基于农艺及品质性状与SSR标记的青贮玉米品种遗传多样性分析
2021-10-05刘少荣田红丽易红梅康定明范亚明葛建镕成广雷王凤格
刘少荣 杨 扬 田红丽 易红梅 王 璐 康定明 范亚明 任 洁 江 彬 葛建镕 成广雷,* 王凤格,*
1 北京市农林科学院玉米研究中心 / 玉米DNA指纹及分子育种北京市重点实验室, 北京 100097; 2 中国农业大学农学院, 北京100193
玉米是中国重要的粮饲兼用作物, 2019年种植面积为4128万公顷[1], 超过稻谷和小麦, 位居农作物第一。随着中国粮改饲产业结构调整及畜牧业发展, 饲用玉米, 特别是青贮玉米的种植面积在逐步扩大, 为畜牧业供给充足的优质饲料[2]。青贮玉米作为优质饲料, 具有产量高、品质好、饲用价值高等特点, 已在畜牧业发展、生态环境保护、农业增收中占有不可或缺的地位, 大力开展青贮玉米的品种选育是解决当下畜牧业迅速发展带来的饲料缺乏问题的有效途径[3]。然而中国青贮玉米研究起步较晚,虽然审定了不少品种, 但对这些品种的遗传来源并不清楚, 所以亟需摸清我国青贮玉米品种遗传背景现状。近年来, 对青贮玉米的研究主要集中在高产优质[4-5]、生态适应性[6-8]和配套栽培技术[9-10]等方面,关于青贮玉米遗传多样性分析的报道不多。
遗传多样性分析是作物种质资源研究的一种重要手段, 旨在了解和掌握不同品种之间的遗传差异,为推动植物育种与遗传改良奠定基础。遗传多样性分析可以从形态学性状和分子标记2个方面进行,形态学性状是从植物的表型性状来区分品种差异,分子标记是通过遗传物质反映品种间遗传变异程度[11]。迄今为止, 形态学性状已被广泛应用到玉米[12]、谷子[13]、水稻[14]、马铃薯[15]等农作物的遗传多样性研究中。在利用形态学性状对青贮玉米品种进行遗传多样性研究方面, 柴华[16]用10个形态学标记将36个青贮玉米自交系划分为五大类, 为选育优质青贮玉米品种提供依据; 吴建忠等[17]基于22个品质性状对14个青贮玉米品种进行遗传变异分析, 发现脂肪、木质素和淀粉的变异系数较大, 分别为20%、19%和16%, 为青贮玉米的品质改良提供了参考。SSR标记具有检测简便快捷、多态性高和重复性好等优点, 被迅速推广应用。李齐向等[18]采用23对SSR标记对4个代表国内主要杂种优势群的普通玉米品种和65个青贮玉米自交系进行聚类分析, 将大部分未知系谱来源的青贮玉米划分为Lancaster群、旅大红骨类群、塘四平头群和Reid群, 初步明确了其系谱来源。
