辽东栎幼苗生长对种子大小和子叶去除处理的响应
2021-10-16朱雅婷王建礼闫兴富张金峰
朱雅婷 王建礼 闫兴富 张金峰
摘 要:該文在温室内盆播条件下,设置不去除子叶(对照)、去除1/4子叶(轻度子叶去除)、1/2子叶(中度子叶去除)和3/4子叶(重度子叶去除)4个处理,研究了种子大小和子叶去除处理对辽东栎幼苗生长及其形态学可塑性的影响。结果表明:(1)大种子幼苗株高、基茎、叶片数、单株叶面积、总干质量和根冠比在所有处理均大于小种子幼苗,除株高外,上述其他参数在轻度子叶去除处理的不同大小种子幼苗间均差异显著(P<0.05),前者总干质量在中度子叶去除处理与后者差异也显著(P<0.05)。大种子幼苗比叶面积和比根长在各处理均大于小种子幼苗,比枝长则相反。(2)随着子叶去除强度增大,大、小种子幼苗株高、基茎、叶片数、单株叶面积和总干质量等均呈减小趋势,株高、基径、叶片数和单株叶面积在重度子叶去除处理均显著小于对照和轻度子叶去除处理;小种子幼苗对子叶损失反应更为敏感,其株高、叶片数、单株叶面积和总干质量在中度子叶去除处理也显著小于对照(P<0.05);小种子幼苗根冠比随子叶去除强度增大而增大,在中度和重度子叶去除处理显著大于对照和轻度子叶去除处理(P<0.05)。(3)随着子叶去除强度增大,大小种子幼苗的比叶面积和比根长均逐渐增大,但大种子幼苗比枝长持续减小,轻度子叶去除处理的小种子幼苗比枝长最大。
关键词:辽东栎,种子大小,子叶去除,幼苗生长,形态学可塑性
中图分类号:Q948.1
文献标识码:A
文章编号:1000-3142(2021)09-1524-10
Abstract:Four treatments of cotyledon excisions (no cotyledon excision,excisions of 1/4,1/2 and 3/4 cotyledon,defined as control,light,medium,and high cotyledon excisions,respectively)were conducted to investigate the effects of seed size and different cotyledon excisions on the growth and morphological plasticity of Quercus wutaishanica seedlings planted in greenhouse pots. The results were as follows:(1)The shoot height (SH),basal stem diameter (BSD),leaf number (LN),leaf area per plant (LAPP),total dry mass (TDM),and root-shoot ratio (RSR)of Q. wutaishanica seedlings germinated from large seeds were higher than those of seedlings from small seeds in all treatments of cotyledon excision. There were significant differences in all above parameters,with the exception of SH,among seedlings from large and small seeds,on which cotyledons were lightly excised (P<0.05). Moreover,TDM of the former was significantly higher than that of the latter for the medium cotyledon excision (P<0.05). Also,higher specific leaf area (SLA)and specific root length (SRL)of seedlings from large seeds than those from small seeds were observed in all treatments of cotyledon excisions,while the specific shoot length (SSL)was detected in reverse. (2)With the intensity of cotyledon excision increasing,SH,BSD,LN,LAPP and TDM of seedlings from both large and small seeds appeared a trend of decline. SH,BSD,LN and LAPP of seedlings emerged from high cotyledon excision seeds were significantly lower than those of seedlings from control and light cotyledon excision seeds (P<0.05). Higher sensitivity to cotyledon excision was found in seedlings from small seeds than from large seeds,and SH,LN,LAPP and TDM of seedlings originated from small seeds,of which cotyledons were mediumly excised were also lower than those of seedlings from control seeds (P<0.05). RSR increased with the intensity of cotyledon excision increasing,and they were markedly higher in seedlings from seeds,regardless of whether they were large or small sized,mediumly and highly excised than those in seedlings from control and light cotyledon excision seeds (P<0.05). (3)With the intensity of cotyledon excision increasing,SLA and SRL gradually increased,while SSL of seedlings emerged from large seeds continuously decreased and the seedlings emerged from small seeds whose cotyledons were lightly excised had the maximum of SSL.
