植物细胞内膜运输调控机制及相关前沿电子显微镜技术应用的研究进展
2022-05-20李彦斌郑晓慧徐一月
李彦斌,齐 青,郑晓慧,徐一月,崔 勇*
(1.厦门大学生命科学学院,细胞应激生物学国家重点实验室,福建 厦门 361102;2.香港中文大学生命科学学院,香港 999077)
与大多数真核细胞一样,植物细胞含有一套由核膜、细胞膜以及多种细胞器组成的内膜系统,包括内质网(ER)、高尔基体(Golgi)、反式高尔基体网络(TGN)、液泡前体/多囊泡体(PVC/MVB)、液泡和自噬体等.蛋白质等物质在植物细胞内经该内膜系统运输,完成细胞内物质及信息的传递和交换;同时,精确和高效的物质运输对生物体的正常生长发育和环境应答都十分重要,因此了解掌握植物蛋白在细胞内运输的调控机制显得尤为关键[1].
蛋白质在不同细胞器之间的运输主要经囊泡介导,而这些囊泡大部分直径都小于200 nm,用普通光学显微镜无法清晰观察,这令此领域研究者长期在一些关键问题上存在争议[2].而随着电子显微镜技术的发展,这些精细的物质运输过程逐渐得以呈现,不仅帮助厘清了此领域中一些长期存在的争议,还让研究者们发现了全新的运输模式,极大地推动了内膜运输在植物细胞研究中的进展.本文重点阐述当前植物蛋白运输途径的研究成果,结合近期研究展示前沿电子显微镜技术在植物细胞内膜运输研究中的应用,并对本领域研究存在的机遇与挑战进行展望.
1 植物细胞内囊泡介导的蛋白运输途径
在植物细胞内膜系统中,一方面,新合成的蛋白质需要先被运送到不同细胞器完成蛋白修饰后才能被运送到目标位置行使功能;另一方面,一些在细胞膜定位的膜蛋白在感知环境变化后会被运送到细胞内的特定部位完成信号的传递.细胞内众多通路彼此高效协作,共同调控蛋白质在细胞内的靶向运输,维持细胞内环境的稳态.下文主要介绍植物细胞内囊泡介导的最重要且常见的3条运输途径:蛋白分泌途径、靶向液泡的运输途径和内吞途径,图1所示为经典分泌途径和内吞途径.
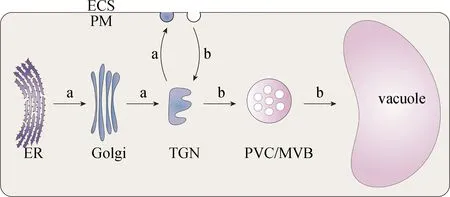
a.经典分泌途径:ER-Golgi-TGN-细胞膜(PM)/细胞外(ECS);b.内吞途径:ECS/PM-TGN-PVC/MVB-液泡.
1.1 植物蛋白分泌途径
在分泌途径中,新合成的蛋白质在完成加工之后会被运输到PM或ECS行使功能,对维持细胞内环境稳定、促进细胞间信息交流以及应答生物或非生物胁迫等有重要的生理意义.植物蛋白分泌途径主要分为经典分泌途径和非经典分泌途径.
1.1.1 经典分泌途径
在经典分泌途径中,对于多数分泌蛋白而言,其氨基端(N端)会含有一段疏水的氨基酸序列,称为信号肽(SP),当肽链在核糖体上开始合成后,SP序列会指引其进入ER腔内进行后续的翻译以及折叠修饰等[3-4].除经典的共翻译转运外,最近的研究还发现了一些翻译后转运的分泌蛋白.此类蛋白尾部含有一个膜锚定结构(TA),通过特定的TA蛋白指引复合物帮助其插入ER膜[5].无论是共翻译还是翻译后转运,肽段进入ER腔内都需要借助Sec转运体系.在酿酒酵母(Saccharomycescerevisiae)中Sec转运体系由形成转运通道的Sec61异源三聚体(Sec61p、Sbh1p和Sss1p)以及Sec62p、Sec63p、Sec71p、Sec72p组成的异源四聚体复合体所构成[6-7].Sec61异源三聚体和Sec63p参与共翻译和翻译后蛋白转位,而Sec62p、Sec71p和Sec72p仅与翻译后转运相关[8].在拟南芥(Arabidopsisthaliana)基因组中有3个基因编码Sec61p的同源蛋白,其中两个与Sec63p同源,另一个与Sec62同源,且最新研究表明sec62突变体表现出生长受阻、花粉败育[9],但具体机制仍有待阐明.