国内外关于玉米遗传多样性的研究, 主要聚集在种质资源多样性评价、杂种优势群划分等方面,而针对青贮玉米品种遗传多样性的评估鲜有报道。本研究对141个来源于2002—2020年国家及各省区(市)审定的青贮玉米品种, 基于其13个农艺及品质性状和40个SSR标记, 并结合品种来源的生态区,进行了遗传多样性分析, 以了解各生态区青贮玉米品种的农艺、品质性状及遗传分化特点, 为青贮玉米的新品种选育及推广种植提供理论依据和参考。
1 材料与方法
1.1 材料
试验材料选自2002—2020年通过国家及各省区(市)审定的141个青贮玉米杂交种。参照2016年国家青贮玉米品种区域试验的生态组别划分, 并结合播种区域、播种时间、品种类型和选育单位等因素将供试品种划分为4个生态区, 分别为东华北区、黄淮海区、西北区和南方区, 详细品种审定来源及生态区分布见表1。农艺及品质分析材料: 在141个青贮玉米品种中, 有19个品种具有多个生态区或年份审定来源, 其中13个品种具有2个审定来源, 3个品种具有3个审定来源, 3个品种具有4个审定来源,即青贮玉米样品由141个扩增到169个。SSR标记分析材料: 包括141个青贮玉米品种和5个普通玉米品种(德美亚1号、郑单958、苏玉29、先玉335、农大108)。从试验材料中选用5个国家或省区(市)级青贮玉米区试对照品种(雅玉青贮8号、雅玉青贮26、大京九26、京九青贮16、桂青贮1号)、2个推广面积较大的粮饲兼用品种(中玉335、京科968)和5个国家普通玉米区试对照品种作为参考对照。

表1 样品信息统计Table 1 Sample information statistics
1.2 试验方法
1.2.1 农艺及品质性状 农艺及品质数据均来自于国家及各省区(市)青贮玉米审定公告, 其中包括13个性状, 分别为生育期、株高、穗位高、绿叶数、穗长、穗行数、粗蛋白、中性洗涤纤维、酸性洗涤纤维、淀粉、干重、鲜重和种植密度。凯氏定氮法测定粗蛋白含量, 旋光法测定淀粉含量, Van Soest法测定中性洗涤纤维和酸性洗涤纤维含量, 具体步骤参见GB/T25882-2010[19], 其他性状测定方法参见审定公告。
1.2.2 SSR标记 采用的40对SSR引物, 均为玉米品种鉴定行业标准中发布的引物, 具有多态性好,均匀分布在玉米的10条染色体上的特征, 具体引物名称、序列及片段长度等信息参考已发表的文献[20-21]。
