滨海围垦地红叶椿光合特性研究
2010-09-04虞木奎王宗星朱春玲成向荣吴统贵
张 翠 ,虞木奎 ,王宗星 ,葛 乐 ,朱春玲 ,成向荣 ,吴统贵
(1.中国林业科学研究院亚热带林业研究所,浙江富阳311400;2.南京林业大学,江苏南京210037;3.上海市林业总站,上海 201300)
随着沿海防护林建设向纵深发展,生态-经济-休闲复合型防护林体系的构建在沿海发达地区日益受到重视。建立在滨海围垦地上的森林公园,既有良好的生态效益,又有很高的景观特色。上海南汇区滨海森林公园,最初为围垦的滩涂地,土壤水分含量高,粘性大,盐分高。从21世纪初开始,先后引进红叶椿(Ailanthus altissima cv.Hongye)、女贞(Ligustrum lucidum)、洋白蜡(Fraxinus pennsylvanica)等盐碱地适生树种,对改善当地生态环境和提升景观效益发挥了重要作用。
红叶椿是近几年培育成的一个臭椿品种,抗逆性强,适应性广,是优良的乔木型红叶观赏品种。目前,有关红叶椿的研究多集中在引种栽培方面[1-2],光合生理特性研究较少。笔者初步探究该树种在上海南汇区围垦滩涂地的适应性,旨在揭示红叶椿光合特性,为深入开展红叶椿的栽培、利用、抗盐碱性、碳氮循环等方面的研究提供实验依据。
1 材料与方法
1.1 试验地概况
研究区位于上海市南汇区书院镇滨海森林公园(北纬 30°58′,东经 121°55′),东临东海,南靠杭州湾,距海2~3 km,平均海拔2~3 m。年平均气温15.6℃,年平均降雨量1 137.1 mm,6~9月降水较多。年日照时数1 993.4 h,无霜期223 d,属北亚热带海洋性季风气候。红叶椿于2006年穴状种植,种植密度为2 m×2 m,平均胸径4.5 cm,平均树高4.2 m。
1.2 试验方法
在红叶椿林分内选择3棵健壮植株为样株,选中部侧枝的中上部健康成熟功能叶片3片,每片叶分别挂牌标记,测定时保持叶片自然生长角度不变,进行活体测量[3]。在天气晴朗的2009年7月下旬,采用Li-6 400便携式光合测定仪,从8:00~18:00,每隔2 h测定叶片光合参数。测定指标包括净光合速率、蒸腾速率、气孔导度、CO2浓度、光合有效辐射、空气相对湿度等。测量每个叶片时,待读数稳定后,读取5~6个数据,取平均值。利用Li-6 400自动lightcurve曲线测定功能进行光响应曲线的测绘,测绘时利用缓冲瓶控制CO2浓度360~400μmol/mol,利用红蓝光源LED设定光合光通量密度梯度。测定数据利用SPSS与EXCEL软件进行分析。
2 结果与分析
2.1 红叶椿净光合速率和蒸腾速率日变化
在全光照条件下,红叶椿净光合速率Pn和蒸腾速率Tr日变化均呈单峰曲线(图1)。日出后,随光照强度增加,气温不断升高,净光合速率Pn随之增加,中午时分,净光合速率达到最大值11.58μmol/m2·s。光照强度在11:00~14:00维持在当天较高水平,14:00以后随着光照强度降低,气温下降,净光合速率逐渐降低,17:00左右,光合作用与呼吸作用基本持平,日落后趋于负值。蒸腾速率的日变化规律与净光合速率的日变化规律基本一致,6:00至12:00蒸腾速率逐渐增强,12点以后蒸腾速率逐渐降低,下午18点时仅为1.13μmol/m2·s。
