沙苑子苷A对L02细胞脂肪变性模型的降脂作用和机制探讨
2021-07-18孟靓魏益谦高晶李丽邵紫萱孟智睿高学敏王景霞
孟靓 魏益谦 高晶 李丽 邵紫萱 孟智睿 高学敏 王景霞
由于遗传因素、饮食习惯、生活方式和年龄、绝经、疾病及药物等原因引起的血清胆固醇或甘油三酯的水平过高或高密度脂蛋白水平过低,称为血脂代谢异常[1]。脂质代谢异常与许多疾病都密切相关,如动脉粥样硬化[2]、肥胖症[3]、胰岛素抵抗[4]、高脂血症[5]、脂肪肝[6]等,因此对血脂代谢异常的防治有着极为重要的意义。沙苑子(SemenAstragaliComplanati)为豆科植物扁茎黄芪(AstragaluscomplanatusR.Br.)的干燥成熟种子,是中医传统补阳药,功能补肾助阳、滋补肾精[7]。药理研究显示[8-10],沙苑子的总黄酮有明显的调脂作用,能够降低血清甘油三酯(triglyceride,TG)、胆固醇(cholesterol,TC)水平,沙苑子苷A是含量较高的黄酮类成分之一[11],本实验选择沙苑子苷A为研究对象,采用游离脂肪酸(free fatty acid,FFA)诱导的肝脂肪变L02细胞模型,观察沙苑子苷A对肝脂质异常的调节作用,并针对TG的合成途径深入探讨沙苑子苷A的降脂机制。
1 材料与方法
1.1 实验细胞
正常人肝细胞株L02,购于武汉普诺赛生命科技有限公司。
1.2 药物与试剂
沙苑子苷A(美国Apexbio公司,批号:A8425),高糖DMEM培养基(hyclone,批号:sh30284.01)、胰蛋白酶-EDTA消化液(批号:T4299)、青链霉素(批号:V900929),牛血清白蛋白(批号:V900933)、棕榈酸钠(Sodium Palmitate,PA,批号:P9575)、油酸钠(Sodium Oleate,OA,批号:07501)、二甲基亚矾(Dimethyl sulfoxide,DMSO,批号:C6295)、蛋白裂解液(RIPA,批号:r0278)均购自美国Sigma公司,胎牛血清(美国Gibco公司,批号:10099-141)、CCK8试剂盒(日本同仁化学研究所,批号:CK04),油红O细胞染色试剂(北京索莱宝生物公司,批号:g1262),BCA法蛋白质定量检测试剂盒(凯基生物公司,批号:KGP902),TG试剂盒(批号:A110-1-1)、TC试剂盒(批号:A111-1-1)、谷丙转氨酶(glutamic pyruvic transaminase,ALT)试剂盒(批号:C00902-1)、谷草转氨酶(Glutamic oxaloacetic transaminase,AST)试剂盒(批号:C010-2-1)、乳酸脱氢酶(lactate dehydrogenase,LDH)试剂盒(批号:A020-2-2)、丙二醛(Malondialdehyde,MDA)试剂盒(批号:A003-1-2)、超氧化物歧化酶(Superoxide dismutase,SOD)试剂盒(批号:A001-3-2)均来源于南京建成生物工程研究所;实时荧光定量 PCR 检测主要实验材料:引物(北京睿博兴科生物技术有限公司合成),Trizol(美国Invitrogen公司,批号:15596018),RNase Inhibitor(批号:eo0382),RNase-free H2O(批号:en0521)均购自美国Fermentas(MBI)公司,转录酶、qPCR Mix(APEXBIO,批号:k1070-5000),固醇调节元件结合蛋白-1c(sterol regulatory element-binding protein 1C,SREBP-1c)抗体(批号:ab133125)、脂肪酸合成酶(fatty acid synthase,FAS)抗体(批号:EPR5700)、过氧化物酶体增殖物激活受体α(peroxisome proliferator activated receptor α,PPARα)抗体(批号:epr18516)、肉毒碱棕榈酰基转移酶1A(carnitine palmitoyl transferase 1A,CPT-1A)抗体(批号:ab189182)、抗兔IgG HRP辣根过氧化物酶(批号:ab6721)均购自英国Abcam公司。