1.2.3 DNA制备和SSR基因分型 采用改良CTAB法[22]提取供试材料基因组DNA, DNA浓度和质量用紫外分光光度计(Nanodrop 2000)测定, 根据测量值调节工作液浓度。SSR-PCR体系: 反应液总体积为20 μL, 包括2×TaqPlus Master Mix 10 μL,ddH2O 7.75 μL, 引物0.25 μL和DNA样品2 μL。PCR程序: 预变性95℃ 5 min;变性94℃ 40 s,退火60℃35 s, 延伸72℃ 45 s, 35个循环; 延伸72℃ 10 min;4℃保存。PCR产物检测: 采用10重PCR产物电泳检测的方法。向96孔电泳板的单个孔中分别加入2 μL 10重PCR的混合产物、10 μL含有1% GS3730-500分子量内标的甲酰胺。将上述混合样品放入PCR仪中95℃变性5 min, 4℃保存10 min, 2000转 min–1离心30 s后, 于ABI 3730XL DNA分析仪上进行荧光毛细管电泳。预电泳时间2 min, 15 kV, 电泳时间30 min, 15 kV, 电泳原始数据由Data Collection软件收集, 用SSR Analyser (v1.2.4)指纹分析器[23]对电泳数据基因分型分析。
1.3 数据统计与分析
采用SPSS 25软件对农艺及品质数据进行平均值、标准差和极值的统计, 并进行方差分析; 利用Microsoft Excel 2016计算变异系数和Simpson多样性指数。基于R语言scale函数对农艺及品质数据标准化处理, 再利用dist函数, 选择“euclidean”方法计算出品种间的欧式距离。将上述品种间的欧式距离矩阵导入PowerMarker V3.25[24]软件得到农艺及品质性状NJ (Neighbor-Joining)聚类结果,结合MEGA7[25]软件绘制聚类图。采用Power Marker V3.25软件对SSR基因型数据进行分析,计算不同生态区品种的等位变异数、基因型数、基因多样性、杂合度和PIC值。同时基于Nei’s(1973)方法计算品种间的遗传距离, 得到SSR标记NJ聚类结果, 结合MEGA7软件绘制聚类图。基于欧式距离矩阵和遗传距离矩阵, 选用多变量统计分析软件MVSP V3.22[26]对品种进行主成分分析并绘制PCA图。
2 结果与分析
2.1 农艺及品质性状遗传多样性分析
对169个青贮玉米样品的13个农艺及品质性状进行了描述性统计(表2), 各性状存在不同程度的变异, 变异系数分布在10.30%~30.31%之间,平均为16.01%, 其中干重和酸性洗涤纤维的变异系数超过20%, 其他性状变异系数在10%~20%之间。Simpson多样性指数的变化区间在0.50~0.71, 平均为0.60,穗长多样性指数最高, 粗蛋白多样性指数最低。在青贮玉米品质性状中, 粗蛋白和酸性洗涤纤维表现出较丰富的遗传多样性。