Key words:Quercus wutaishanica,seed size,cotyledon excision,seedling growth,morphological plasticity
富含营养的栎属(Quercus)植物种子是脊椎动物的重要食物资源,种子成熟散落后遭动物大量取食(孙书存和陈灵芝,2001; 高贤明等,2003; 闫兴富等,2011),即使在幼苗建立期间,捕食动物也可能以幼茎为搜寻线索(Pyare & Longland,2000)而大量取食其子叶(高贤明等,2003; 闫兴富等,2011,2012),从而增大其幼苗死亡的风险。植物从种子到幼苗的转变是其生活史中对环境因子反应最为敏感的阶段,在从种子萌发开始的幼苗建植过程中,种子贮藏的营养物质为种子萌发、幼苗生长和茎叶损伤后再生所需营养提供了保障(闫兴富等,2012)。有研究认为种子遭受取食危害后严重影响其萌发及其幼苗生长(Lombardo & McCarthy,2009; 刘艳等,2013; 刘影等,2017; 姜晓旭等,2018),特别是高强度子叶损失可减少幼苗根茎生物量积累(Shi et al.,2017),甚至导致幼苗建植失败(Mancilla-Leytón et al.,2013),但轻度子叶损失不会影响种子萌发率(Mancilla-Leytón et al.,2012),且可通过加快渗透(Finch-Savage & Clay,1994)或促进IAA等生長调节物质合成(Prewein et al.,2006)而促进种子快速萌发和同步出苗(Hou et al.,2010; Giertych & Suszka,2011; Perea et al.,2011; Shi et al.,2017)。子叶部分损失对幼苗生长发育无显著影响(Giertych & Suszka,2011),因为子叶中的营养物质在幼苗建立过程中已大量转移并贮藏于主根(Giertych & Suszka,2011; 闫兴富等,2012; 姜晓旭等,2018),而这些营养物质有助于幼苗在子叶或茎叶受损后的生长恢复和新芽萌生,可增大幼苗在林下成功建植的机会(Perea et al.,2011; 闫兴富等,2012)。
种子子叶损失对幼苗生长的影响可能与种子大小(Yi et al.,2015; Lebrija-Trejos et al.,2016)、子叶受损程度(Giertych & Suszka,2011; 姜晓旭等,2018)和损失部位(Hou et al.,2010)等因素有关,尤其是种群内和种群间甚至同一植株内普遍存在的种子大小差异(朱金雷和刘志民,2012)可能对子叶损失后幼苗的生长发育具有重要影响。贮藏更多营养物质的大种子萌发建立幼苗的机会更大(黄双全等,2007),且具有更强的耐受恶劣环境能力和对光照和养分等资源的竞争能力(Lebrija-Trejos et al.,2016)。因为种子营养物质向幼苗的转移在其早期发育过程中具有重要作用,所以,大种子萌发幼苗寿命更长,生物量积累更多(Jrgensen et al.,2019),而且大种子贮藏的营养物质远远多于种子萌发和幼苗生长的营养需要(Yi et al.,2015)。有研究认为,较大种子的更多营养物质不仅作为种子萌发和幼苗建立的营养来源(Yi et al.,2015),而且在大多情况下,动物是从距胚根更远的子叶末端开始消耗营养物质,这些被消耗的营养物质可能是植物对捕食动物的饱和作用或为扩散动物提供食物“回报”而额外储备的冗余营养(Perea et al.,2011; Yi et al.,2015),动物对种子的这一取食行为还与Prewein et al. (2006)报道的“栎属植物种子子叶近胚轴端单宁含量相对较高”和Hou et al. (2010)报道的“子叶顶端的营养物质对维持种子活力和幼苗建立更为重要”的种子结构相关特征高度吻合。因此,相同的子叶损失量对较大种子萌发幼苗生长发育的负面影响可能相对较小(Yi et al.,2015)。
辽东栎(Quercus wutaishanica)是我国暖温带落叶林的重要优势树种之一,其种子在秋季成熟散落后立即萌发并将子叶的大部分营养物质快速转移至主根,胚芽在翌年春季伸长形成幼苗(孙书存和陈灵芝,2001; 高贤明等,2003),这一特殊的萌发和幼苗建立方式不仅降低了种子脱水死亡和被动物捕食的风险,而且即使在幼苗子叶遭动物捕食的情况下仍能正常存活与生长(闫兴富等,2011,2012)。已有研究结果显示,即使在结实大年辽东栎种子也会因捕食动物取食而大量损失(孙书存和陈灵芝,2001; 闫兴富等,2011),但其种子结实量在不同年份间的明显波动(孙书存和陈灵芝,2001; 程积民等,2009)和种子大小变异(程积民等,2009; 闫兴富等,2011)在种群水平(Wang et al.,2000)和种子水平上(Yi & Yang,2010)对捕食动物的饱和效应可使动物捕食危害得以缓解。