黄绿蜜环菌分批发酵菌丝动力学模型
2010-11-10余梅
余 梅
(青海普兰特药业有限公司,青海西宁810007)
黄绿蜜环菌分批发酵菌丝动力学模型
余 梅
(青海普兰特药业有限公司,青海西宁810007)

黄绿蜜环菌,分批发酵,动力学模型,拟合
黄绿蜜环菌[Armillaria luteo-virens(Alb.et Schw. Fr.)Sacc]属于白蘑科(Tricholomataceae);蜜环菌属(Armillaria),俗称黄蘑菇,是一种名贵食用菌[1]。主要分布于青海省的海北(祁连、海晏、刚察),黄南(泽库、河南),海南(共和、贵德、兴海),果洛(玛沁、甘德、久治),玉树等地。黄绿蜜环菌子实体肥厚,味道鲜美,营养丰富。干菇中粗蛋白含量高达39%,此外还含有19种氨基酸等营养物质[2-7]。由于黄绿蜜环菌具有较高的营养价值,尤其是蛋白质含量很高,优异的活性蛋白使其具有较好的防癌、抗癌、降血糖和提高免疫力的功效。可是黄绿蜜环菌至今不能进行人工种植,只能依靠采摘野生菇体满足日益增长的市场需求,加上青藏高原恶劣的生态环境,使得野生资源相当贫乏,给采摘工作带来很大困难。因此采用液体深层发酵菌丝粉来代替子实体进行资源的开发利用,既不违背对高原珍稀资源保护的大趋势,又可以创造高原特色资源开发的经济价值,还易于实现工业化大生产。为此实验针对黄绿蜜环菌在摇瓶培养过程的特性和基本规律,探讨了该菌在70L发酵罐中的液体培养动力学,建立了用以描述黄绿蜜环菌液体培养菌丝体过程的菌体生长、底物消耗的动力学模型,并研究了整个发酵过程各动态参数的变化情况,同时得出相关动力学曲线,旨在为黄绿蜜环菌的液体大规模培养研究提供理论基础。
1 材料与方法
1.1 材料与仪器
菌种 黄绿蜜环菌(Armillaria luteo-virens Sacc),由青海大学农牧学院农学系提供;葡萄糖、磷酸氢二钾、磷酸二氢钾、硫酸镁等 分析纯;酵母膏购于上海医药集团;固体斜面培养基 PDA培养基;液体种子培养基 葡萄糖15g/L,酵母膏5g/L,磷酸氢二钾 1g/L,磷酸二氢钾 0.5g/L,硫酸镁0.3g/L,pH自然;发酵培养基 葡萄糖34.3g/L,酵母膏1.91g/L,磷酸氢二钾1g/L,磷酸二氢钾0.5g/L,硫酸镁0.45g/L。
FA1004型电子天平 上海精科天平;HWY-111恒温培养摇床 上海智城分析仪器制造有限公司、真空冷冻干燥机 美国Svant公司;手提式压力蒸汽灭菌器 上海博迅实业有限公司医疗设备厂;低LXJ -IIB速大容量多管离心机 上海安亭科学仪器总厂;101-3B型电热鼓风干热箱 上海市实验仪器总厂;GUJS7A-70C型发酵罐 镇江东方生物工程设备技术公司;KQ-C型全自控蒸汽发生器 上海奉贤协新机电厂;V-0.25型空气压缩机 沈阳市达力压缩机厂。
1.2 实验方法
1.2.1 种子活化 将黄绿蜜环菌菌丝块接种于PDA斜面培养基上,28℃下培养6d。共活化2次。
1.2.2 种子扩大培养 500mL三角瓶中装10颗小玻璃珠,干热杀菌后装入种子培养基100mL,湿热灭菌后接种五块 0.5cm左右的菌块,在 27±1℃下,200r/min的摇床上振荡培养4d左右。
1.2.3 分批发酵 将液体种子培养基合并后接入70L发酵罐,接种量为15%,发酵温度27±10℃,搅拌转速250r/min,通气量为1.2L/min,pH由中间取样后流加碱来控制。