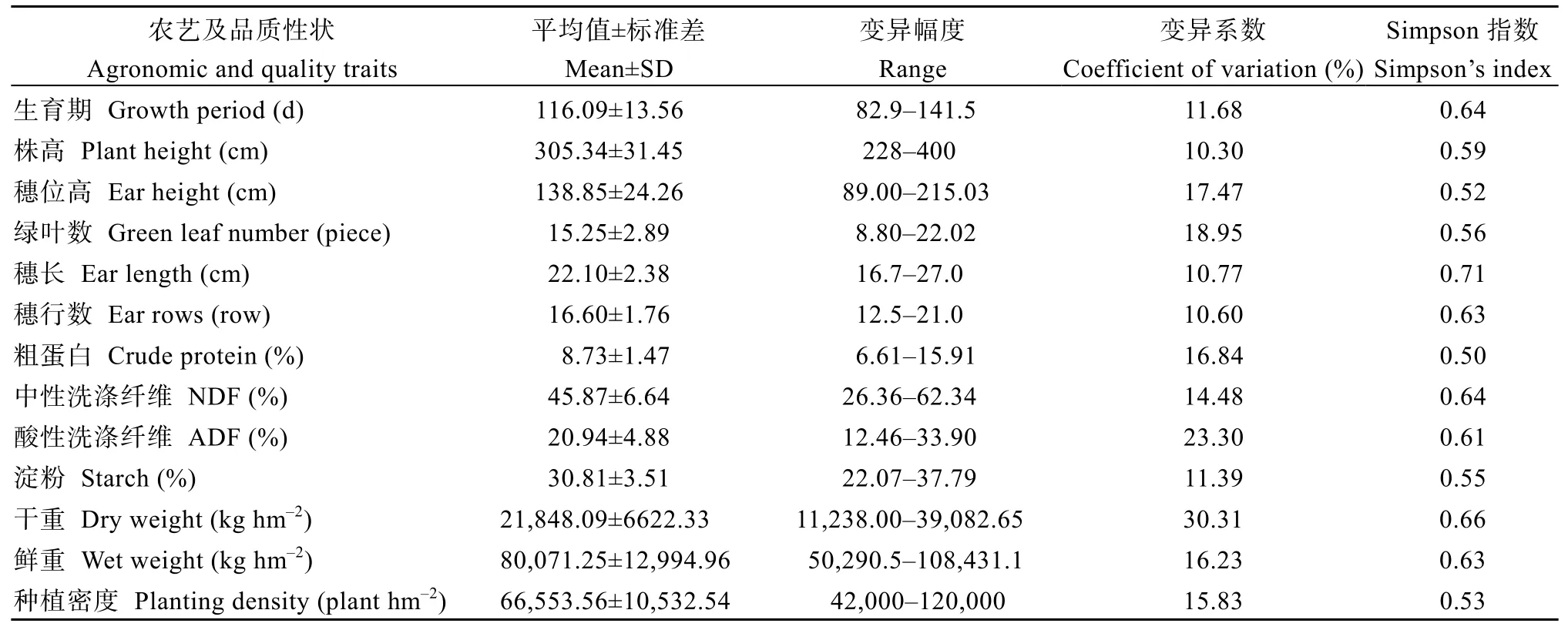
表2 169个样品的农艺及品质性状特征Table 2 Agronomic and quality characteristics of 169 samples
基于13个农艺及品质性状对供试品种进行聚类分析, 从图1-A可知, 169个青贮玉米样品可被划分为5个组, 大部分样品聚集在X3、X4组。X1组包括以桂青贮1号为代表的19个样品, X2组包括以中玉335为代表的13个样品, X3组包括以京九青贮16、京科968及雅玉青贮8号为代表的72个样品, X4组包括以大京九26为代表的45个样品, X5组包括以雅玉青贮26为代表的20个样品。对农艺及品质性状聚类组群进行主成分分析(图1-B), 根据5个聚类组群的具体分布情况将坐标图划分为I、II、III、IV四个区域, 从整体上来看, 各组群分布均相对集中, X1、X5组分布在I、III区, 但主要集中在III区,X2组主要分布在I区, X3组主要分布在I、II区, X4组分布在III、IV区。各组群坐标分布结果与聚类分组结果基本一致。
2.2 SSR标记遗传多样性分析
40对SSR引物在141个青贮玉米品种中共检测到482个等位变异, 每对引物等位变异数量范围为3~27个, 平均为12.05个; 基因多样性的变化范围为0.29~0.90, 平均为0.71; 杂合率的变化范围为0.27~0.94, 平均为0.68; PIC的变化区间为0.27~0.88, 平均为0.68。
以普通玉米为参考对照, 分析青贮玉米品种的遗传背景, 对141个青贮玉米品种和5个普通玉米品种进行聚类分析(图2-A), 供试品种被划分为5个组。S1组包括以雅玉青贮28、雅玉青贮8号和德美亚1号为代表的33个品种, S2组包括以大京九26为代表的24个品种, S3组包括以京科968和郑单958为代表的14个品种, S4组包括以京九青贮26、苏玉29和先玉335为代表的62个品种, S5组包括以中玉335和农大108为代表的13个品种。SSR标记聚类组群的主成分分析显示, 虽然存在极少数的离散品种, 但整体上各组群分布相对集中(图2-B),基本上各自占据相应的位置。S1、S3组分布均主要集中在II区, S2、S5组在III和IV区均有分布, S4组分布在I、II、III区, 但主要聚集在I、III区。各组群坐标分布结果与聚类分组结果一致, 且两者分析结果可以相互佐证。