然而,种子萌发后辽东栎幼苗的子叶仍面临较高动物捕食风险(闫兴富等,2011,2012),已有研究采用人工切除子叶的方式模拟研究不同程度动物取食子叶对种子萌发和幼苗早期生长的影响(闫兴富等,2014),但其使用的种子大小较为均一,且幼苗生长时间相对较短,而大小各异的种子在去除子叶后对幼苗生长的影响仍值得进一步探索。本研究以不同大小的辽东栎种子为材料,在盆播条件下研究不同程度人工去除子叶对幼苗生长的影响,研究结果有助于进一步深入了解捕食动物取食种子危害对辽东栎幼苗生长和种群实生更新的影响机制,并为森林培育和退化植被恢复实践中的种苗繁育提供参考。
1 材料与方法
1.1 种子的采集和处理
实验用辽东栎种子采自宁夏六盘山林业局所属龙潭林场的大倒沟林区树龄约25年以上辽东栎灌丛植株。于2017年9月上旬,在采种林区选择相对较大(株高2 m以上)、冠层生长健壮的灌丛植株,在结果集中的枝条上摘取足够数量的不同大小的成熟辽东栎种子,大、小种子鲜重分别为(3.05±0.38)g (n=100)和(1.46±0.27)g (n=100)。于采摘次日带回实验室,以水浸法除去昆虫浸染的种子,于阴凉处晾干种子表面水分并经短期贮藏(约1周)后用于播种和实验幼苗的繁育。
1.2 研究方案和幼苗繁育
为避免冬季和早春低温造成幼苗死亡,盆栽实验在玻璃温室内进行,温室内光照强度约为自然全光照的55%,温室在冬季以普通暖气片供暖(11月1日—3月31日),白天温度约(23±5)℃,夜间温度约(16±3)℃;考虑到辽东栎幼苗发育早期喜荫的特点,在温室内用单层黑色尼龙网眼布进行遮阴,遮阴棚下相对光强约为自然全光照的18.9%。于2017年9月20日,取32个高23.5 cm、内径20 cm的塑料花盆,等分为2组,分别播种大种子和小种子,每组的16盆再分成4个小组(4盆为1小组)分别作为大种子和小种子幼苗的4个不同程度子叶去除处理,各播入大、小种子的4盆分别作为大、小种子幼苗的4次重复。取上述处理好的大、小辽东栎种子各480粒,将种子用75%乙醇进行表面消毒和清水冲洗后,自种子基部一端用单面刀片划开并去除种皮,其中1组种子(大、小种子各120粒)直接播种于花盆中作为对照(不去除子叶处理),其余3组分别为去除子叶1/4/、1/2和3/4 3个处理,用单面刀片分别将3个处理的种子切除子叶基部长度的1/4、1/2和3/4,上述4组处理分别定义为对照、轻度、中度和重度子叶去除处理。子叶去除处理后立即播入上述预先准备好的花盆中。
播种前先将经多年种植小麦熟化的风沙土装入花盆并充分浇水,种子子叶去除处理完成后立即进行播种;按上述方案在每一花盆内播入对应子叶去除处理的种子30粒,将种子胚根一端朝下播入土中,播种后覆盖厚约1 cm的湿润沙土。播种后约1周左右胚根伸长萌发,翌年4月初胚芽伸长出苗,约6周后出苗结束。出苗结束2周后,每盆选留生长健壮、植株大小基本一致的幼苗15株定苗,浇水后连根挖除多余幼苗。试验期间适时浇水保持盆内土壤湿润以满足种子萌发和幼苗生长对水分的需要,并定期进行松土、除草等日常管理。
1.3 幼苗收获和生长参数的测定与计算
2018年10月20日,收获上述各花盆的全部幼苗,花盆充分浇水后小心将幼苗从土中连根挖出,用自来水清洗干净后带回实验室,用洁净滤纸吸干表面水珠后测定记录幼苗的株高(shoot height,SH)、基径(basal stem diameter,BSD,用电子数显式游标卡尺测定)、叶片数(leaf number,LN )、单株叶面积(leaf area per plant,LAPP,用美国产LI-3100叶面积测定仪测定);将幼苗分根、茎和叶片装入纸袋中置于数字显示电子烘干箱中烘干(控制在85 ℃条件下),48 h后用电子天平分别称取根、茎和叶干质量;根据上述测定的数据计算幼苗的总干质量(total dry mass,TDM)、根冠比(root shoot ratio,RSR)、比叶面积(specific leaf area,SLA)、比根长(special root length,SRL)和比枝长(special shoot length,SSL)。相关参数的计算公式如下:TDM=根干质量+茎干质量+叶干质量;RSR=根干质量/(茎干质量+叶干质量);SLA=LAPP/叶干质量(cm2·g-1);SRL=主根长/主根干质量(cm·g-1);SSL=主茎长/主茎干质量(cm·g-1)。
1.4 数据统计分析
