动物鼠疫周期流行的生态位影响因素研究进展
2021-07-16秦婧靓杨瑞馥崔玉军
秦婧靓,杨瑞馥,崔玉军
传染病是病原微生物或寄生虫感染人体后产生的具有传染性的疾病,可在人群中传播与流行[1]。地理、气候、社会因素等对传染病的流行及静息均可产生影响。若该传染病在当地一直处于停息状态但没有被根除,认为其处于静息期(latent enzootic phase);若该病发病率显著高于历年一般水平时,则认为该病处于流行期(epizootic phase)[1]。鼠疫(Plague)是我国法定的甲类传染病,其发病急、传染性强,治疗不及时病死率高,不依赖于人类而独立存在于自然疫源地中[2]。在鼠疫自然疫源地中,可观测到鼠疫存在周期性流行,即流行期与静息期发生交替的现象。但是交替周期并不固定,其中静息期的时间跨度可长达数十年或更久。如:我国广西鼠疫静息53年后,于2000年7月在隆林县天生桥库区及周边发生动物间疫情,并波及人间[3-4]。云南省动物间鼠疫于1982年在滇西复燃,随后滇南、滇中、滇东也出现疫情。其中普洱、思茅、石屏、宜良县鼠疫疫源地分别静息72年、93年、100年、134年后又开始活跃并出现人间疫情[5]。
生态位(niche)是有机体种群对所生存环境条件的耐受性和生境资源需求的综合。鼠疫生态位包括疫源地的环境、宿主、媒介等因素[6]。值得注意的是:鼠疫耶尔森菌(Yersiniapestis,简称鼠疫菌)本身也是鼠疫生态位中重要一环。鼠疫菌会通过对媒介/宿主的影响,改变生态位的整体平衡。在适宜生态条件下,鼠疫菌、宿主动物、媒介生物的种群数量达到动态平衡,鼠疫菌可进行稳定的定植和传播,导致鼠疫在动物间流行。而当生态位的动态平衡被破坏时,动物间鼠疫就可能静息或者随宿主、媒介迁徙溢出到其他地区[7]。现有研究表明动物间鼠疫周期性流行特征的出现与鼠疫自然疫源地生态位的动态变化密切相关[8-9]。例如中世纪鼠疫大流行时,欧洲大部分地区存在鼠疫疫情,但啮齿类动物种群改变[10];居民生活水平提高[10-11];动物栖息地丧失和工业化发展[12]等因素破坏原生态位的动态平衡,则使欧洲鼠疫菌灭绝,疫源地消失[13]。本文将从影响鼠疫自然疫源地生态位稳定的主要因素入手,对宿主动物、蚤类媒介、土壤、鼠疫菌多样性及人类活动5个方面的相关研究进行梳理总结(图1、表1)。

表1 影响动物间周期性鼠疫流行的主要因素Tab.1 The key factors affecting periodic plague epizootics
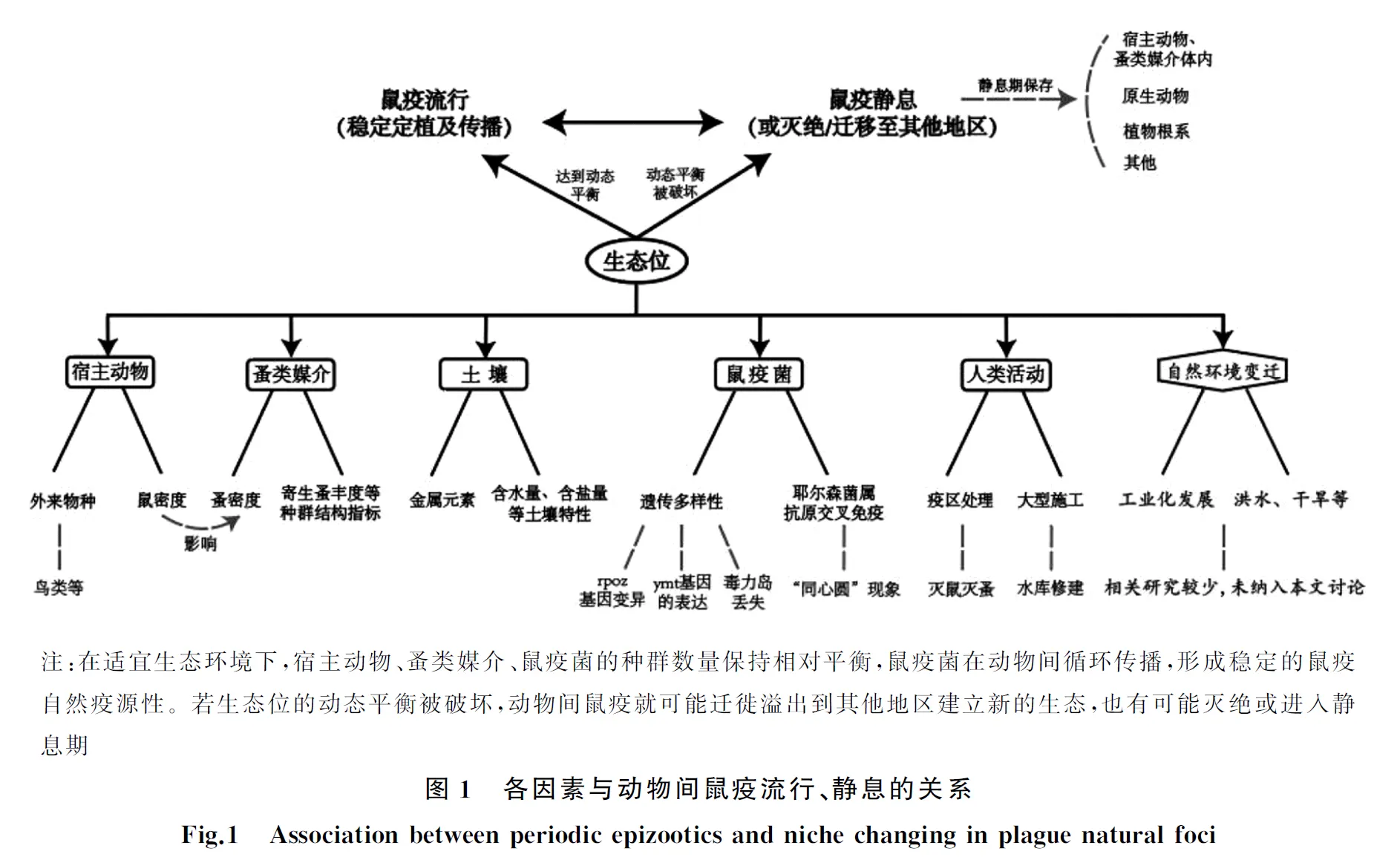
1 宿主动物与生态位