1.2.4 生物量的测定[8]发酵液在3000r/min的条件下离心20min,将得到的菌丝体多次洗涤,最后至菌丝体于60℃条件下干燥至恒重。
1.2.5 还原糖的测定 3,5-二硝基水杨酸(DNS)法[9]。
2 结果与分析
2.1 黄绿蜜环菌菌丝发酵过程代谢变化
菌丝形态和发酵过程有关联[10],结合菌丝形态变化与图1所示的各种代谢指标变化情况可以看出,在0~12h发酵处于延迟期,菌丝球(直径4mm左右)在剪切力作用下由球状逐渐变为半球状,葡萄糖消耗与菌体浓度增长缓慢;12~36h进入对数期,不规则的菌丝球碎片逐渐形成大量菌丝体包裹且松散的菌丝小球(直径2mm左右),残糖浓度迅速下降,菌体生长加速,浓度增加;36~56h处于稳定期,菌丝球逐渐变大(直径3mm左右),且球体密度增大,有些菌体发生了自溶成为空心球,菌丝生长减速;56h后处于衰亡期,菌丝球密实且多数出现自溶空心,菌体浓度几乎没有变化,因菌体自溶和发酵液中各种酶的存在,使大量的胞内多糖释放到发酵液中并迅速被酶降解,发酵液的残糖浓度有所回升。
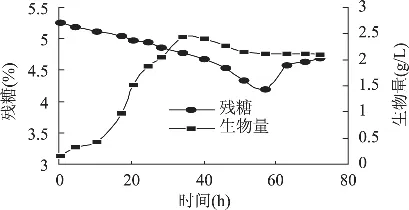
图1 黄绿蜜环菌分批发酵曲线
2.2 Armillaria luteo-virens Sacc菌体生长动力学模型
细胞生长动力学最常用Monod及Logistic方程描述。Monod[11]方程模型基于以下假设建立:a.菌体生长为均衡型非结构式生长,描述细胞生长的唯一变量是细胞的浓度;b.培养基中只有一种基质是生长限制性基质,其它营养成分过量不影响细胞生长;c.将微生物生长视为简单反应,菌体得率为常数,没有动态滞后。显然b、c不符合Armillaria luteo-virens Sacc发酵的特征,采用 Monod方程有偏差。由于Armillaria luteo-virens Sacc菌丝细胞生长曲线为 S形,而Logistic[12]模型是一个典型的S型曲线,用于拟合分批发酵的菌体生长过程具有广泛的适用性,因此,可应用该模型来描述Armillaria luteo-virens Sacc生长动力学特性。在该方程中将比生长速率与尚未利用的负载能力相关联,即生长期细胞的增长速率用式(1)表达:
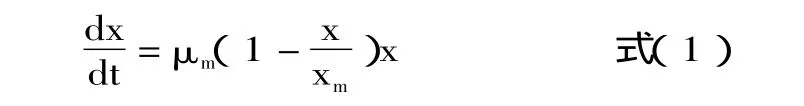
式中,μm代表最大细胞比生长速率,h-1;Xm是基质最大负载的细胞浓度(g dry cell weight·L-1);初始条件为t=0时,X=X0,将式(1)积分得:
将式(2)经对数转化后为:

这样,通过测定X0和μm,可完成上述表达式。首先应用DPS软件算出不同时间的菌含量、残糖含量,然后利用麦夸脱法进行迭代确定模型参数。其模型方程和参数分别见式(5)和表1。
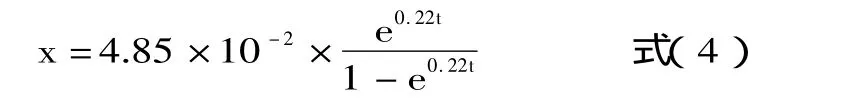
两边取对数得到速率模型:

模型参数经F检验显著性很高(P<0.0001),即模型在99%的概率水平上是非常显著的。t检验结果显示,3个参数中X0不是很显著,只有P=0.287;而最大细胞生物量Xm最为显著(P<0.001),细胞比生长速率μm比较显著(P<0.01)。
图2为菌体生长模型计算值与实验值的比较,可以看出,该菌体生长模型在发酵80h内能很好地描述分批发酵过程的菌体生长情况。在延滞期实验值要高于拟合值,其原因一方面与菌丝细胞的可利用率有关;另一方面因Logistic方程主要反映的是菌体浓度增加和营养物质消耗对自身生长的抑制作用,在延滞期和加速生长期,菌体浓度不高,营养物质丰富,对自身生长的抑制作用不明显,因此拟合情况不如稳定期理想。