所有实验数据在统计分析前经平方根转换,用两因素方差分析法(two-way ANOVA)分析种子大小和子叶去除处理及其交互作用对幼苗生长参数是否具有显著影响,用最小显著差异法(LSD)分析幼苗生长参数在不同大小种子幼苗间和不同子叶去除處理间的差异显著性。全部数据统计分析均在SPSS 13.0中进行,结果中的数据以平均值呈现。
2 结果与分析
2.1 子叶去除对不同大小种子萌发幼苗生长的影响
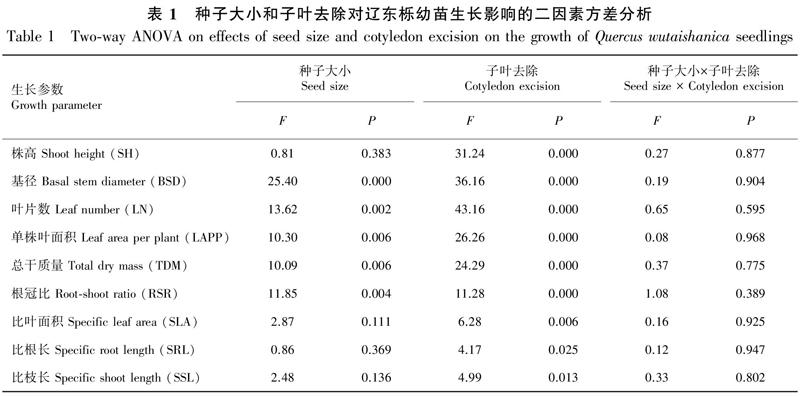
种子大小对幼苗基茎、叶片数和单株叶面积的影响均达极显著水平,但对株高无显著影响;子叶去除处理对上述参数均具有极显著影响,但种子大小和子叶去除处理的交互作用对上述4个生长参数的影响均不显著(表1)。
从图1可以看出,不同子叶去除处理的大种子幼苗株高均不同程度大于小种子幼苗,但大、小种子幼苗间株高差异均不显著;大、小种子幼苗株高在对照中均为最大(分别为14.40、13.37 cm),随子叶去除强度增大都依次减小,其中大种子幼苗在重度、小种子幼苗在中度和重度子叶去除处理的株高均显著小于对照(P<0.05),小种子幼苗在3个子叶去除处理间的差异也均达显著水平(P<0.05)。大种子幼苗基径均大于小种子幼苗,且对照和轻度子叶去除处理的不同大小种子幼苗间差异显著(P<0.05);不同大小种子幼苗基径都随子叶去除强度增大而减小,大种子幼苗在不同子叶去除处理及其与对照间差异均显著(P<0.05),小种子幼苗仅在重度子叶去除与对照、轻度子叶去除处理间差异显著(P<0.05)。
大种子幼苗叶片数在各处理都大于小种子幼苗,且前者仅在对照和轻度子叶去除处理显著大于后者(P<0.05);不同大小种子幼苗叶片数在对照中均为最大,3个子叶去除处理的大种子幼苗叶片数均显著减小(P<0.05),重度子叶去除处理还显著小于轻度子叶去除处理(P<0.05),小种子幼苗在不同子叶去除处理及其与对照间均差异显著(P<0.05)。大种子幼苗单株叶面积也都大于小种子幼苗,前者在轻度子叶去除处理显著大于后者(P<0.05);大种子幼苗单株叶面积在中度和重度子叶去除处理均显著小于对照和轻度子叶去除处理(P<0.05),而小种子幼苗除在中度和重度子叶去除处理间无显著差异外,其他处理及其与对照间均差异显著(P<0.05)。
2.2 幼苗生物量积累和形态学可塑性
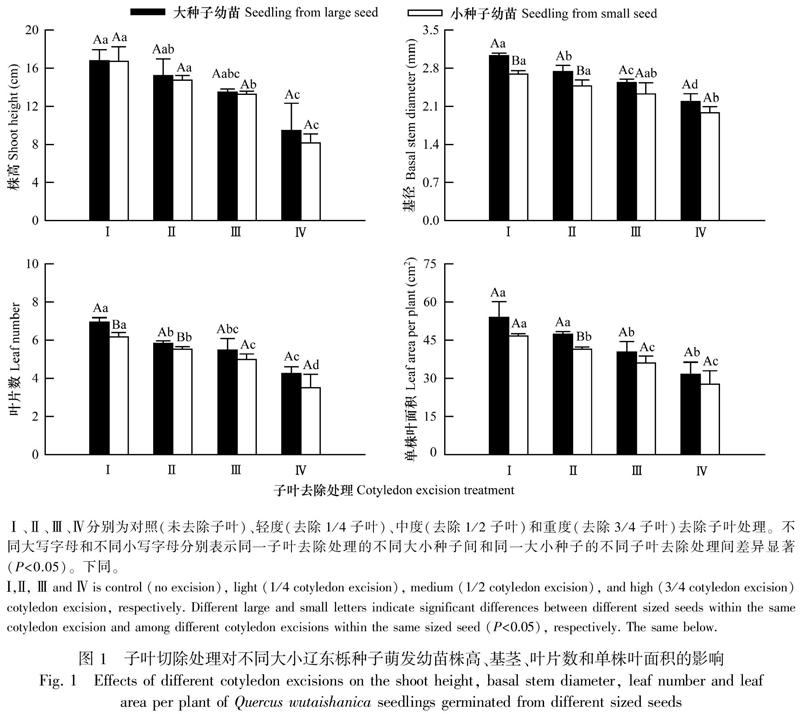
2.2.1 幼苗生物量积累及分配 种子大小和子叶去除均显著影响幼苗总干质量和根冠比,但两者的交互作用对幼苗总干质量及其分配(根冠比)无显著影响(表1)。
从图2可以看出,大种子幼苗总干质量均高于小种子幼苗,轻度和中度子叶去除处理的不同大小种子幼苗间差异显著(P<0.05);随着子叶去除强度增大,大、小种子幼苗总干质量分别从对照的0.53、0.44 g减小至0.37、0.34 g,其中大种子幼苗除在轻度和中度子叶去除处理间无显著差异外,其他处理及其与对照幼苗间差异均显著(P<0.05),小种子幼苗总干质量在3个子叶去除处理均显著小于对照(P<0.05),中度和重度子叶处理间差异均显著(P<0.05)。大种子幼苗根冠比在对照和3个不同强度子叶去除处理均大于小种子幼苗,不同大小种子幼苗间差异不显著;子叶去除增大了幼苗根冠比,大、小种子幼苗根冠比均在重度子叶去除处理最大(分别为0.73、0.68),其中,前者在轻度子叶去除处理显著大于对照(P<0.05),后者除在轻度子叶去除处理与对照间无显著差异外,其他处理及其与对照间差异均显著(P<0.05)。
2.2.2 幼苗形态学可塑性 种子大小对幼苗比叶面积、比根长和比枝长均无显著影响,而子叶去除处理对这些参数影响均显著,两者的交互作用对上述3个参数无显著影响(表1)。从图3可以看出,大种子幼苗比叶面积在不同子叶去除处理均大于小种子幼苗,差异不显著;大、小种子幼苗比叶面积均在对照最小(分别为244.53、220.47 cm2·g-1),且均随子叶去除强度增大逐渐增大,在重度子叶去除处理最大(分别为364.07、307.66 cm2·g-1),其中,小种子幼苗在重度子叶去除处理显著大于对照(P<0.05)。
大种子幼苗比根长在对照和不同强度子叶去除处理均大于小种子幼苗,不同大小种子幼苗间差异不显著;随着子叶去除强度增大,大、小种子幼苗比根长分别从对照的86.15、76.42 cm·g-1逐渐增大至重度子叶去除处理的114.39、109.95 cm·g-1,各处理及其与对照间无显著差异。对照和不同强度子叶去除处理大种子幼苗比枝长均小于小种子幼苗,差异不显著;大种子幼苗比枝长随着子叶去除强度增大从对照的最大值(100.73 cm·g-1)逐渐减小至重度子叶去除处理的73.62 cm·g-1,但仅有中度子叶去除处理显著小于对照(P<0.05),小种子幼苗在轻度子叶去除处理略大于对照,但在中度和重度子叶去除处理持续减小,各子叶去除处理及其与对照间差异均不显著。