1.3 主要仪器与设备
超净工作台(青岛海尔股份有限公司),二氧化碳恒温培养箱(上海一恒科学仪器有限公司),高速低温离心机(德国Eppendorf公司),倒置显微镜(德国Leica),全自动生化分析仪(Rayto),酶标仪(Thermo),7500时实定量PCR仪(美国BIO-RAD公司),电泳仪、转膜仪(美国Bio-Rad) ,凝胶成像系统(美国 UVP)。
1.4 脂肪变肝细胞模型的建立
1.4.1 CCK8检测确定FFA浓度范围 将L02细胞铺于96孔板中进行培养,待贴壁后,分别加入不同浓度的FFA(OA∶PA为2∶1),诱导24小时,用CCK8试剂盒测定细胞增殖情况,测出OD值,筛选出FFA浓度范围。
1.4.2 油红O染色的细胞形态观察 油红O染色可以对肝细胞内的脂滴进行定性观察,以0.8×105/mL的密度铺于6孔板中,每孔2 mL,24小时贴壁后加入不同剂量的FFA(OA∶PA为2∶1)作用于L02细胞,培养24小时后,进行染色处理,PBS漂洗2~3遍,4%多聚甲醛固定,光镜下观察细胞内脂滴的数量和大小。
1.4.3 细胞内TG的检测 将L02细胞以1×105/mL种植于24孔板,每孔1 mL,分为对照组和模型组,每组3孔。待细胞贴壁后,加入不同浓度的FFA(OA∶PA为2∶1)干预,培养24小时后,检测TG含量。
1.5 沙苑子苷A对L02 细胞活力的影响
称取沙苑子苷 A 溶于 DMSO 中,获得沙苑子苷 A 储备液。分装后放置于-20℃冰箱保存。临用前用完全培养基稀释,得到沙苑子苷 A 溶液。经过直径为 0.22 μm 的一次性针头滤器,至容积为 15 mL 的经过灭菌处理的离心管中。沙苑子苷 A 溶液终浓度为 1×10-6mM,1×10-5mM,1×10-4mM,1×10-3mM,1×10-2mM,1×10-1mM,1 mM。沙苑子苷 A 溶液中 DMSO 终浓度小于等于0.1%(V/V)。加入贴壁生长的 L02 细胞中,每个浓度设置 3 个重复孔。孵育 24 小时,随后每孔加入 10μL 的 CCK8 试剂,避光室温反应 30 分钟。使用酶标仪 520 nm 波长检测其 OD 值,计算细胞活力百分率。
1.6 分组与给药
用完全培养基(含10%胎牛血清的高糖DMEM)传代培养正常人肝细胞株L02,待细胞呈对数生长是即开始实验。以0.5×105/mL的密度铺板种植,将体外培养的肝细胞分为对照组(Control)、模型组(Model)、沙苑子低、中、高剂量组(S-Low 25 μM、S-Middle 50 μM、S-High 100 μM)。对照组:加入完全培养基;模型组:待细胞贴壁后,加入1 mM FFA(1.4.1中确定的造模浓度)孵育24 小时;沙苑子低、中、高剂量组:细胞贴壁后,沙苑子低、中、高剂量先预处理24小时,再加入1 mM FFA干预24小时。然后测定相关指标。
1.7 指标检测
1.7.1 L02细胞TG、TC、ALT、AST、LDH、MDA含量,SOD活性水平的测定 收集各组细胞,PBS冲洗并1000 r/min离心2~3遍,酶标仪测出相应OD值,同时用BCA蛋白浓度试剂盒检测蛋白含量。严格按照试剂盒说明书,应用全自动生化分析仪检测。