在ER完成加工后,包被蛋白(COPⅡ)囊泡会介导蛋白货物从ER传递至Golgi.这一过程最早在酵母中被发现,此后证明其在动物和植物中都是保守的[10].COPⅡ囊泡的形成首先是将胞质中小G蛋白Sar1招募到ER膜上,然后Sec12作为其特异的鸟苷酸交换因子(GEF)使得Sar1变成结合三磷酸鸟苷(GTP)的激活形式,随后激活态Sar1招募一个由GTPase激活蛋白Sec23和接头蛋白Sec24所组成的异源二聚体,形成一个出芽的复合体,其中Sec24可以结合可溶性货物的受体蛋白或跨膜的货物蛋白,从而捕获货物[11];此复合体又招募Sec13-Sec31异源四聚体,作为COPⅡ最外层的包被蛋白,最终形成完整的COPⅡ囊泡[12-13].通过生物信息学分析,COPⅡ囊泡的某些蛋白组分的表达水平还会随着各种胁迫条件和发育阶段的变化而改变,表明植物中COPⅡ囊泡的数量处于动态变化中[14].
货物蛋白被运输至Golgi后,大多数会通过TGN分泌至PM或ECS,但最近研究也发现存在由Golgi直接产生的分泌囊泡[15].研究分泌蛋白运输途径最直接的方式就是监测PM上新合成的膜蛋白运动,如利用免疫荧光技术观察细胞膜表面受体激酶(BRI1)蛋白的分布,发现在被运输至PM之前,BRI1还有TGN的定位,说明BRI1的分泌是由TGN衍生的囊泡所介导的[16].除分泌单个的囊泡外,研究还发现TGN衍生的分泌囊泡能聚集成束,形成由5~12个囊泡组成的不规则大囊泡,直径为50~100 nm,极大地提高了运输效率,如分泌载体相关膜蛋白2(SCAMP2)就是通过此种途径运输的[17].
1.1.2 非经典分泌途径
近些年来,越来越多的研究表明很多分泌型蛋白并不含SP序列,其分泌是通过非经典分泌途径完成的.前期在动物和酵母的研究中就已经证明了非经典分泌途径的存在,如动物中成纤维细胞生长因子2(FGF2)蛋白和白介素-1α(IL-1α)蛋白都是在疏水性释放复合物的协助下,通过跨膜通道直接穿过PM[18-19].
在植物中,非经典分泌途径也被慢慢发现.布雷菲德菌素A(BFA)是一种特异性阻断蛋白质从ER转运至Golgi的大环内酯类抗生素,且用BFA处理后的细胞会产生Golgi衍生囊泡聚集体,称为“BFA体”[20],因此BFA常被用于非经典分泌途径的研究.HYGR是一种缺乏SP的植物胞质蛋白,可以被分泌到ECS,且BFA处理后并不会影响该分泌过程[21];此外,甘露糖脱氢酶在水杨酸诱导下的分泌也表现出BFA不敏感性,表明非经典分泌途径在植物应答胁迫过程中也发挥作用[22].然而在这两种蛋白的运输过程中,并未鉴定到任何囊泡状结构,暗示着植物可能存在非囊泡介导的Golgi旁路分泌途径.事实上,在动物细胞中确实发现PM上孔状结构的形成有利于蛋白的直接整合,但这一结构在植物中还未被发现[23].
此外,非经典分泌途径也介导了植物胞外囊泡(EV)的形成.EV这一概念在2011年才被正式提出,用来定义细胞向外分泌的各种具有脂质层包被的囊泡,如外泌体、凋亡小体和微泡等[24].最初EV被认为是细胞向外排出代谢废物的一种途径,但最近研究发现哺乳动物细胞分泌的外泌体还包裹着抗原物质和信使RNA(mRNA)[25-26],可以促进细胞间信息交流,具有很高的临床应用价值,对外泌体等EV的研究在生物学界掀起了热潮.早在20世纪60年代,研究者们用透射电镜观察胡萝卜(Daucuscarota)细胞时就发现了植物中的EV[27],但受限于当时的分离纯化技术,EV在植物中的研究远滞后于动物中的研究.直至2017年,研究者们利用优化的差速离心法结合碘克沙醇密度梯度离心,才首次在植物叶片中分离出纯度较高的EV[28].进一步通过生化分析鉴定出植物EV中含有核酸、蛋白质、脂质等活性物质,而这其中就包括很多参与细胞壁重塑和降解的酶[29],以及可以沉默病原菌基因表达的小分子RNA[30].上述证据说明,EV对植物正常生理活动的进行以及防御病原菌等方面都具有重要意义.植物中的EV主要包括exocyst亚基EXO70E2标记的EXPO(exocyst positive organelle)、四跨膜蛋白8(TET8)标记的外泌体和PEN1(plant syntaxin)标记的EV.