3 讨论与结论
植物种群间、种群内甚至同一植物个体内部普遍存在种子大小的差异(朱金雷和刘志民,2012)。与小种子相比,大种子因含有较多营养物质而被种子扩散者有效贮藏并建立幼苗的机会更大,且大种子幼苗对不利环境条件的耐受能力以及对光照和养分等资源的竞争力更强(Lebrija-Trejos et al.,2016)。武高林和杜国祯(2006)认为幼苗早期阶段生长与初始大小密切相关,而幼苗初始大小可能直接取决于种子大小差异,因为在萌发过程中种子营养物质可转移并贮藏于幼苗中,从而使大种子幼苗发育出比小种子幼苗更为完善的根系(李荣平等,2004; 闫兴富等,2012)。本研究发现,辽东栎大种子幼苗株高、基茎、叶片数和单株叶面积等生长参数在各子叶去除处理均不同程度地大于小种子幼苗,其中轻度子叶去除处理幼苗基茎、叶片数和单株叶面积在不同大小种子幼苗间差异显著,而中度和重度子叶去除处理在不同大小种子幼苗间差异不显著。这些结果表明,辽东栎种子大小是影响幼苗生长的重要因素。一方面大种子借助其对捕食动物的饱和效应(Yi & Yang,2010)优势而具有比小种子更强的耐受动物取食危害能力;另一方面,大种子可通过其特殊的萌发方式发育出更为完善的主根,从而具有比小种子幼苗更强的耐受不良环境能力和资源竞争等方面的优势(Lebrija-Trejos et al.,2016)。
種子大小对幼苗存活的影响可能主要与幼苗早期阶段生物量分配模式和资源利用方式有关(武高林和杜国祯,2008)。本研究中,辽东栎大种子幼苗总干质量和根冠比在对照和不同强度子叶去除处理都高于小种子幼苗,且总干质量在轻度和中度子叶去除处理的不同大小种子幼苗间差异显著。幼苗这一干物质积累和分配模式可能与辽东栎种子秋季胚根快速伸长而胚芽翌年出土的出苗方式有关。一方面,处于萌发阶段的种子尽可能多地将营养转移至主根以减少子叶被动物取食造成的营养损失(闫兴富等,2012),从而使大种子幼苗积累干物质更多(何彦龙等,2007; Jrgensen et al.,2019),尤其在高强度子叶损失情况下,具有比小种子幼苗更为明显的干物质积累优势(Yi et al.,2015);另一方面,随着幼苗生长后期源—库关系转化和对种子大小依赖的消失或减弱(何彦龙等,2007; Shi et al.,2017),对相对生长率较低的大种子幼苗来说,其后期地上部分生长可能主要依赖于土壤养分资源,生长优势逐渐减弱(何彦龙等,2007; 黄双全等,2007),因而其根冠比大于小种子幼苗。本研究发现,尽管种子大小对辽东栎幼苗比叶面积、比根长和比枝长影响不显著,但大种子幼苗比叶面积和比根长在对照和不同强度子叶去除处理均大于小种子幼苗,而比枝长则相反,表明不同大小种子幼苗形态学可塑性存在一定差异,何彦龙等(2007)也曾报道,草本植物黄帚橐吾(Ligularia virgaurea)不同大小种子幼苗比叶面积并无明显差异,比叶面积变化作为植物维持最优捕获光能能力的一种机制,可通过增大单位叶面积光能捕获量而提高光能利用率和相对生长率。本研究结果与一些研究报道的“小种子幼苗相对生长率较高”(Wu et al.,2006; 何彦龙等,2007)的结论有较大出入,较小的比叶面积和相对生长率可能有助于小种子幼苗通过维持更大的碳水化合物和矿质资源库来增强对环境胁迫的耐受能力(Westoby et al.,1992),而大种子幼苗较高的相对生长率可能导致生长后期主根营养过度透支而变得纤细,从而表现为比根长增大的形态学可塑性变化(许中旗等,2009)。小种子幼苗比枝长较大可能是其在发育早期采取的一种优先保障地上部分营养供应的策略,但发育后期因营养枯竭变得“消瘦”而增大比枝长,有关青藏高原高寒草甸植物的研究也报道了小种子物种幼苗具有更大形态可塑性的结论(Wu et al.,2006),但该结果与本研究的幼苗比叶面积和比根长的结论并不一致,产生这些结果差异的原因还有待于进一步研究。
栎属植物种子子叶是幼苗发育的主要营养物质来源,在种子萌发和幼苗的建立、生长与存活中都具有重要作用(高贤明等,2003; 闫兴富等,2012),较大种子能够耐受一定程度子叶损失仍可建立幼苗(Fukumoto & Kajimura,2000; Giertych & Suszka,2011),而且适度子叶损失可能会减少种子内多酚类生长抑制物质向幼苗转移而促进幼苗生长(刘艳等,2013),因为栎属植物种子的营养贮藏远大于幼苗生长发育的需要,甚至大种子的冗余营养物质还可通过快速萌发转移至主根后以残留的子叶“诱使”捕食动物“优先”将其取食(高贤明等,2003; 闫兴富等,2012; Yi et al.,2015),从而降低整个种子损失的风险。