目前,世界上共发现2 369种啮齿类动物[14], 截至2017年8月底,我国记录在册啮齿目达到9科78属220种[15],染疫啮齿类动物53种[16],传播鼠疫菌的主要宿主动物(鼠类)有14种[17],分布于我国北部、西北部、西南部及东南沿海地区[18]。啮齿类动物作为鼠疫菌的主要宿主,对维持鼠疫自然疫源地生态位的动态平衡具有重要作用。我国鼠疫历年监测数据显示,动物间鼠疫流行及静息与该疫源地主要宿主动物的密度有直接的关联。研究人员通过对甘宁阿拉善黄鼠疫源地50年间鼠疫疫情进行分析,发现该疫源地发生的3次动物间鼠疫流行期的年均鼠密度显著高于静息期平均值,流行期当地黄鼠年均密度均高于1只/hm2[19]。高少坤等[20]发现河北省鼠疫自然疫源地1993-1995年、2001-2005年宿主动物密度明显高于1981-2013年的平均水平,使鼠疫在动物间流行可能性增大,影响了鼠疫疫情的发生和发展。也有学者认为外来物种介入会对疫源地生态位的相对平衡产生影响,如:被鼠疫感染的鸟类或其他物种进入处于静息期或鼠疫非流行地区时,会引起该疫源地鼠疫再次流行[21]。
2 媒介生物与生态位
据统计,全世界已发现蚤类2 500余种和亚种,分别隶属于5总科16科238属。我国已发现4总科10科75属655种和亚种[22],传播鼠疫菌的主要媒介昆虫(蚤类)有28种[23]。蚤类作为鼠疫菌传播的主要媒介及储存宿主,主要通过在不同宿主间吸血传播鼠疫菌。蚤类密度增高不仅会增加对宿主的叮咬次数,而且提高了鼠疫菌在宿主间的传播概率,对疫源地内动物间鼠疫流行和鼠疫菌延续起重要作用[24]。在漫长的生物进化中,蚤类与宿主形成寄生或共生关系,蚤类数量的变化往往又受宿主数量的影响。前期研究表明:体蚤指数与鼠密度呈正相关关系,鼠密度增加则体蚤指数相应增高[25]。若生态环境改变,疫源地的主要宿主动物随之变化,也间接影响了相应的主要寄生蚤数量及分布。此外,鼠疫生态系统的稳定还与蚤类群落其他结构指标密切相关。叶超等[26]研究发现,准噶尔盆地大沙鼠疫源地内寄生蚤对大沙鼠的侵染能力(染蚤率)和寄生数量(总蚤指数)的季节性变化与该地鼠疫季节性的流行特征一致。与此同时,蚤类群落的多样性和丰富度增高、优势度降低。这些结构指标的变化有利于维持疫源地生态位的动态平衡。
3 土壤与生态位
土壤是微生物的聚集地,它与宿主、媒介构成的小型生态系统共同维持鼠疫菌生命的延续[27]。鼠疫菌能够在土壤中生存,其与含盐量、含水量、金属元素等土壤特性息息相关。相对湿润、中等盐碱的土壤不仅为微生物和地表动物的生存和繁殖提供了有利条件,而且益于鼠疫菌在疫源地的长期保存[28]。此外,土壤中的金属元素(例如铁、钙)可参与调节自然界中动物感染鼠疫的过程[29],促进细菌生长,增强其毒力[30],提高鼠疫菌感染宿主的能力[31]。但金属元素过量或缺乏都会影响细菌的生存和繁殖,鼠疫菌对各类金属元素的摄取应受到精确调控[32]。当疫源地内土壤条件适宜、微生物种类丰富、金属离子浓度相对平衡时,有利于宿主或鼠疫菌生存,可能促进鼠疫发生或流行。而暴雨洪涝等极端气候可使土壤盐度、含水量、金属离子浓度发生变化,有可能导致鼠疫疫源地局部灭绝或进入静息期[33]。
4 “外延的表型”:鼠疫菌遗传变异与生态位