EXPO是一种新型的双层膜包被的细胞器,膜上含有特殊的exocyst复合体,可以介导分泌囊泡与质膜的拴系和融合.在对拟南芥EXO70E2蛋白进行亚细胞定位研究时,研究者们发现其定位既不受BFA、ConA、wortmannin等经典分泌途径抑制剂的影响,也不与自噬体、Golgi、TGN、PVC/MVB和液泡膜标记蛋白共定位[31].利用免疫电镜技术,观察到其标记的一种双层膜结构与PM融合后可以释放单层内膜包裹的囊泡(直径为200~500 nm),使胞外也能检测到EXO70E2蛋白,由此将此种新发现的双层膜细胞器称为EXPO[32].S-腺苷甲硫氨酸合成酶2是一种维持细胞壁结构的木质素生物合成酶,正是通过此种途径分泌的[33].
对动物中分离出来的外泌体进行分析时,研究者们发现其膜上有很多TET,如CD9、CD37、CD63等[34],这些TET可以用作动物外泌体的标记物.拟南芥中有17个TET家族成员,通过序列比对和表达分析发现TET8与动物中CD家族同源性最高[35].且通过荧光共定位发现TET8与植物中Rab5蛋白(ARA6和RHA1)在胞内共定位[36],即TET8在胞内定位于MVB上.结合TET8有PM和ECS的定位,可以认为TET8是植物外泌体特有的一个标记物.当用灰霉菌(Botrytiscinerea)侵染植物时,TET8的表达丰度还会显著提高,暗示植物分泌外泌体很有可能也是抵御病原菌入侵的一种防御手段.事实证明这种TET8标记的EV中含有的小分子RNA可以有效沉默病原菌特定基因的表达,从而达到防御目的[30].
PEN1是一种具有病原菌渗透抗性的蛋白,在拟南芥中亦称为SYP121,属于PM定位的Qa-可溶性N-乙基马来酰亚胺敏感性因子附着蛋白受体Qa-SNARE;其分布在PM和ECS,但并不与Rab5共定位,也不与TET8共定位,由此认为PEN1标记的EV不同于TET8标记的EV[36].且在分离纯化时,这两种EV位于不同的密度梯度组分中,验证了二者的差异[28].
此外,液泡也可以介导非经典分泌途径的发生.植物细胞的液泡有较低的pH值且含各种水解酶,其功能类似于动物细胞中的溶酶体,是细胞中物质降解的中心.通常细胞中需要被降解的蛋白、代谢废物等都会被运送到液泡降解,但植物在抵御病原菌入侵时,也需要这些水解酶或是其他具有抗性的次生代谢产物,此时就需要液泡再向外分泌这些物质.当用病原菌Pst(Pseudomonassyringaepathovartomato)菌株DC3000侵染拟南芥时,在电镜下可以观察到液泡膜与PM的融合过程,同时此过程需要病原菌的抗原蛋白AvrRpm1与植物抗斑生假单胞菌(P.maculicola)蛋白RPM1的互作[37].关于植物如何触发液泡与PM的融合以及具体的膜融合机制仍有待进一步研究.
1.2 植物蛋白靶向液泡的运输途径
1.2.1 Rab5和Rab7双依赖型靶向液泡的运输途径
在经典的蛋白分泌途径中,新合成的蛋白会从ER运输至Golgi,再进入TGN进行分选,对于含有液泡分选序列的蛋白来说,它们会靶向液泡[38].液泡货物蛋白在TGN上被特有的液泡分选受体(VSR)所识别并结合,VSR与货物蛋白结合后并非独立地将蛋白运输至液泡,而是以网格蛋白包被囊泡(CCV)的形式不断与中间过程的细胞器融合,最终到达液泡.因此对VSR的鉴定,最初是通过纯化CCV来实现的[39].现已证明接头蛋白1(AP-1)/AP-4复合体参与CCV的形成[40].当货物被运输至MVB后,MVB进而与液泡融合.在拟南芥中,MVB也有两种状态,分别由Rab5和Rab7标记.通过寻找动物中的同源蛋白,发现莫能菌素敏感性1(MON1)/钙咖啡因锌敏感性1(CCZ1)复合体可以与GTP结合的Rab5互作,亦可与二磷酸鸟苷(GDP)结合的Rab7互作,从而揭示了植物中Rab5与Rab7转换的分子机制[41].此条途径可以简单概括为:Rab5首先招募C类核心液泡内体束缚(CORVET),亦可招募MON1/CCZ1作为Rab7的GEF,从而激活Rab7,激活态的Rab7又可以招募同型融合和蛋白质分选复合体(HOPS)等效应蛋白,最终介导MVB与液泡的融合(图2-a).拟南芥中运输至蛋白储存型液泡的12S球蛋白,以及运输至溶解型液泡的液泡硫基蛋白酶(aleurain),均是通过此种方式运输的.
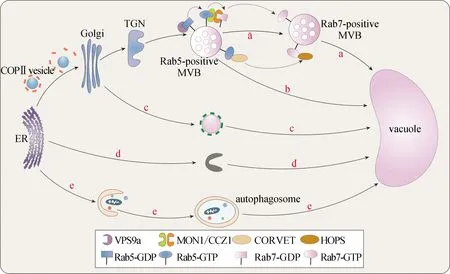
a.Rab5和Rab7依赖途径,Rab5招募MON1/CCZ1复合体激活Rab7,最终由Rab7介导MVB与液泡的融合;b.Rab5依赖途径;c.AP-3复合体依赖途径;d.从ER直接运输到液泡途径;e.自噬途径.