2.3 不同生态区品种遗传多样性分析
为了探究各生态区品种间是否存在遗传差异,对不同生态区品种进行方差分析及遗传多样性比较(表3和表4)。结果表明, 东华北品种各性状相对居中, 基因多样性、杂合度和PIC均最低, 分别为0.68、0.65和0.64; 黄淮海品种生育期最短, 达到显著水平, 等位变异数最低, 为7.40; 西北品种生育期、穗位高、干重和鲜重最高, 且均达到显著水平,等位变异数和基因型数均最高, 分别为9.13和16.30;南方品种穗长、穗行数和鲜重最低, 粗蛋白最高, 且均达到显著水平, 基因型数最低, 为10.30, 基因多样性、杂合度和PIC均为最大, 分别为0.71、0.72和0.68。结果表明, 西北和南方品种表现出丰富的遗传多样性, 出现不同程度的性状分化, 产生了具有地方性特征的性状特点。
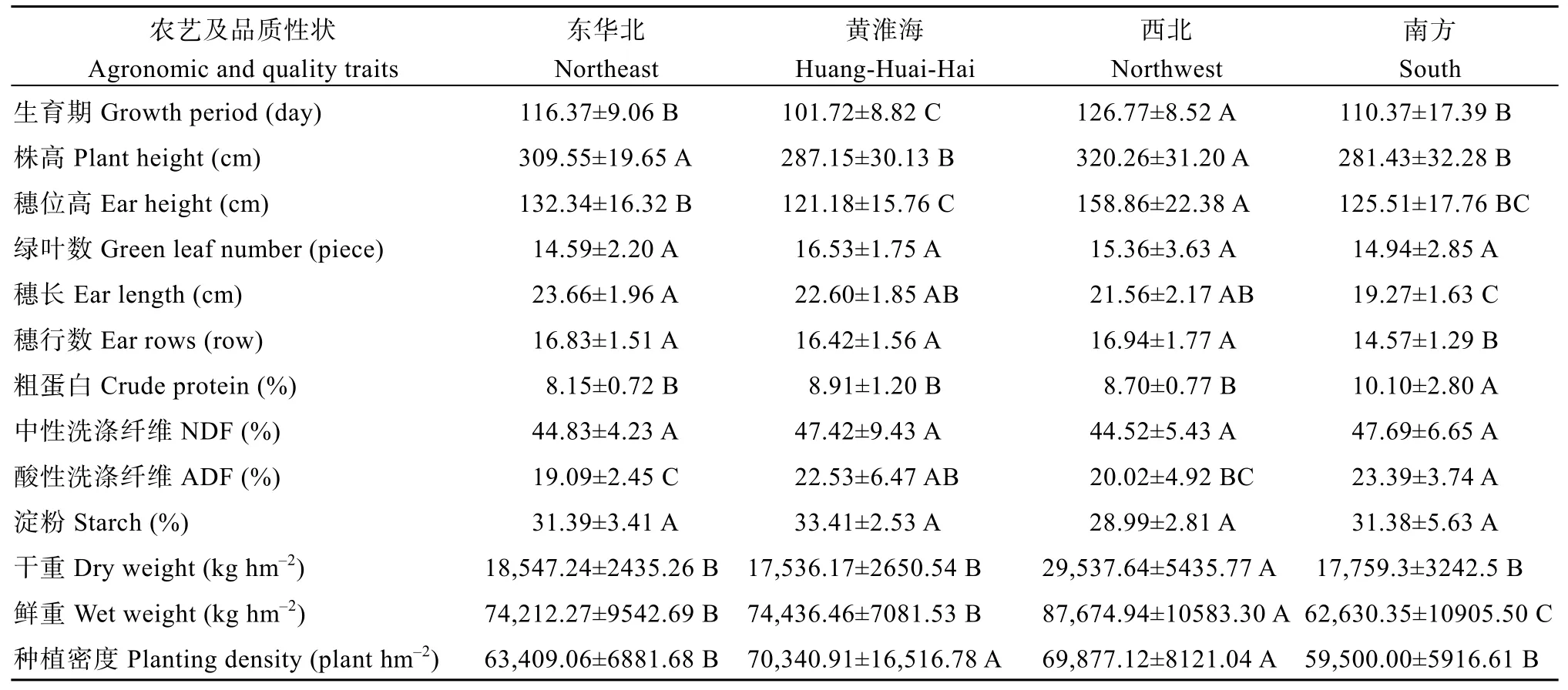
表3 4个生态区品种农艺及品质性状的方差分析Table 3 Analysis of variance of agronomic and quality traits of varieties from four ecological regions
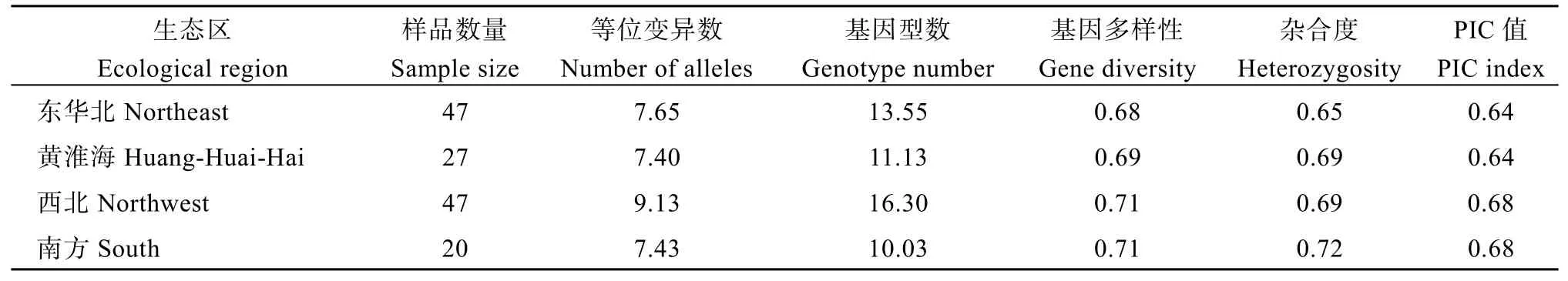
表4 4个生态区品种间的遗传多样性比较Table 4 Comparison of genetic diversity among varieties from four ecological regions
分析不同生态区品种在两类方法的聚类分布情况。由农艺及品质性状聚类可知(图2-A), 有3个生态区的大多数品种聚在相同的组群, 其中37个西北样品(62.7%)聚于X4组, 28个黄淮海样品(84.8%)和13个南方样品(59.1%)聚于X3组, 但东华北样品分布相对离散, 24个样品(43.6%)聚于X3组, 11个样品(20%)聚于X1组。各生态区品种之间的遗传距离结果表明, 南方品种与东华北、黄淮海和西北品种遗传距离较大, 分别为0.054、0.047、0.046, 而东华北、黄淮海和西北品种两两间遗传距离较小, 均在0.01左右。由SSR标记聚类可知(图2-A), 15个南方品种(75%)聚于S1组, 东华北、黄淮海和西北品种主要聚于S4组, 分别为23个(48.9%)、11个(40.7%)和22个(46.8%)。比较2种聚类结果发现, 两者都能将南方品种聚集在一起, 但农艺及品质性状还能将黄淮海、西北品种聚集在一起。
3 讨论