1.7.2 L02细胞炎性因子IL-6、TNF-α的测定 收集各组细胞,ELISA法检测白细胞介素-6(interleukin 6,IL-6)、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)的含量。
1.7.3 实时荧光定量PCR检测SREBP-1c、FAS、PPARα、CTP-1A mRNA表达 提取各组细胞总RNA,根据产品说明书操作。用生物分光光度计测定 RNA浓度和纯度。取 2 μg RNA 样本,用逆转录试剂盒进行逆转录合成 cDNA,并以此为模板进行 PCR 扩增反应。逆转录和 PCR反应条件按照试剂盒说明书设置。PCR进行35~40个循环:预变性 95℃,2分钟;变性95 ℃,30秒;58 ℃退火,30秒 ;72 ℃延伸30秒。以β-actin 为内参,所得 CT 值按照2-△△CT的方法进行均一化处理后再进行统计学分析。引物序列见表1。

表1 引物序列
1.7.4 蛋白免疫印迹法检测SREBP-1c、FAS、PPARα、CTP-1A蛋白表达 收集各组细胞,RIPA裂解液裂解细胞,5000 rpm 离心去除沉淀,BCA 法测定蛋白含量,总蛋白50 μg上样,经10% SDS-聚丙烯酰胺凝胶电泳,电转移至 NC膜上,用含5%脱脂奶粉的TBST封闭后加入1∶1000稀释的相应的一抗,4℃摇床孵育过夜,TBST洗3次,加入1∶2000稀释的辣根过氧化物酶(HRP)标记的二抗,室温孵育1小时,TBST洗3次后,用Western blot荧光检测试剂盒显影、定影。结果用图象分析仪分析。
1.8 统计学方法
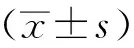
2 结果
2.1 L02细胞脂肪变模型的建立
2.1.1 CCK8法检测不同浓度FFA对L02细胞活性的影响 分别将0.25 mM、0.5 mM、0.75 mM、1 mM、1.5 mM不同浓度FFA干预L02细胞培养24小时后观察细胞存活状况。实验结果显示,FFA浓度在1 mM以下时,细胞死亡漂浮较少。CCK8结果显示,当FFA浓度超过1mM是吸光度显著降低,细胞增殖影响较大(见表2)。

表2 CCK8检测不同浓度的FFA对L02细胞活性的影响
2.1.2 油红O染色观察不同浓度FFA对L02细胞内脂滴形成的影响 光镜下观察可见,对照组细胞边缘完整,胞浆丰富,细胞内未见红色脂滴,实验组随着游离脂肪酸造模浓度的增加,细胞内脂滴呈递增趋势,当FFA浓度大于1mM时,细胞数量明显减少(见图1)。

注:A:对照组;B:0.25 mM剂量组;C:0.50 mM剂量组;D:0.75 mM剂量组;E:1.00 mM剂量组;F:1.5 mM剂量组图1 油红O染色法观察细胞内脂滴(油红O染色,×200)
2.1.3 不同浓度FFA对L02细胞TG含量的影响 当FFA作用浓度为1 mM时,细胞活性及增殖不受大的影响,且在该作用浓度时,细胞内脂滴形成最多、甘油三酯含量最高。而当FFA作用浓度高于1 mM时,细胞的活性与增殖均受到很大影响,且细胞内脂滴与脂质含量减少较多,因此,选择1 mM作为FFA诱导造模的最佳浓度(见表3)。
表3 不同浓度的FFA对L02甘油三酯水平的影响
以上实验结果可知,当FFA作用浓度为1 mM时,细胞活性及增殖不受大的影响,且在该作用浓度时,细胞内脂滴形成最多、甘油三酯含量最高。而当FFA作用浓度高于1 mM时,细胞的活性与增殖均受到很大影响,且细胞内脂滴与脂质含量减少较多,因此,选择1 mM FFA诱导24小时造模成功。
2.2 沙苑子苷A对正常L02细胞活性的影响
CCK-8检测结果显示,当沙苑子苷 A 浓度很小时,低浓度的沙苑子苷 A 对 L02 细胞有增强活性的作用,随着沙苑子苷 A 浓度的升高,细胞活力逐渐与空白对照组相近(见表4)。