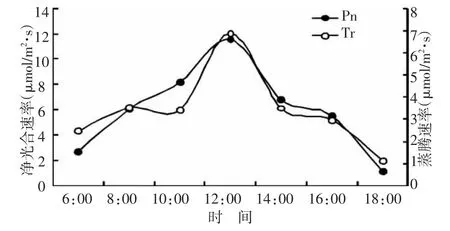
图1 红叶椿净光合速率和蒸腾速率的日变化
2.2 气孔导度和胞间二氧化碳浓度的日变化规律
由图2可以看出,气孔导度在早、晚比较低,随着光强逐渐升高,气温逐渐上升,空气相对湿度不断降低,叶片气孔开始扩张,气孔导度与蒸腾速率随之增高,在中午13:00出现一天中的最高峰,然后开始逐渐下降,到傍晚时分气孔基本关闭。气孔导度与蒸腾速率在日变化过程中呈同步变化趋势,具有极显著的正相关关系(r=0.665**,p<0.01)。胞间CO2浓度在早、晚比较高,随着光强逐渐升高、气温逐渐上升,胞间CO2浓度开始下降,结合图1,净光合速率低的时候,胞间CO2浓度高,且随着净光合速率的增加,胞间CO2浓度降低,二者具有极显著的负相关关系(r=-0.859**,p<0.01)。
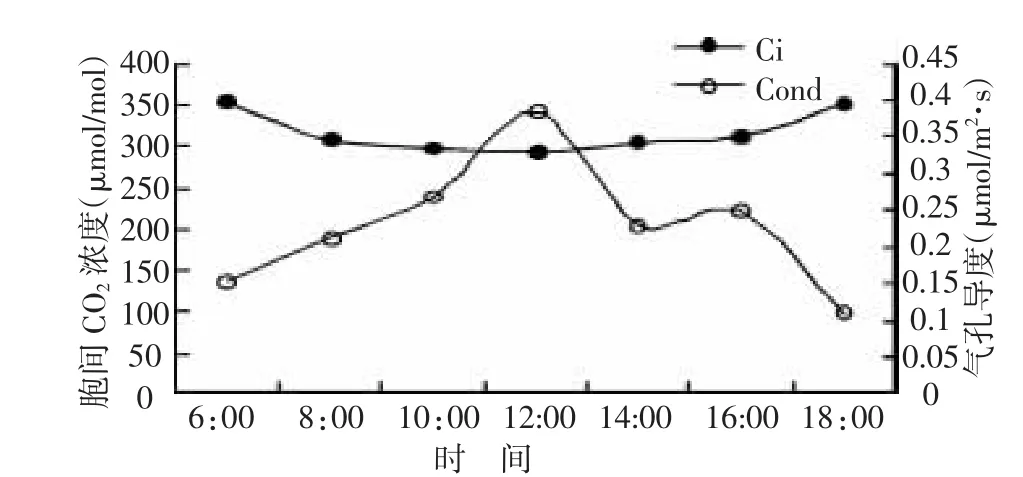
图2 气孔导度和胞间二氧化碳的日变化规律
2.3 光合有效辐射的日变化规律
由图3可以看出,日出后光合有效辐射不断增加,11:00~14:00点光合有效辐射稳定在较高的水平,近 13:00 时达到最大值 1 701 μmol/m2·s,下午光合有效辐射不断下降,至日落后光合有效辐射近于零,而此时的光合作用停止。此外,外界温度的变化规律与外界的光照辐射强度变化规律相似,而相对湿度在早晚较高,13:00出现最低值(55.4%)。
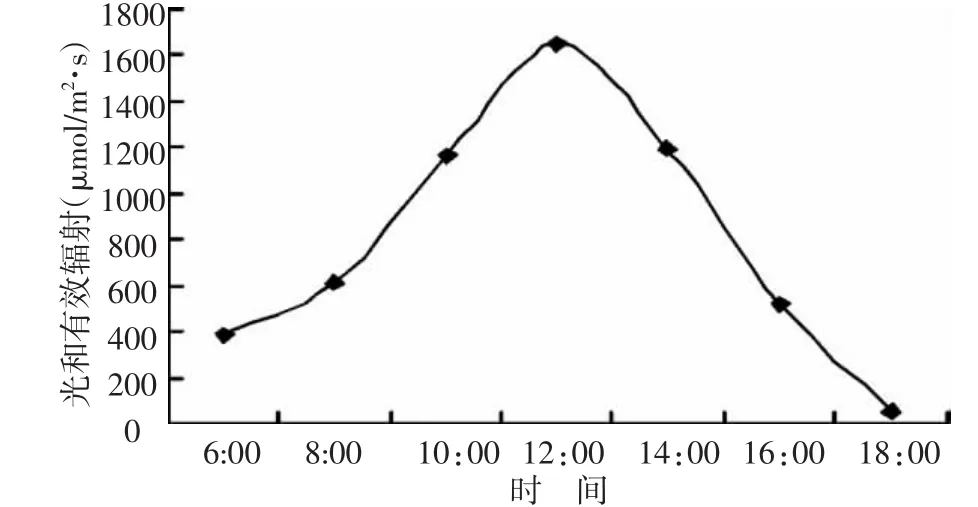
图3 光合有效辐射的日变化规律
2.4 环境和生理因子与光合特征参数之间的关系
植物的光合作用是植物生长快慢的重要影响因素,植物的光合作用和其他生命活动一样,经常受着外界环境因子的影响而不断发生变化[4]。如大气的温度、湿度、外界CO2浓度以及光合有效辐射等环境因子,以及由此而引起的净光合速率、蒸腾速率、胞间CO2浓度、气孔导度等生理生化因子。经相关性分析,净光合速率与光合有效辐射、蒸腾速率、气孔导度呈正相关关系,与胞间CO2浓度、大气相对湿度呈负相关关系。光合有效辐射与蒸腾速率呈极显著正相关关系,与胞间CO2浓度呈极显著负相关关系,与气孔导度呈正相关关系;大气CO2浓度与胞间CO2浓度呈显著负相关关系,与蒸腾速率和气孔导度呈负相关关系;大气温度与蒸腾速率呈极显著正相关关系,与胞间CO2浓度呈极显著负相关关系;大气湿度与蒸腾速率呈极显著正相关关系,与胞间CO2浓度呈极显著负相关关系;大气温度与大气湿度均与气孔导度呈正相关关系,但相关性不显著。环境因子对光合作用的影响主要是通过蒸腾速率来作用,外界大气CO2浓度的影响最小,光合有效辐射对蒸腾速率的变化起决定作用,进而影响到了净光合速率的变化(见表1)。
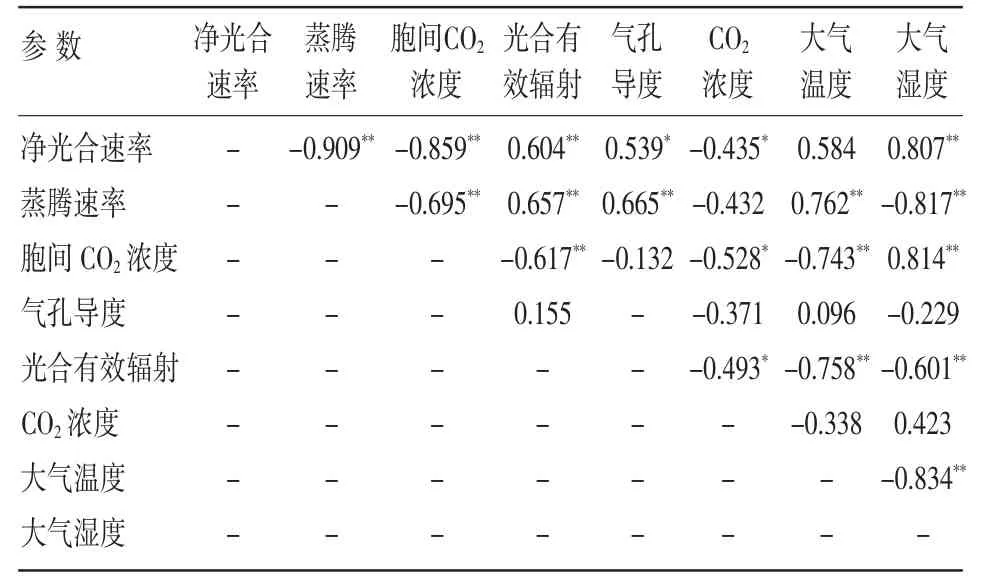
表1 红叶椿光合速率与环境因子、生理因子的相关系数
