
2.3 PCR扩增产物的电泳检测
运用 PCR技术从两株虫草核 DNA中均扩增出rDNA的 ITS序列、18S序列和 28S序列,扩增产物经琼脂糖凝胶电泳检测,可以看到大约 500 bp(ITS)、300bp(18S)、900bp(28S)的条带 (图5),大小与预期结果一致。
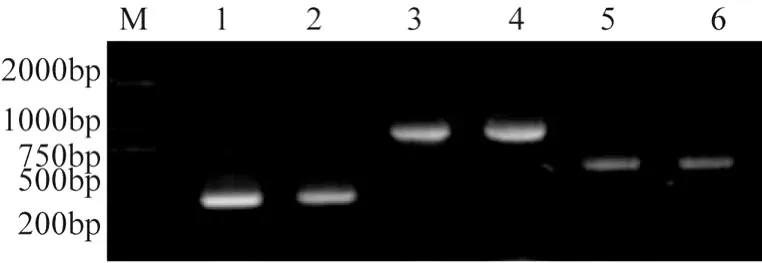
图5 PCR扩增产物电泳图谱
2.4 序列分析
通过对两株虫草的 ITS区、18S区、28S区的序列进行比对分析发现,通用引物 NS7、NS8扩增出的18S片段长度均为 372 bp,共 3个变异位点,占测序总位点的 0.807%。其中发生碱基转换的有 2个,占测序总位点的 0.537%;发生颠换 1个,占测序总位点的 0.269% 。转换与颠换的比值为 2∶1,同源性高达 99%(表2)。
ITS区测序表明,虫草 T1长度为 567 bp,而 T2虫草 565 bp,共发现 6个变异位点,占测序总位点的1.058%。碱基转换的有 3个,占测序总位点的0.529%;发生颠换的有 1个,占测序总位点的0.1764%。缺失 2个。转换与颠换的比值为 3∶1,同源性高达 99%(表3)。
虫草 T1和 T2的 28S区长度为 926 bp,共发现9个变异位点,占测序总位点的 0.971%。碱基转换7个,占测序总位点数的 0.756%;碱基颠换 2个,占测序总位点数的 0.216%。转换与颠换的比值为5∶1,同源性达 99%。
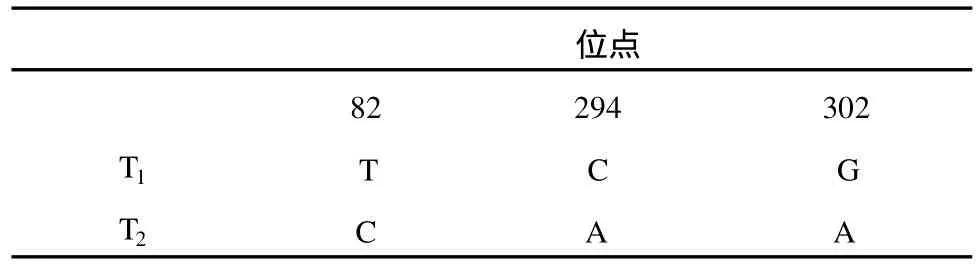
表2 18S区段序列核苷酸差异位点

表3 ITS区段序列核苷酸差异位点
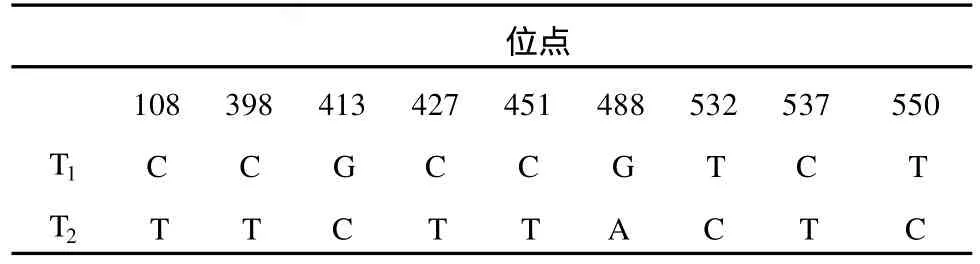
表4 28S区段序列核苷酸差异位点
3 讨 论
小林义雄与清水大典在有关冬虫夏草菌的专著中记载了寄生在日本鳞翅目幼虫上的九州虫草特征:子座椭圆形至圆柱形、子囊壳半裸生、卵形、长410~580μm ×210~330μm[11]。郭英兰在沈阳采集的九州虫草标本,其形态学特征与小林义雄描述的非常相似[12]。本文所研究的虫草 T1和 T2,经显微结构和超显微结构观察,无论子实体的外部形态,还是子囊壳的形态、排列方式等特征与小林义雄、郭英兰等的描述都非常相似。因此,根据 Kobayasi分类系统可以初步判断泰山虫草与九州虫草应归属于同一种。
通过对 rDNA 18S,28S,ITS序列的比较还可以发现,同一引物扩增出的片段长度基本一致,两虫草的同源性较高。在 18S,28S高度保守区,两虫草分别有 3个和 9个碱基差异,其变异位点占总测序位点的 0.807%和 0.971%,在较能体现菌物遗传多样性的 ITS区段,两株虫草的碱基差异也只有 6个,占测序总位点 1.058%,两者所测核苷酸序列同源性高达 99%,其 18S、28S、ITS序列基本没有差异。结合两株虫草的形态特征和分子生物学证据,初步推断泰山虫草与九州虫草为同物异名。虫草无性型的确定及分子系统学研究正在进行中。
[1] Kuo H C,Su YL,YangH L,et al.Identification of Chinesemedicinal fungus Cordyceps sinensis by PCR-single-stranded conformation polymorphis m and phylogenetic relationship[J].J Agric Food Chem,2005,53(10):3963-3968.
[2] 宋 斌,林群英,李泰辉,等.Known species of Cordyceps from China and their distribution[J].Journal of Fungal Research,2006,4(4):10-26.
[3] 凌建亚.九州虫草无性型鉴定暨虫草属分子系统学及虫草素与DNA直接作用机理研究[D].济南:山东大学,2003.
[4] LiuB,Yuan P G,Cao J Z.A new speciesof Cordyceps from China[J].ActaMycologia Sinica,1984,3(4):192-195.
[5] 梁宗琦.我国虫草属真菌研究开发的现状及思考[J].食用菌学报,2001,8(2):53-62.
[6] Sung G H,Sung JM,NigelL H J,et al.A multi-gene phylogeny of Clavicipitaceae(Ascomycota,Fungi):Identification of localized incongruence using a combinational bootstrap approach[J].Molecular Phylogenetics and Evolution,2007,44:1204-1223.
[7] 刘作易.虫草属及其无性型关系研究[D].华中农业大学博士论文,1999:1-7.
[8] 郭宝林,林 生.丹参干叶片的DNA提取[J].中草药,2002,33(5):418-420.
[9] White T J,Bruns T,Lee S,et al.Amplification and direct sequencing of fungal ribosomalRNA genes forphylogenetics.In PCR Protocols:A guide to methods and applications[M].San Diego:Academic Press,1990:315-322.
[10] 安秀荣,杨永恒.泰山虫草菌调查简报[J].食用菌,2003,3:4.
[11] 小林义雄,清水大典.冬虫夏草菌图谱[M].大坂:保育社,1983.
[12] 郭英兰,李春燕.中国一新记录种—九州虫草[J].菌物系统,2000,19(2):296.