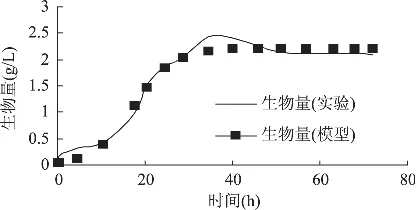
图2 黄绿蜜环菌菌体生长动力学模型值与实验数据拟合
2.3 Armillaria luteo-virens Sacc底物消耗动力学模型
在分批发酵过程中,碳源作为底物,一部分底物用于细胞生长,一部分合成产物,一部分用于能量消耗以维持细胞生命之用。其消耗方程可用类似Luedeking-Piret方程[13]来描述。
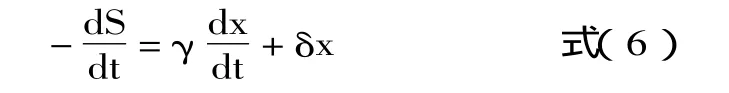
方程(6)中,γ=1/Yx/s和δ=ms。这两个参数可以利用上述方法进行计算。在静止期,细胞的比生长速率为0,细胞生物量达到最大,因此δ可以利用下式进行求解。
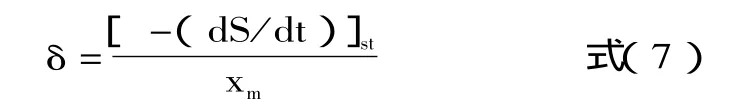
为了求出γ的值,方程(6)经变换得方程:

采用一定的数值求解法,对方程(8)进行变换,得到方程:
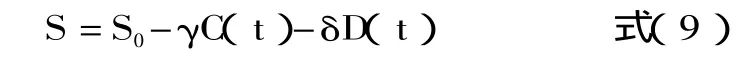
其中C(t),D(t)由下式得到:
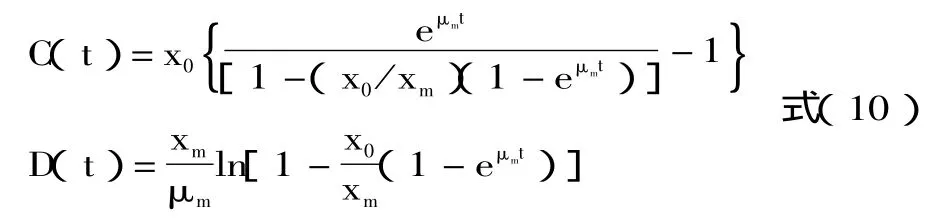
解方程得到优化后的动力学模型参数见表1。其消耗动力学模型见方程(12)。
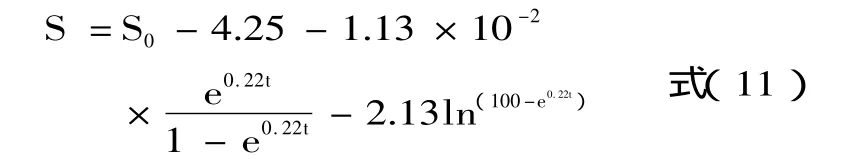
两边取对数得到速率模型:

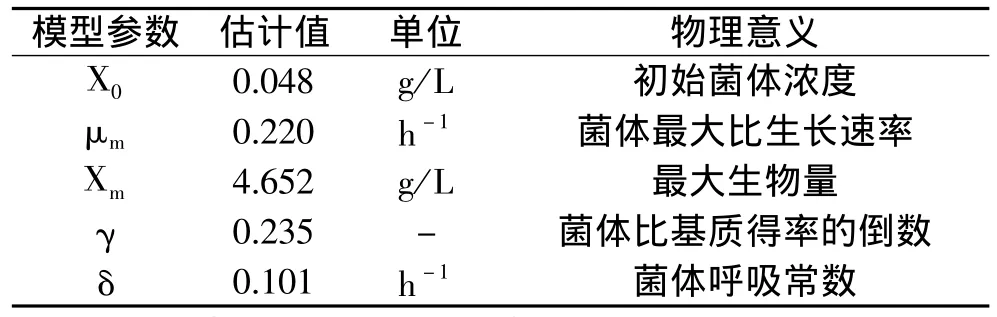
表1 动力学模型参数估计值
底物消耗的模型值与实验值的比较结果见图3,可以看出二者能很好地吻合,所选模型能够很好地描述实验值,这个结论还可以通过模型参数的F检验结果和模型的方差检验结果得到验证(P<0.0001)。t检验分析结果表明模型系数为 P= 0.00129,说明该模型在99%水平上差异显著。
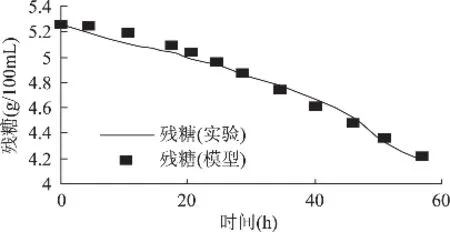
图3 黄绿蜜环菌底物消耗动力学模型与实验数据的拟合
3 讨论
菌体增殖动力学模型在发酵旺盛期有一个生长高峰,随后由于菌体的自溶使菌体生物量趋于稳定;而底物消耗动力学模型拟合度误差主要产生于发酵初期和后期:发酵初期,菌体生长较迅速,对糖的消耗较快,测定出发酵液中的残糖含量低于模型值;发酵后期,即发酵60h以后的预测值与实测值误差较大,主要原因是菌体分泌产生了大量的胞外多糖以及菌体发生自溶,多糖在酶作用下被分解,发酵液的残糖浓度比实际值偏高。但是在选取底物消耗动力学模型时没有考虑到这一点,若以总糖量来评价底物消耗,可以部分消除这种实验偏差。由于发酵体系本身是一个很复杂的体系,尤其是能产多糖和发酵后期具有自溶性的食用真菌,不可避免地会导致测定上的偏差,而且发酵体系本身具有动态性,很难用一种模型去完全拟合菌体生长的整个阶段。鉴于本实验的目的是获得较多菌体生物量,因此底物消耗动力学模型以发酵60h为基准(略去其后过程)进行实验数据的拟合。
[1]刁治民.青海草地黄绿蜜环菌生态学特性及营养价值的研究[J].中国食用菌,1997,6(4):21-22.
[2]李渝珍.青海野生黄绿蜜环菌人工驯化技术途径的探讨[J].青海师范大学学报:自然科学版,2005(1):74-76.
[3]周劲松,熊辉岩.黄绿蜜环菌子实体挥发油的化学成分[J].农业科学与技术:英文版,2008,9(2):90-92.