表4 CCK8检测不同浓度沙苑子苷A对L02细胞活性的影响
2.3 沙苑子苷A对脂肪变L02细胞TG、TC、ALT、AST、LDH的影响
与空白对照组比较,模型组L02细胞TG、TC、ALT、AST、LDH含量均明显增加,差异有统计学意义(P<0.05);与模型组比较,沙苑子低、中、高剂量组L02细胞TG、ALT、AST、LDH含量均显著降低,差异有统计学意义(P<0.05),沙苑子苷A中、高剂量组L02细胞TC含量均显著降低,差异有统计学意义(P<0.05)(见表5)。
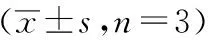
表5 沙苑子苷A对L02细胞内TG、TC、ALT、AST、LDH含量的影响
2.4 沙苑子苷A对脂肪变L02细胞MDA、SOD、IL-6、TNF-α水平的影响
与空白对照组比较,模型组L02细胞MDA、IL-6、TNF-α含量明显增加,差异有统计学意义(P<0.05);与模型组比较,沙苑子苷A中、高剂量组L02细胞MDA含量均显著降低,差异有统计学意义(P<0.05),沙苑子苷A低、中、高剂量组L02细胞IL-6、TNF-α含量均显著降低,差异有统计学意义(P<0.05)。与空白对照组比较,模型组L02细胞SOD活力水平明显降低,差异有统计学意义(P<0.05);沙苑子低、中、高剂量组L02细胞SOD活力水平均显著升高,差异有统计学意义(P<0.05)(见表6)。
表6 沙苑子苷A对L02细胞内MDA、SOD、IL-6、TNF-α含量的影响
2.5 沙苑子苷A对脂肪变L02细胞SPEBP-1c、FAS、PPARα、CPT-1A mRNA表达水平的影响
与对照组比较,模型组L02细胞SPEBP-1c、FAS mRNA的表达均明显升高,差异有统计学意义(P<0.05),PPARα mRNA的表达有升高趋势,但是无统计学意义,CTP-1A mRNA的表达显著降低,差异有统计学意义(P<0.05);与模型组比较,沙苑子苷A低、中、高剂量L02细胞FAS、SPEBP-1c mRNA的表达均显著降低,差异有统计学意义(P<0.05),沙苑子中、高剂量组L02细胞PPARα、CPT-1A mRNA的表达显著升高,差异有统计学意义(P<0.05)(见表7)。
表7 沙苑子苷A对L02细胞SPEBP-1c、FAS、PPARα、CTP-1α mRNA的影响
2.6 沙苑子苷A对脂肪变L02细胞SREBP-1c、FAS、PPARα、CPT-1A蛋白表达水平的影响
与对照组比较,模型组L02细胞SREBP-1c、FAS蛋白表达均明显升高,差异有统计学意义(P<0.05),PPARα蛋白的表达有升高趋势,但是无统计学意义,CPT-1A蛋白的表达显著降低,差异有统计学意义(P<0.05);与模型组比较,沙苑子苷A低、中、高剂量L02细胞SREBP-1c、FAS蛋白的表达均显著降低,差异有统计学意义(P<0.05),沙苑子中、高剂量组L02细胞PPARα、CPT-1A蛋白的表达显著升高,差异有统计学意义(P<0.05)(见表8,图2)。
表8 沙苑子苷A对L02细胞SREBP-1c、FAS、PPARα、CPT-1A蛋白的影响

注:A:空白对照组;B:1mM游离脂肪酸;C:沙苑子苷A低剂量组;D:沙苑子苷A中剂量组;E:沙苑子苷A高剂量组图2 甘油三酯代谢相关蛋白免疫印迹图
3 讨论