3.1 青贮玉米杂交品种的品质性状评价及遗传多样性分析
粗蛋白、酸性洗涤纤维、中性洗涤纤维和淀粉是反映青贮玉米品质的重要指标。粗蛋白提供动物的蛋白质和氨化物两大营养物质, 其含量越高, 青贮玉米品质越好。中性洗涤纤维和酸洗洗涤纤维是评价动物消化率的重要指标, 其含量越低, 动物可消化的干物质越多。根据2010年国家颁布的青贮玉米品质分级标准[19], 在本研究2002—2020年通过国家及各省区(市)审定的169个青贮玉米样品数据中,136个样品达到粗蛋白国家一级标准(≥7%), 56个样品达到中性洗涤纤维国家一级标准(≤45%), 86个样品达到酸性洗涤纤维国家一级标准(≤23%), 40个样品达到淀粉含量国家一级标准(≥25%), 且品种质量呈现为逐年提高的趋势。以上表明育种家在青贮玉米的研究方面得到深度和广度拓展, 更加注重饲喂效果, 把为牲畜养殖提供更多能量作为出发,使选育品种的目标更明确, 体现中国培育出的青贮玉米新品种质量越来越好[27]。
青贮玉米品种遗传多样性评价可为新品种选育、品种生产应用提供重要参考。在供试品种的产量及品质性状分析中, 发现干重、粗蛋白和酸性洗涤纤维遗传多样性较丰富。本文研究结果与吴建忠等[17]、吴欣等[28]相比较, 干重、粗蛋白和酸性洗涤纤维的变异系数均高出许多, 这可能与品种数目及种植区域差异大有关。以上研究表明青贮玉米品种的干重、粗蛋白和酸性洗涤纤维存在较大的改良空间, 可为以后优质品种选育提供方向。本研究在王凤格等[29]分析32个青贮玉米品种遗传多样性的基础上将样品数增至141个, 发现等位变异数、基因多样性、PIC、杂合率普遍升高, 但与易红梅等[30]研究参加2014—2019年国家区试的127个青贮玉米品种结果基本一致, 这可能受样品总数大小的影响。
3.2 不同生态区品种遗传多样性
从生态区角度来对青贮玉米进行遗传多样性分析, 能更精准的了解中国各种植区域品种的遗传分化特点。根据方差分析及遗传多样性结果表明, 西北和南方品种基因多样性、杂合度、PIC指标略高于东华北和黄淮海品种, 显现出丰富的遗传多样性,且出现了明显的性状分化现象。分析其可能原因,有如下3点: (1) 生态环境差异。不同生态区的地形、气候、土壤条件等因素影响玉米品种形态性状表达。(2) 生态区地理跨度及需求区域差异。东华北和黄淮海品种种植区域跨度较小, 而西北和南方品种种植区域跨度较大, 且青贮玉米主要需求区域在西北和南方地区, 两地区品种类型多, 故遗传多样性较高。(3) 不同生态区畜牧结构及生产要求不同。西北生态区畜牧业发展迅速, 饲料相对缺乏, 对青贮玉米品种的产量要求较高, 故西北品种生育期及生物产量等性状指标高, 产生性状分化现象。