[4]李海波,吴学谦.青藏高原黄绿蜜环菌纯培养菌种的分离培养及分子鉴定[J].菌物学报,2008,27(6):873-883.
[5]周劲松,盛海彦.青海野生黄绿蜜环菌生态环境及不同生态区菌株间比较[J].食用菌,2007,29(2):9-10.
[6]周劲松,熊辉岩.生长调节剂对黄绿蜜环菌菌丝生长的影响[J].食用菌学报,2007,14(3):44-46.
[7]卢素锦,李军乔.青海黄绿蜜环菌植被类型及伴生植物的初步调查[J].食用菌,2006,28(3):4-5.
[8]柳焕章,王贺祥,刘庆红,等.黄绿蜜环菌菌丝体培养条件初探[J].中国食用菌,2007,24(4):16-19.
[9]大连轻工业学院等和编.食品分析[M].北京:中国轻工业出版社,1994:173-174.
[10]李平作,徐柔,章克昌.灵芝液体发酵过程中菌体形态与胞外多糖产量的关系[J].工业微生物,2000,30(3):20-23.
[11]Bailey JE,Ollis DF.Biochemical Engineering Fundamentals[J].New York:McGrawl HillBook Company,1986:421-440.
[12]MuratElibol,Ferda Mavituna.A kinetic modelfor actinorhodin production by Streptomyces coelicolor A3(2)[J]. Process Biochemistry,1999,34:625-631.
[13]Sinclair CG,Kristiansen B.Fermentation kinetics and modeling[M].Milton Keynes:Open University Press,1987.
[14]Wei GY,Li Y,Du GC,et al.Kinetic models for the effect of temperature on batch glutathione fermentation by Candida utilis[J].Chinese Journal of Biotechnology,2003,19(3):358-363.
[15]VandanaGoswami,A K Srivastava.Fed-batchpro pionic acid production by Propionibacterium acidipro pionici[J].Biochemical Engineering Journal,2000(4):121-127.
Kinetic model for mycelium of Armillaria luteo-virens Sacc in batch fermentation
YU Mei
(Qinghai Plateau Pharmaceutial Co.,Ltd.,Xining 810007,China)
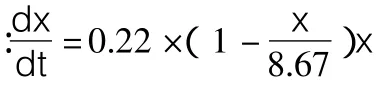
Armillaria luteo-virens Sacc;batch fermentation;kinetic model;fit
TS201.3
A
1002-0306(2010)11-0135-03
2009-10-30
余梅(1977-),女,生产部主管,研究方向:制药及微生物工程。