因此,人工去除部分子叶可作为一种促进种子整齐萌发的技术应用于容器苗生产实践(Mancilla-Leytón et al.,2013; 史文辉等,2018; Shi et al.,2018)。然而,高强度子叶损失显著影响幼苗生长和根系发育,并对后续的土壤养分和水分吸收产生影响(Giertych & Suszka,2011; Mancilla-Leytón et al.,2013),甚至有研究报道Pasania edulis幼苗的生长受昆虫取食种子危害的负面影响可持续3年之久(Sone et al.,2016)。本研究中,随着子叶去除强度增大,辽东栎大、小种子幼苗株高、基茎、叶片数和单株叶面积等均呈减小趋势,且在重度子叶去除处理均显著小于对照和轻度子叶去除处理,尤其小种子幼苗株高、叶片数和单株叶面积在中度子叶去除处理均显著小于对照,表明高强度子叶损失严重影响幼苗存活与生长(刘艳等,2013; 闫兴富等,2014; 姜晓旭等,2018),甚至有研究发现高强度子叶损失可影响幼苗第二个生长季节的株高生长和干物质积累(Giertych & Suszka,2011)。因此,栎属植物种子耐受子叶损失的量可能有一个临界值,而且不同大小种子的临界值存在差异。
研究发现,随着子叶去除强度增大,辽东栎不同大小种子幼苗总干质量均逐渐减小,根冠比则逐渐增大;总干质量在不同强度子叶去除处理均显著小于对照,小种子幼苗生物量分配对子叶去除处理的反应比大种子幼苗更为敏感,尤其根冠比在中度和重度子叶去除处理均显著大于对照,而大种子幼苗仅在轻度子叶去除处理与对照间差异显著。一方面,低强度子叶损失不同程度地抑制幼苗茎、叶生物量和总生物量积累,对根冠比影响较小(刘影等,2017),但中度和重度子叶损失导致幼苗茎、叶生物量及总生物量积累显著减少(Perea et al.,2011; 閆兴富等,2014; 刘影等,2017),甚至导致幼苗建植失败(Mancilla-Leytón et al.,2013);另一方面,幼苗子叶部分损失后倾向于优先将更多营养物质分配至根系还可增强幼苗茎叶的萌生更新能力(Shi et al.,2017; Yi et al.,2019)和幼苗发育过程中对环境风险的防御能力(闫兴富等,2012)。辽东栎主根干物质积累对种子子叶的依赖及幼苗可能在种子萌发期间建立起上述防御机制,其特殊的萌发和出苗方式既有利于幼苗营养物质分配在秋季和春季之间进行时间上的权衡,又有利于在主根和留土子叶之间进行空间上的权衡,从而确保幼苗茎杆发育的营养来源在子叶遭动物取食损失后转向依赖主根贮藏的营养物质(高贤明等,2003; 闫兴富等,2011,2012)。事实上,捕食动物对辽东栎幼苗子叶取食的同时,往往连同顶芽一起取食(闫兴富等,2012),主根之所以能稳稳地“铆”在土壤中完全得益于其从子叶获取的大量营养物质。因此,辽东栎子叶受损幼苗根冠比增大可能是其应对子叶营养损失的一种重要机制,而这些贮藏于主根中的营养物质也为幼苗顶芽受损(包括幼苗发育期间茎叶受损)后的萌生提供了营养保障(闫兴富等,2012)。
辽东栎幼苗可对子叶损失引起的营养亏缺表现出一定的形态学可塑性(闫兴富等,2014),随着子叶去除强度增大,幼苗比叶面积和比根长均逐渐增大,大种子幼苗比枝长持续减小,而小种子幼苗比枝长在轻度子叶去除处理略有增大,表明幼苗对子叶损失引起的营养亏缺能够在形态学方面做出有利于提高资源利用效率的补偿性响应,即通过增大比叶面积来提高光能利用率,或通过增大比根长来增加根系对土壤深层养分和水分的吸收(Westoby et al.,1992)。有研究认为,子叶损失对幼苗地上部分营养的限制可能更大(Giertych & Suszka,2011),从而加剧胚根在种子萌发期间快速伸长对营养的需求与地上部分间的竞争,子叶残留的少量营养在满足幼苗发育早期的茎秆伸长后,难以维持新梢抽生所需的营养。因此,对子叶去除处理幼苗来说,实验结束时茎秆仍是种子萌发期间上胚轴伸长的产物,比枝长要小于对照幼苗。小种子幼苗比枝长在轻度子叶去除处理最大可能与营养较少的小种子在萌发过程中对子叶损伤刺激的应激反应有关,即幼苗发育早期在茎生长投资过大而后期缺乏持续的营养供应导致比枝长增大。
参考文献:
CHENG JM,ZHAO LP,CHENG J,2009. Seed quality and forest regeneration of a 60-year Quercus liaotungensis forest in the Ziwuling Region,Northwestern China[J]. J Beijing For Univ,31(2):10-16.[程积民,赵凌平,程杰,2009. 子午岭60年辽东栎林种子质量与森林更新[J]. 北京林业大学学报,31(2):10-16.]