农艺及品质性状聚类显示, 同一生态区的大部分品种聚集在一起。分析其可能原因为不同生态区的育种目标及生产需求不同, 即不同生态区形成了适应当地需求的育种模式, 对目标性状存在定向选择, 导致品种出现一定的性状分化。除此之外, 该结果体现了我国青贮玉米的品种选育正在逐步出现针对于特点区域、生态区进行品种改良的情况。SSR标记聚类显示, 15个南方品种(75%)聚于S1组, 而其他组群中各生态区品种掺杂, 单生态区品种无明显聚集现象, 结合不同生态区之间的遗传距离, 表明仅南方品种有倾向于聚在一起。分子聚类结果说明,东华北、西北和黄淮海生态区品种基因来源复杂,关键种质资源接近, 不同品种间存在基因交流, 造成亲缘关系接近、遗传差异较小。同时, 南方地区品种与其他生态区品种的遗传分化现象在一定程度上反映出青贮玉米亲本具有区域选择差异。比较两种聚类分析结果发现, 两者均能将大多数南方品种聚集在一起, 但农艺及品质性状还能将黄淮海、西北品种聚在一起。究其原因为这两种分析方法是青贮玉米品种遗传多样性在不同层面上的体现, 形态学聚类方法是依据作物的表型性状来区分不同品种间的遗传差异, 然而表型性状易受很多复杂因素影响如标记数量少、人为测量误差、环境条件等, 造成遗传表达不稳定或不同基因型的品种表现出相同表型特征的结果。相比之下, 分子标记是从DNA水平上反映不同个体间的遗传变异现象, 不受外界环境影响, 结果相对准确。因此, 农艺及品质性状与分子标记的分析结果不完全一致是合理的, 将两类方法相结合能够更加准确、全面的了解物种的遗传变异并描述和解释其遗传背景。
本文利用农艺及品质性状与SSR标记评价了我国4个生态区青贮玉米品种的遗传多样性, 了解了各生态区品种的农艺、品质性状及遗传分化情况,其结果对不同生态区的育种策略调整具有参考价值,但仅从生态区角度对我国主要生产利用的青贮玉米品种进行了分析评价, 今后还有待从统一田间试验方面对品种进行更深层次了解, 以期更加客观评价不同品种间形态差异, 为优质品种推广种植提供科学依据。
4 结论
通过利用农艺、品质性状和SSR标记分析141个国审和各省区(市)青贮玉米品种的遗传多样性,发现我国青贮玉米品种存在丰富的遗传多样性, 尤其是西北和南方品种出现明显的性状分化现象; 南方品种在农艺及品质性状和分子遗传上均具有特异性, 西北和黄淮海品种在农艺及品质性状上具有特异性; 综合两类方法能更准确、全面揭示品种遗传多样性, 其研究结果对不同生态区育种策略调整具有一定参考价值。