1.2.2 仅依赖Rab5靶向液泡的运输途径
拟南芥中共有3个编码Rab5的同源蛋白ARA7、RHA1和ARA6.其中ARA7和RHA1负责介导蛋白靶向液泡的运输;而ARA6为植物特有,负责介导MVB向PM的运输[42].尽管它们发挥不同的功能,但都有一个共同的GEF(VPS9a),可以使得Rab5以GTP结合的方式锚定在MVB膜上[43].最近的研究发现VPS9a的羧基端(C端)结构域可以与ARA6互作,但并不与ARA7或RHA1互作,暗示此区域的互作位点可用于区分不同的Rab5[44].vps9a突变体表现出生长缺陷,也证明Rab5在植物液泡运输中发挥不可或缺的作用[45].虽然多数液泡蛋白通过经典途径靶向液泡,但是也存在只依赖于Rab5的蛋白靶向液泡运输途径(图2-b).SYP22是一个靶向液泡膜的SNARE蛋白,在vps9a-2突变体中SYP22的运输受到抑制,但在mon1-2突变体中却表现正常,证明SYP22的运输不依赖Rab7而只依赖Rab5[46].
1.2.3 依赖AP-3复合体靶向液泡的运输途径
AP-3复合体介导的液泡运输最初在酵母中发现,被鉴定为碱性磷酸酶的运输途径[47],后续在动物中也被证明介导溶酶体的靶向运输[48],且均无需再经过中间细胞器,通过Golgi后可直接将货物运输至液泡或溶酶体[49].AP-3复合体是一个异源四聚体,由两个大亚基(δ和β3)、一个中亚基(μ3)以及一个小亚基(σ3)构成[50].在拟南芥中,跨膜蛋白PAT10已被证明是AP-3复合体介导的蛋白靶向液泡运输途径中的货物[51](图2-c).在ap-3β突变体中,发现一种靶向液泡的货物蛋白SUC4被滞留在Golgi而非TGN中.
1.2.4 ER直接靶向液泡的运输途径
上述几种植物蛋白靶向液泡的运输途径都需要经过Golgi.而在2013年,有研究者利用遗传学结合光学和电子显微镜成像等多种技术,在拟南芥根分生组织细胞中发现H+三磷酸腺苷(ATP)酶VHA-a3可以从ER直接靶向液泡[52](图2-d),但具体的分子机制尚不明确.
1.2.5 自噬途径
自噬在真核生物中高度保守,是受损的蛋白质和细胞器循环再利用的重要途径,在植物细胞中通过自噬体与液泡融合完成,是靶向液泡运输的一种类型,对植物维持细胞稳态和响应各种环境胁迫(如营养缺陷、逆境、病原体感染等)必不可少(图2-e).自噬由一系列核心的自噬相关基因(ATG)蛋白起始和调控(图3).ATG蛋白根据其功能可分为4组:ATG1/ATG13激酶复合体、磷脂酰肌醇-3-激酶(PI3K)复合体、ATG9包含囊泡、ATG8/ATG12类泛素化结合系统[53].ATG1/ATG13激酶复合体负责响应细胞营养信号以起始自噬体的形成;PI3K复合体在自噬体膜上催化产生磷脂酰肌醇-3-磷酸(PI3P),通过招募不同的PtdIns3P结合蛋白来调控自噬过程,包含VPS34、VPS15、ATG6、ATG14共4个亚基;ATG9包含囊泡穿梭于不同膜来源的细胞器,运输膜脂以促进自噬泡的扩张;ATG8/ATG12类泛素化结合系统促进自噬体的扩张和成熟.首先ATG5与ATG12共价连接以形成复合体,然后该复合体进一步与ATG16互作以产生ATG5-ATG12/ATG16多聚体.该多聚体作为E3连接酶发挥作用,将ATG8连接到自噬体的磷脂酰乙醇胺(PE)上.

植物自噬体膜来源众多,包括ER、ATG9囊泡和COPⅡ囊泡等.这些膜结构首先形成前自噬体结构(PAS),然后在需要降解的货物(如线粒体、叶绿体)周围形成杯状双层的吞噬体膜;接着吞噬体膜伸长并密封形成自噬体;最后自噬体与液泡融合,其内容物被液泡水解酶分解,完成自噬过程.