FINCH-SAVAGE WE,CLAY HA,1994. Water relations of germination in the recalcitrant seeds of Quercus robur L.[J]. Seed Sci Res,4(3):315-322.
FUKUMOTO H,KAJIMRUA H,2000. Effects of insect predation on hypocotyls survival and germination success of mature Quercus variabilis acorns[J]. J For Res,5(1):31-34.
GAO XM,DU XJ,WANG ZL,2003. Comparison of seedling recruitment and establishment of Quercus wutaishanica in two habitats in Dongling Mountainous area,Beijing[J]. Chin J Plant Ecol,27(3):404-411.[高贤明,杜晓军,王中磊,2003. 北京东灵山区两种生境条件下辽东栎幼苗補充与建立的比较[J]. 植物生态学报,27(3):404-411.]
GIERTYCH MJ,SUSZKA J,2011. Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing[J]. Ann For Sci,68(2):433-442.
HE YL,WANG MT,DU GZ,2007. Seed size effect on seedling growth under different light conditions in the clonal herb Ligularia virgaurea in Qinghai Tibet Plateau[J]. Acta Ecol Sin,27(8):3091-3098.[何彦龙,王满堂,杜国祯,2007. 不同光照处理下青藏高原克隆植物黄帚橐吾(Ligularia virgaurea)种子大小对其幼苗生长的影响[J]. 生态学报,27(8):3091-3098.]
HOU XG,YI XF,YANG YQ,et al.,2010. Acorn germination and seedling survival of Quercus variabilis:effects of cotyledon excision[J]. Ann For Sci,67(7):711-718.
HUANG SQ,LIU GX,HAN JG,2007. Effects of seed mass and sowing depth on seedling establishment[J]. Partac Sci,24(6):44-49.[黄双全,刘桂霞,韩建国,2007. 种子大小和播种深度对种苗建植的影响[J]. 草业科学,24(6):44-49.]
JIANG XX,LI GL,SHI WH,et al.,2018. Effects of cotyledon loss intensity and time on seedling growth status and reserves translocation in Quercus variabilis [J]. Sci Silv Sin,54(8):56-64.[姜晓旭,李国雷,史文辉,等,2018. 子叶丢失强度和时间对栓皮栎幼苗生长状况和营养物质转移的影响[J]. 林业科学,54(8):56-64.]
JRGENSEN MS,LABOURIAU R,OLESEN B,2019. Seed size and burial depth influence Zostera marina L. (eelgrass)seed survival,seedling emergence and initial seedling biomass development[J]. PLoS ONE:e0215157.
LEBRIJA-TREJOS E,REICH PB,HERNNDEZ A,et al.,2016. Species with greater seed mass are more tolerant of conspecific neighbours:A key driver of early survival and future abundances in a tropical forest[J]. Ecol Lett,19(9): 1071-1080.
LI RP,JIANG DM,LIU ZM,et al.,2004. Effects of sand-burying on seed germination and seedling emergence of six psammophytes species[J]. Chin J Appl Ecol,15(10):1865-1868.[李荣平,蒋德明,刘志民,等,2004. 沙埋对六种沙生植物种子萌发和幼苗出土的影响[J]. 应用生态学报,15(10):1865-1868. ]
LIU Y,LI QM,LIU Y,et al.,2013. Effects of mechanical treatments on germination and growth of Quercus fanshanensis and Q. robur seeds[J]. Sci Silv Sin,49(4):39-45.[刘艳,李庆梅,刘勇,等,2013. 机械处理对房山栎和夏栎种子萌发和生长的影响[J]. 林业科学,49(4):39-45.]
LIU Y,ZHAO Y,CUI D,et al.,2017. Effect of cotyledon damage on the early growth of Sophora alopecuroides seedlings[J]. Acta Pratacult Sin,26(8):139-145.[刘影,赵玉,崔东,等,2017. 子叶损伤对苦豆子幼苗早期生长的影响[J]. 草业学报,26(8):139-145.]
LOMBARDO JA,MCCARTHY BC,2009. Seed germination and seedling vigor of weevil-damaged acorns of red oak[J]. Can J For Res-Revue Canadienne De Recherche Forestiere,39(8):1600-1605.
MANCILLA-LEYTN JM,CAMBROLL J,FIGUEROA ME,et al.,2013. Growth and survival of cork oak (Quercus suber)seedlings after simulated partial cotyledon consumption under different soil nutrient contents[J]. Plant Soil,370(1):381-392.
MANCILLA-LEYTN JM,CAMBROLL J,MARTN VICENTE ,2012. The impact of the common rabbit on cork oak regeneration in SW Spain[J]. Plant Ecol,213:1503-1510.
PEREA R,MIGUEL AS,GIL L,2011. Leftovers in seed dispersal:ecological implications of partial seed consumption for oak regeneration[J]. J Ecol,99(1):194-201.