2.5 红叶椿光响应曲线
光响应曲线反应了植物净光合速率随光强度变化而变化的规律。光响应曲线采用非直角双曲线进行拟合[5],当PPFD<200时,对PPFD-A进行直线回归,光合速率随光强的增加而呈比例的增加,净光合速率(y)与光照强度(x)之间呈线性相关y=0.0 375 x-1.191 6。由线性方程可知,光补偿点为31.78μmol/m2·s,表观量子效率为0.037 5,暗呼吸速率为1.191 6μmol/m2·s。当光照强度变化范围在200~1 400μmol/m2·s时,光合速率随光照强度的增加而增加,光合速率从4.88μmol/m2·s增加到12.01 μmol/m2·s,光照强度为 1 400~2 000 μmol/m2·s时,光合速率不再增加,呈现光饱和状态,根据光响应曲线计算得出光饱和点为1 365.11μmol/m2·s。
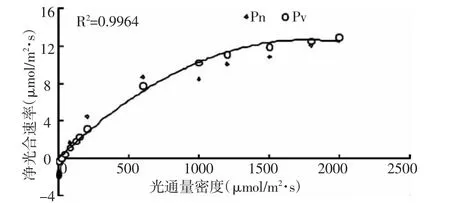
图4 红叶椿光响应曲线
3 小结与讨论
植物的光合作用是个复杂的生理过程,不仅取决于植物本身的遗传特性,同时还受到外界环境的影响和制约。红叶椿净光合速率为单峰曲线,未出现光合午休现象,气孔导度和光量子通量密度的日变化为单峰曲线,这与西藏巨柏,扁穗牛鞭草等植物的日变化规律相一致[6-16]。宋庆安等研究发现臭椿胞间CO2浓度随着气孔导度的降低而减少,臭椿的光合速率控制因素为气孔限制[7]。结合图1和图2可发现,净光合速率随着气孔导度的降低而降低,但胞间CO2浓度并没有随之降低。这说明光合速率受到非气孔因素的影响。植物光合作用的温度范围在28~36.5℃之间。在试验地7月的晴朗天气条件下,一天时间内,空气温度在此范围内变化不大,其变化对净光合速率的影响不及光合有效辐射明显。
已有的研究结果指出,强光抑制是引起光合午休的主要原因。由光响应曲线可知,红叶椿光补偿点低,光饱和点1 365.11μmol/m2·s,当天的最高光照辐射达到1 700μmol/m2·s,远大于光饱和点,但红叶椿并没有出现光合午休现象。可能由于环境因子的影响,植物的日变化规律发生了改变,植物在光强适度变化的条件下,Pn日变化曲线由双峰转变为单峰[8-9]。这与吴统贵对旱柳不同月份净光合速率日变化研究和张友炎对紫穗槐的净光合速率日变化研究结果一致[11-12]。研究者已对臭椿的生理生态特性开展了一系列的研究,发现臭椿的净光合速率与气孔导度均为双峰曲线,有明显的午休出现[10]。红叶椿作为臭椿的变种,光合日变化与臭椿明显不同,其原因还有待于进一步研究。
蒸腾速率的变化与红叶椿的光合速率最为密切相关,光合有效辐射对蒸腾速率的变化起决定作用,大气相对湿度是最直接的限制因子(表1)。这与绝大多数的木本植物相一致[13~15]。
综上所述,红叶椿的净光合速率呈单峰曲线。中午时分,净光合速率达到最大值11.58μmol/m2·s。胞间CO2浓度的变化趋势与净光合速率的变化趋势相反,光合速率受到非气孔因素的影响。红叶椿的表观量子效率是0.037,红叶椿光饱和点为1 365.11μmol/m2·s,光补偿点为 31.78 μmol/m2·s,为喜光树种,能忍受强光的照射,这与上海滨海森林公园的光照条件相适应。