自噬的诱导受细胞营养状态的调节.在营养丰富的条件下,雷帕霉素靶蛋白(TOR)和其他激酶诱导ATG13亚基过磷酸化和ATG1激酶低磷酸化,促使ATG1/ATG13激酶复合体解离.最近研究表明,拟南芥肿瘤坏死因子受体相关因子(TRAF)家族蛋白中的TRAF1a和TRAF1b可作为支架蛋白,与RING E3泛素蛋白连接酶SINAT1、SINAT2互作,通过泛素化途径降解ATG13或ATG6,将自噬维持在适当水平[54-55].然而,在饥饿等营养缺乏时,蔗糖非发酵相关蛋白激酶1(SnRK1)会抑制TOR活性,导致ATG13去磷酸化以及ATG1/ATG13激酶复合体的结合以启动自噬.
自噬体的膜起源是植物自噬领域长期研究的问题.近年研究发现,拟南芥中ATG9的缺失可导致在自噬过程中扩大的自噬体相关小管连接到ER,为ER作为植物自噬体的膜来源提供了结构学证据[56].此外,COPⅡ囊泡不但介导蛋白质从ER到Golgi的正向运输,还可在自噬过程中作为自噬体生物发生的膜来源之一[57](图3).
一些非ATG蛋白也参与自噬过程,如乳腺癌 1号基因邻近基因1(NBR1)、SH3结构域蛋白2(SH3P2)等.NBR1和ATG8-PE互作,作为选择性自噬受体在蛋白质聚集体的清除中发挥重要功能[58].SH3P2是一种膜相关蛋白,与拟南芥中PI3K复合体结合并与ATG8互作,协同促进膜扩张或成熟[59].最新研究表明自噬受体蛋白NBR1的数量增减会影响ATG8与SH3P2的互作[59],细胞以此来调节选择性自噬途径的活跃程度,即按需要将自噬货物运送至植物液泡降解,维持营养和能量的平衡.
自噬体在完成货物装载后会与液泡融合,其内容物被液泡水解酶分解,完成自噬过程(图3).然而在植物细胞中,自噬体膜的闭合过程以及与液泡的融合机制还不是很清楚.通常认为多种Rab蛋白、拴系复合物和SNARE蛋白等都可能参与自噬体与液泡的融合.
1.3 植物内吞途径
与分泌途径相对,内吞作用也是真核生物维持胞内环境稳定、应答外界环境信号的重要方式.相比于动物细胞,植物细胞特有的细胞壁和液泡结构使其多了一道天然屏障,并大大提高了膨压,但同时在一定程度上阻碍了PM内陷.植物细胞是否会发生内吞作用曾引发学界的激烈讨论.
深入的研究发现植物细胞不仅会发生内吞作用而且内吞作用还具有重要的生理意义.在植物细胞中,根据货物蛋白在PM上凹陷出芽形成的小泡类型,将内吞途径分为网格蛋白介导的内吞(CME)途径和非网格蛋白介导的内吞(CIE)途径.
1.3.1 CME途径
CME是植物中主要的内吞途径,也是研究得较为清楚的途径.生长素极性运输载体PIN2、水通道蛋白PIP、铁转运蛋白IRT1等均通过此种内吞途径进入细胞[60].CME发生的过程包括以下5个步骤.1)内吞货物的募集:此步骤需要AP复合体.在真核生物中共鉴定到5种AP复合体,其中AP-2复合体在内吞途径中发挥重要作用.AP-2复合体也是一个异源四聚体,由两个大亚基(α和β2)、一个中亚基(μ2)和一个小亚基(σ2)构成.其中AP-α可以与PM上二磷酸磷脂酰肌醇或三磷酸磷脂酰肌醇结合稳定内吞装置,AP-μ2可以特异识别内吞货物将其分选募集[61].2)网格蛋白的招募:AP-2复合体的作用类似桥梁,一端连接着内吞货物,另一端则通过AP-β2招募网格蛋白,并在PM形成内陷的小窝(CCP).此过程还需要其他辅助蛋白以促进网格蛋白的招募和膜内陷,如Epsin1和AP180[62]等.3)CCP从膜上缢裂形成CCV.缢裂的过程需要膜缢裂相关蛋白(DRP),这是一类大GTPase蛋白家族,通过缠绕在CCP茎上水解GTP,释放能量以促进CCP从膜上脱落[63].DRP的招募由含有Bin/两性蛋白/Rvs(BAR)结构域的endophilin和SNX9蛋白完成,在拟南芥中DRP1和DRP2表现出对内吞过程的调控[64].4)CCV 脱去包被蛋白:auxilin、热激同源蛋白70(HSC70)等蛋白会破坏AP-2与网格蛋白间的连接[65],从而促进网格蛋白的释放.5)最终内吞囊泡与内体融合,完成货物运输.
内吞作用很大程度上会影响PM表面的分子分布,进而触发信号转导等过程,因此也会受到生物或非生物因子的调控.最新的一项研究发现,三价稀土元素镧[La(Ⅲ)]可以调控CME:用低剂量La(Ⅲ)处理叶片后,可以激活叶细胞的CME过程,且还会将此信号长距离传导至根;而这种长距离运输的机制是La(Ⅲ)激活叶细胞CME后,触发AtrbohD(一种产生活性氧的还原型辅酶Ⅱ氧化酶)和茉莉酸的协同作用,使信号得以传递[66].