目前,西医常用的调脂药有他汀类、贝特类、胆酸螯合树脂类、烟酸类及其衍生物等,但都有一定的胃肠道不良反应、头痛或者肌肉疼痛,严重的可出现肝功能损害或横纹肌溶解症[12]。黄酮类化合物是一种广泛存在于自然界中的天然有机化合物,具有较高的安全性和广泛的生物活性[13],越来越多的研究显示黄酮类化合物在调节血脂和预防冠心病方面发挥了重要作用[14],如:罗布麻叶总黄酮能减轻高血脂导致的血管内皮损伤和改善体内氧化应激水平[15],山楂叶总黄酮可调控法尼醇受体(farnesoid X receptor,FXR)/ SREBP-1c通路改善非酒精性脂肪肝大鼠肝脏脂质代谢紊乱[16],还有沙棘总黄酮[17],银杏叶总黄酮[18]等等,具有良好的开发和应用前景。沙苑子黄酮是沙苑子中含量较高的一类化合物,目前已经从沙苑子中提取到超过16 种的黄酮类化合物,其中沙苑子苷A是含量较高的黄酮类成分之一[11,19]。目前国内外学者对沙苑子总黄酮的药理作用研究较多,但是对单体成分的药理研究还未见报道。本实验采用FFA诱导 L02 构建肝细胞脂肪变模型,观察沙苑子苷A对脂肪变L02细胞的影响,探讨其降脂作用及分子机制。
肝脏脂肪变的细胞模型较高脂饲养建立的动物模型个体差异较小,可较好的对实验条件进行把控,其中油酸和棕榈酸以2∶1比例而成的FFA混合物建立细胞脂肪变模型更符合人体脂肪变的病理过程[20]。本实验采用CCK8法测定了不同浓度FFA对细胞活性的影响,同时结合油红O染色光镜下细胞内脂滴形成情况和定量检测细胞内TG水平,最终确定1 mM FFA诱导24小时可成功造模。沙苑子苷A干预治疗后,能显著降低L02细胞TG、TC、ALT、AST、LDH含量,同时降低MDA含量、提高SOD活性,降低IL-6、TNF-α的水平,说明沙苑子苷A能减轻脂毒性造成的肝细胞损伤,缓解氧化应激状态和炎症反应,提示沙苑子苷A是沙苑子降脂保肝的有效单体成分。
研究表明,TG合成途径中两个重要的核受体转录因子SPEBP-1c和PPARα对其起关键作用。SREBP-1c是脂肪酸和甘油三酯合成中的关键分解因子,主要参与肝脏脂肪酸合成和脂肪前体细胞的分化,其过表达会引起与脂质异常相关的疾病如糖尿病、高脂血症、代谢综合征的发生[21-22],可通过调节其下游靶基因FAS、ACC、GPAT的表达来调节肝脏中TG的合成[23]。其中乙酰辅酶A羧化酶(ACC)催化乙酰辅酶A形成丙二酰辅酶A,合成长链脂肪酸前体,脂肪酸合成酶(FAS)是一种多功能复合酶,催化乙酰辅酶A和丙二酰辅酶A,参与长链脂肪酸生产的内源性蛋白酶[24],在合成脂肪酸方面发挥着关键的作用[25]。GPAT是TG生物合成的限速酶[26]。本实验结果显示,给予沙苑子苷A预处理,模型细胞SREBP-1c、FAS mRNA和蛋白的表达量均显著,表明沙苑子苷A可通过抑制肝脏SREBP-1c的表达,下调肝脏FAS,抑制肝脏中TG的合成。
PPARα是核受体超家族的成员之一,可显著改善血脂异常和减轻动脉粥样硬化相关疾病的风险[27]。PPARα激活后与配体形成异二聚体[27],可调控与脂质代谢相关的多种基因编码的蛋白质,如脂肪酸转位酶、乙酰辅酶A氧化酶等[28],从而介导TG和脂肪酸的代谢分解。CPT-1A是线粒体内脂肪酸β氧化途径的限速酶[29],其作用是将长链脂肪酸从胞浆转运如线粒体内部,进行β氧化[30]。本实验结果显示,给予沙苑子苷A预处理,模型细胞PPARα、CPT-1A mRNA和蛋白表达量均显著升高,说明沙苑子苷A通过上调肝脏PPARα的表达水平,提高CPT-1A的活性,加速了脂肪酸的β氧化分解,进而使TG的合成减少。
综上所述,沙苑子苷A对脂肪变性L02肝细胞模型具有良好的降脂和保肝作用,其降脂机制可能是沙苑子苷A通过抑制肝细胞SREBP-1c的表达,降低TG合成途径中限速酶FAS水平,降低TG合成速度;同时上调PPARα蛋白表达,提高脂肪酸β氧化途径中CPT-1A水平,加速脂肪酸的β氧化,减少TG合成,从两方面共同作用从而达到降脂和保肝效果。