PREWEIN C,ENDEMANN M,REINOHL V,et al.,2006. Physiological and morphological characteristics during development of pedunculate oak (Quercus robur L.)zygotic embryos[J]. Trees,20(1):53-60.
PYARE S,LONGLAND WS,2000. Seedling-aided cache detection by heteromyid rodents[J]. Oecologia,122(1):66-71.
SHI WH,BLOOMBERG M,LI GL,2017. Combined effects of cotyledon excision and nursery fertilization on root growth,nutrient status and outplanting performance of Quercus variabilis container seedlings[J]. PLoS ONE,12(5):e0177002.
SHI WH,LI GL,SU SC,et al.,2018. Combined effects of cotyledon excision and nursery fertilization on field performance of Quercus variabilis container seedlings[J]. Sci Silv Sin,54(1):64-73.[史文輝,李国雷,苏淑钗,等,2018. 子叶切除与苗圃施肥对栓皮栎容器苗造林效果的影响[J]. 林业科学,54(1):64-73.]
SHI WH,VILLAR-SALVADOR P,JACOBS DF,et al.,2018. Simulated predation of Quercus variabilis acorns impairs nutrient remobilization and seedling performance irrespective of soil fertility[J]. Plant Soil,423:295-306.
SONE K,OISHI K,SHIBATO T,et al.,2016. Effects of acorn attack by curculio weevils on the germination and early growth of Pasania edulis (Makino)seedlings[J]. J For Res,21(4):185-192.
SUN SC,CHEN LZ,2001. The effects of animal removal and groundcover on the fate of seeds of Quercus liaotungensis [J]. Acta Ecol Sin,21(1):80-85.[孙书存,陈灵芝,2001. 动物搬运与地表覆盖物对辽东栎种子命运的影响[J]. 生态学报,21(1):80-85.]
WANG W,MA KP,LIU CR,2000. Seed shadow of Quercus liaotungensis in a broad-leaved forest in Dongling Mountain[J]. Acta Bot Sin,42(2):195-202.
WESTOBY M,JURADO E,LEISHMAN M,1992. Comparative evolutionary ecology of seed size[J]. Trends Ecol Evol,7(11): 368-372.
WU GL,CHEN M,ZHOU XH,et al.,2006. Response of morphological plasticity of three herbaceous seedlings to light and nutrition in the Qinghai-Tibetan Plateau[J]. Asian J Plant Sci,5(4):635-642.
WU GL,DU GZ,2008. Relationship between seed size and seedling growth strategy of herbaceous plant:A review[J]. Chin J Appl Ecol,19(1):191-197.[武高林,杜国祯,2008. 植物种子大小与幼苗生长策略研究进展[J]. 应用生态学报,19(1):191-197.]
WU GL,DU GZ,SHANG ZH,2006. Contribution of seed size and its fate to vegetation renewal:A review[J]. Chin J Appl Ecol,17(10):1969-1972.[武高林,杜国祯,尚占环,2006. 种子大小及其命运对植被更新贡献研究进展[J]. 应用生态学报,17(10):1969-1972.]
XU ZQ,HUANG XR,XU CL,et al.,2009. The impacts of light conditions on the growth and morphology of Quercus Mongolica seedlings[J]. Acta Ecol Sin,29(3):1121-1128.[许中旗,黄选瑞,徐成立,等,2009. 光照条件对蒙古栎幼苗生长及形态特征的影响[J]. 生态学报,29(3): 1121-1128.]
YAN XF,DU Q,SHI C,et al.,2011. Seedling regeneration of Quercus liaotungensis in Liupan Mountains,China[J]. Chin J Plant Ecol,35(9):914-925.[閆兴富,杜茜,石淳,等,2011. 六盘山区辽东栎的实生苗更新及其影响因子[J]. 植物生态学报,35(9):914-925.]
YAN XF,FANG S,SHI C,et al.,2014. Effects of simulated cotyledon predation on the seed germination and early seedling growth of Quercus wutaishanica [J]. Chin J Ecol,33(4):973-981.[闫兴富,方苏,石淳,等,2014. 模拟动物取食子叶对辽东栎种子萌发和幼苗早期生长的影响[J]. 生态学杂志,33(4):973-981.]
YAN XF,ZHOU LB,ZHANG KW,et al.,2012,Cotyledon loss and its effects on survival and growth of Quercus wutaishanica seedlings under different densities[J]. Chin J Plant Ecol,36(8):831-840.[闫兴富,周立彪,张靠稳,等,2012. 不同密度下辽东栎幼苗子叶丢失及其对幼苗存活和生长的影响[J]. 植物生态学报,36(8):831-840.]
YI XF,BARTLOW AW,CURTIS R,et al.,2019. Responses of seedling growth and survival to post-germination cotyledon removal:An investigation among seven oak species[J]. J Ecol,107(4):1817-1827.
YI XF,YANG YQ,2010. Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena [J]. Plant Ecol,209(2):291-300.
YI XF,WANG ZY,LIU CQ,et al.,2015. Acorn cotyledons are larger than their seedlings need:Evidence from artificial cutting experiments[J]. Sci Rep-UK,5:8112.
ZHU JL,LIU ZM,2012. Major terminologies and concepts in seed dispersal biology[J]. Chin J Ecol,31(7):2397-2403.[朱金雷,刘志民,2012. 种子传播生物学主要术语和概念[J]. 生态学杂志,31(7):2397-2403.]
(责任编辑 李 莉)