1.3.2 CIE途径
在动物细胞中,CIE途径主要包括脂筏蛋白介导的内吞、巨胞饮以及非网格蛋白、脂筏蛋白依赖的胞吞等,例如IL-2受体就是由endophilin所介导的CIE途径进行内吞的货物.研究表明endophilin通过小G蛋白Cdc4招募Formin结合蛋白17(FBP17)和Cdc42相互作用蛋白4(CIP4)蛋白在PM上富集,FBP17和CIP4又招募磷酸酶SHIP2和lamellipodin,使得PM上局部产生二磷酸磷脂酰肌醇,最终endophilin被富集在PM上特定区域引发内吞作用[67].最近关于植物CIE的研究日益增多,如有研究证明烟草BY-2(NicotianatabacumL.cv.Bright Yellow 2)细胞被CME途径的抑制剂斑鸠霉素(ikarugamycin)处理后仍能吸收外界环境中的葡萄糖[68],暗示植物体内也存在CIE途径.此外,通过对拟南芥质膜内在蛋白2;1(PIP2;1)的研究发现,在不同状态下该蛋白可以依赖不同途径进行内吞作用[69]:在正常情况下,细胞通过CME途径进行PIP2;1蛋白的内吞作用;而在高渗胁迫时,脂筏介导的内吞途径被激活,共同参与该蛋白的内吞过程.进一步研究发现,PIP2;1蛋白的内吞过程部分与脂筏标记蛋白1(Flot1)介导的PM微区内吞有关,抑制Flot1的表达会对植物的生长发育造成影响.另外呼吸爆发氧化酶同系物D(RbohD)蛋白的内吞作用也同时受网格蛋白和PM微区的调控,特别是受到盐胁迫时,由Flot1富集的PM微区内吞作用变强[70].除了葡萄糖、蛋白质可以经CIE途径进入植物细胞外,植物激素如油菜素内酯(BR)也被CIE途径调控.施加BR可以促进其受体BRI1进入Flot1富集的PM微区并由其介导内吞[71].由此可见,当植物经受外源刺激时,CIE在植物体内发挥重要调控作用[72-73].
2 前沿电子显微镜技术在植物内膜系统研究中的应用
在过去的20年中,研究者们借助于显微镜技术直接明确地观察到植物内膜系统中的囊泡运输过程,并解析出不同蛋白在其中发挥的作用,极大地加快、加深了人们对植物内膜系统精准调控的理解.激光共聚焦扫描显微镜作为一种高端的光学显微镜,被广泛应用于观察细胞内囊泡的实时动态运输过程.相较于传统光学显微镜,电子显微镜的分辨率达到0.2 nm左右,可以提供分子甚至原子水平的信息.特别是随着科技的发展,越来越多前沿电子显微镜技术在提高观察分辨率的同时,也最大限度地保留了样品真实的生理状态.这些电子显微镜技术的使用令人们逐步看清了这些看似沧海一粟的荧光点的庐山真面目,极大地推动了本领域的研究进展,为更深入地理解分子机制提供了不可估量的帮助.根据研究对象和目的,下文将具体介绍在植物内膜系统研究中逐渐开始应用的4种前沿电子显微镜技术,分别为用于解析蛋白结构的冷冻电子显微镜技术(cryo-EM)、着重进行细胞器三维重构的电子断层扫描技术(ET)、用来减薄冷冻样品的冷冻聚焦离子束技术(cryo-FIB),以及将光学显微镜与电子显微镜联用的光电联用技术(CLEM).
2.1 cryo-EM
目前解析蛋白结构的方法主要有X射线晶体衍射、核磁共振(NMR)和cryo-EM 3种.X射线晶体衍射是解析蛋白质大分子和晶体结构的主流手段,NMR技术则在小分子量的蛋白质结构解析中发挥重要作用,然而这两种技术都有一定的局限性.当蛋白质无法形成高质量晶体且分子量较大时,就需要用cryo-EM来解析蛋白结构,其主要原理是利用透射电镜在低温下观察包埋于玻璃态冰中的样品,然后通过软件对数据进行处理,从而得到样品的精细结构.1975年,Henderson等[74]首次利用cryo-EM和电子晶体学解析出细菌视紫红质膜蛋白的结构,开启了cryo-EM在结构学中的应用;Lepault等[75]于1983年发明了将样品速冻于玻璃态冰中的方法,使cryo-EM变得更为实用.自此,cryo-EM成为揭示蛋白结构的一个强有力工具.