[1]王鸣凤,陈柏林.红椿树的生物学特性及人工栽培研究[J].林业科技通讯,1999,(1):15-17.
[2]王勤华,曹春英,孙曰波,等.红叶椿的组织培养和快速繁殖[J].植物生理学通讯,2004,40(1):73-78.
[3]殷 工,徐 立.新疆沙漠桑树光合特性日变化的研究[J].中国农学通报,2008,24(8):257-259.
[4]王 忠.植物生理学[M].北京:中国农业出版社,1999.
[5]刘宇锋,萧浪涛.非直线双曲线模型在光合光响应曲线数据分析中的利用[J].农业基础科学,2005,21(8):76-79.
[6]兰小中,廖志华,王景升.西藏高原濒危植物西藏巨柏光合作用日进程[J].生态学报,2005,25(12):3172-3175.
[7]宋庆安,童方平,易霭琴.臭椿光合生理生态特性日变化研究[J].中国农学通报,2007,23(12):148-153.
[8]He J,Chee C W,“Photoinhition”of Heliconia under natural tropic condition:the inportance of leaf orientation for light interception and leaf temperatue[J].Plant,Cell and Enviroment,1996,19(11):1238-1248.
[9]Senevirathna A M,Stirling C M,Rodrigo V H L.Acclimation of photosynthesis and growth of banana(M usa sp.)to natural shade in the humid Tropics [J].Experimental Agriculture,2008,44(3):301-312.
[10]梁月,郭建斌,殷丽强.黄土高原侧柏、臭椿光合、蒸腾作用比较研究[J].水土保持研究,2008,15(2):156-158.
[11]张友焱,刘致远,周泽福,等.环境因子对毛乌素沙地紫穗槐光合、蒸腾速率的影响[J].林业科学研究,2006,19(4):463-466.
[12]吴统贵,周和锋,吴 明.旱柳光合作用动态及其与环境因子的关系[J].生态学杂志,2008,27(12):2056-2061.
[13]郭志华,张宏达,李志安,等.鹅掌楸苗期光合特性研究[J].生态学报,1999,19(2):164-169.
[14]邵玺文,韩 梅.不同生境条件下黄芩光合日变化与环境因子的关系[J].生态学报,2009,29(3):1420-1476.
[15]张 璐,张纪林.不同光照条件下3种冬青属植物的光合特征日变化研究[J].西北植物学报,2006,26(3):0490-0495.
[16]陈灵鹫,杨春华,傅鲜桃,等.扁穗牛鞭草光合特性研究[J].四川农业大学学报,2007,25(4):484-488.