cryo-EM最初主要用于结构生物学,随着技术的不断进步和革新,如今已广泛应用于微生物学、神经生物学、免疫学、细胞生物学等多个学科研究领域[76-78].在植物学研究中,应用cryo-EM在亚纳米水平解析了拟南芥自噬蛋白中唯一跨膜蛋白ATG9的三维结构,发现ATG9组成一个同源三聚体,每个单体至少有6个跨膜的α-螺旋,在三聚体的中心,相邻单体通过C端细胞质区域和膜嵌入区域的互作形成三聚体界面;结合使用序列共同进化信息对蛋白质结构的预测,该结构提供了ATG9分子水平的解析,并为ATG9调节自噬进展的分子机制提供了可检验的假设[79].类似地,研究者们通过cryo-EM方法解析出玉米(Zeamays)花粉中F-肌动蛋白的结构,显示其为一个右旋、双链(两条平行链)和交错的结构,该结构通过链内和链间的互作得以稳定,这对理解肌动蛋白如何作为长距离囊泡和细胞器运输的轨道至关重要[80].
对于体积和厚度较小的样品(如上述蛋白),利用直插式冷冻固定,对包被在冰膜中分散的、取向随机的成千上万的同一种蛋白分子进行拍照,将获得的蛋白照片进行叠加,即可获得这个蛋白接近于原子尺度分辨率的三维结构,该过程称作单颗粒分析(SPA).随着直接探测电子相机的出现和新算法的开发,SPA可达到原子分辨率水平[81].而当研究对象不具备周期性或全同性,如生物大分子复合体或细胞器、细胞骨架、囊泡运输系统时,样品不仅在体积和厚度上大过蛋白质且异质性高,直插式冷冻无法固定样品,且无法用SPA方式对图像进行叠加,研究者们于是开始寻求另一套冷冻固定、电子显微镜成像技术和三维重构的算法.
2.2 ET
ET是对样品中同一物体沿着垂直轴通过不同的倾斜角度进行拍摄,获得一系列二维投影图像后再重构其三维结构的方法.一般倾斜角度在-60°~+60°范围内,每1°~2°拍摄一次.根据样品的特性,ET又细分为树脂包埋样品的常温ET和非树脂包埋的冷冻ET(cryo-ET),常温ET的应用较为广泛,cryo-ET的应用则是近期的热点,尤其在植物中的应用才刚开始.通常为了最大限度地保存细胞器结构,在固定植物细胞时,相对于传统的化学固定,优选采用高压冷冻法(HPF)和冷冻替代法(FS)来制备样品.使用此方法固定的植物材料已被证明可以很好地应用于后续ET[82].目前在植物内膜系统研究中主要为常温ET,应用于细胞器起源以及新型细胞器的鉴定.在对拟南芥自噬体的研究中,研究者利用ET首次揭示了自噬体小管和ER膜之间的直接联系,为植物自噬体起源于ER膜提供了关键证据[83].而同样利用ET,在重构出的植物根细胞三维图像中,研究证实植物细胞中存在多个互相独立的液泡,并首次鉴别出一个直径400~1 000 nm的小液泡(SV),其中包含很多腔内囊泡;结合统计分析及对MVB形成或融合缺陷突变体的分析,最终证明植物中央大液泡是通过MVB融合形成SV,SV再进一步融合而来[79].由此解答了植物液泡的起源之谜,揭示了拟南芥根细胞中液泡形成的过程,结束了40多年关于液泡起源的争论[84-86].除此之外,研究者还利用ET鉴定出苜蓿(Medicagosativa)根缘细胞中一种源于TGN独特的介导半乳糖醛酸分泌的囊泡[87].在酵母和哺乳动物细胞等其他真核生物系统中,ET还成功应用于膜接触位点的研究,帮助阐明不同细胞器之间的连接机制[88-89].
为将样品更好地保持在其生理状态,cryo-ET提供了解析更接近自然状态的细胞器的方法.在样品制备方面,根据样品的厚度可以采用直插式冷冻法(样品厚度小于10 μm)或HPF(样品厚度小于300 μm).由于电子的穿透能力有限,通常要对样品进行减薄处理,虽然可以通过冷冻超薄切片完成,但是这些冷冻切片常出现因切割导致的压缩、折皱等假象[90],结果并不稳定且对操作者技术能力要求高,工序费时.于是研究者们将目光转向了材料学中常见的双束聚焦离子束(FIB)减薄法,尝试将其应用于ET的样品制备,并拓展至冷冻条件下的cryo-ET的样品减薄.另外,目前针对cryo-ET图像处理的子断层图像平均(STA)技术也正在悄然崛起.虽然目前其最终图像分辨率(0.3 nm)不及上述的SPA[91],但是该技术可以对较大、较厚、缺乏重复性的生物样品在其原位进行研究,这些优势都是SPA不可比拟的,有望成为日后研究细胞结构与功能的重要技术手段[76-78].
2.3 cryo-FIB
FIB技术是利用静电透镜将高能离子束聚焦成非常小的束斑,对材料表面进行轰击,从而达到材料的剥离、沉积和改性的一种方法,主要应用于材料学领域[92].目前FIB通常配置在扫描电镜(SEM)上,使加工过程可以实时观察,令操作更加精准.但FIB-SEM设备速度较慢,运行的时间有限,如果收集大体积样品,中途停顿会导致图像堆栈缺乏连续性.针对较大体积的生物样品,为了使FIB-SEM在数月的连续运行中变得更快、更可靠,研究者们改进FIB-SEM信号的检测方式,在合理的时间范围内对完整的果蝇(Drosophilamelanogaster)大脑进行成像,并使堆栈中各图像的间隙仅为8 nm[93].此外,高能离子束对样品具有很高的破坏性[94],但cryo-FIB技术的发展减少了离子束对样品的损伤[95-96].
在植物内膜系统研究中,FIB-SEM已应用于本氏烟草(Nicotianabenthamiana)叶细胞和大豆(Glycinemax)根瘤细胞的细胞器观察.在烟草叶片细胞中,研究者观察到紧密堆积的细胞器可能形成膜接触[97].利用ET重构出的大豆根瘤细胞的三维模型表明,与二维电子显微镜超薄切片显示的许多单独的共生体不同,受感染细胞中的类菌体全部包含在一个大的膜结构中[97].虽然cryo-FIB已在线虫、酵母、细菌、哺乳动物细胞、小鼠脑组织中有所应用[98-100],但是在植物领域条件仍未优化,其应用还鲜有报道.
2.4 CLEM
借助高分辨率重构的三维植物细胞模型,可以看到植物细胞内环境复杂且拥挤,内膜系统包含的细胞器众多[84].如上所述,为了更好地分析各细胞器,研究者们通常利用多种荧光蛋白或染料对数个细胞器或细胞结构进行标记,在光学显微镜下进行成像观察.然而受限于荧光显微镜约200 nm的分辨率限制,标记部分的超微结构无法看清.随着科技的发展,现在已经可以将观察得到的光学显微镜信号(通常是荧光)和EM图像相结合,从而获得感兴趣区域的高分辨率结构信息,并把这两种技术结合的方法称为CLEM.在CLEM发展早期,由于光学显微镜分辨率较低,光学显微镜与EM关联的准确性并不非常高.随着超高分辨率光学显微镜(SR-LM)的发展,也产生了如基于单分子成像技术的光激活定位显微术与常温透射EM结合[101]或与SEM结合的应用[102].这些工作预示着CLEM已进入SR-CLEM的发展阶段.随后,研究者们又成功地推动常温CLEM向冷冻方向发展,利用cryo-CLEM/cryo-ET观察了黄色黏球菌(Myxococcusxanthus)的第六分泌系统的结构[103]以及线粒体膜上转位酶TOM20的精细定位分布[104].与常温CLEM相比,cryo-CLEM的荧光强度大幅提高,且冷冻样品更接近细胞的自然状态.目前在冷冻条件下应用CLEM可帮助ET定位,大部分都应用于动物细胞和细菌[105].在植物中,常温下应用此技术的研究主要集中在TGN[106],而冷冻条件下CLEM和cryo-ET结合应用的条件仍需优化,一旦成功,其应用潜力将会非常大.
3 总结与展望
内膜系统是细胞中结构和功能的重要组成部分,负责将生物大分子等选择性运送到目标部位并发挥功能.植物蛋白运输主要通过囊泡介导,主要运输途径包括蛋白分泌途径、靶向液泡的运输途径和内吞途径,而细胞器则是这些运输途径的中转站并发挥多种功能.囊泡运输的调控和细胞器的生物发生一直以来都是细胞生物学研究的热点和难点.植物细胞含有不同于动物细胞的细胞壁,这对研究者在植物细胞中开展研究提出了更多的挑战.尽管越来越多的分子、新的运输途径甚至未知的细胞器被鉴定出来,人们对植物细胞内膜系统运输调控机制的理解仍处于初级阶段,对这些井然有序过程的调控机制还知之甚少,仍有大量重要的问题等待回答.例如:植物细胞内不同运输途径之间是否存在交叉互作?如果有,哪些蛋白介导了这些互作?互作的生物学意义是什么?当植物受到生物或非生物胁迫时,植物细胞内不同运输途径又是如何协调运作使植物在逆境中进行抵抗最终得以生存?能否利用基因工程的手段通过优化各蛋白运输途径以达到增加粮食产量、提高品质的目的?想要回答上述问题,多学科交叉(如与生物物理、计算机等学科)和新技术发展(如前沿电子显微镜成像技术)都是不可或缺的.在实验条件优化和自动化大数据高分辨率图片处理的配合下,有望逐步实现对植物细胞内囊泡运输、细胞器生成等生理过程的原位分析,在分子甚至原子层面阐明其调控机制,将结构与功能有机结合,进一步加深对本领域的认识;同时为内膜系统在动物细胞、真菌等不同系统的运作机制提供线索,最终实现在农业、医疗等领域的应用,助力可持续发展.