“外延的表型”(extended phenotype)是理查德道金斯在1982年提出的概念,强调基因作为被选择的基本单元,可在生物体外产生表型效应(通过基因组变异造成个体周围环境改变)[34],并且能够延伸至更广泛的环境或其他个体中。例如鸟巢就是鸟类基因组通过某种形式所编码的具体实物。我们的研究表明,细菌中也存在类似现象[35]。因此,鼠疫菌遗传多样性能够通过“外延的表型”机制,与生态位发生作用。
4.1 鼠疫菌基因组多样性对生态位的影响和关联 鼠疫菌可以在蚤类前胃中形成生物膜,从而阻碍血液进入蚤类消化道,使其一直处于饥饿状态,吸血行为更为频繁,最终促进鼠疫菌的快速播散[36-37],其可通过遗传变异调节与蚤体内生物膜形成相关的表型,从而改变鼠疫菌经蚤传播能力,以适应生态环境选择压力[35]。rpoZ基因主要编码RNA聚合酶ω亚基,与鼠疫菌的生长速率和生物膜的形成能力有关[38]。在新疆天山古尔图鼠疫自然疫源地的研究发现:该地区秋冬季发生异常寒冷、干旱时,鼠疫菌rpoZ基因上可观测到很强的正向选择信号。体外实验表明该基因突变会导致鼠疫菌生物膜形成能力变化,从而将鼠疫菌基因变异与蚤类媒介吸血行为改变关联起来[35]。该研究提示鼠疫菌基因组变化可以通过媒介等级联作用,最终影响鼠疫流行强度。
绝大多数鼠疫菌携带3个质粒。其中pMT1质粒上的鼠毒素基因ymt(磷脂酶D)对于鼠疫菌在蚤肠道中的正常存活必不可少。它不仅可以保护鼠疫菌免受跳蚤肠道内含物的细胞毒性影响,而且有利于其在节肢动物肠道中的定殖[39]。ymt活性依赖于生物下调因子rcsA、PDE2(rtn)、PDE3(y3389)[40]和ureD[41]等基因的假基因化,假基因的形成增强了鼠疫菌生物膜的形成能力,以及经蚤传播的能力[42]。而这些基因所参与的调控子自身表达(如RcsAB)往往又受气候/环境信号因子(pH值、温度、代谢物水平、离子浓度)[43]等的调控。适宜的温度(26 ℃)可使ymt基因的表达量增高[44],利于鼠疫菌的定殖及传播,从而促进鼠疫流行。
鼠疫菌在自然条件下或传代过程中,有可能获得或缺失部分质粒或毒力岛,对宿主动物的侵袭能力产生影响[43-44]。以pgm位点为例:该位点是鼠疫菌染色体上一段长度为102 kb的强毒力岛[45],两侧存在同向重复的插入元件IS100,可介导鼠疫菌在传代中发生同源重组及序列缺失[46]。pgm位点的色素沉着区与pgm阳性表型相关,该表型在动物间传播过程中起主导作用[47];而其携带的hmsHFRS操纵子是生物膜在蚤体内形成的关键基因[48],该基因的转录有利于鼠疫菌生物膜的形成,对跳蚤传播鼠疫十分重要[49]。现有研究表明鼠疫菌pgm位点在实验室及自然界传代过程中均可发生缺失[47],其缺失形成的种群遗传多样性将对鼠疫菌在媒介和宿主间的传播造成影响。
4.2 耶尔森菌属抗原交叉免疫对生态位的影响 除了鼠疫菌自身外,耶尔森菌属其他近缘物种的抗原交叉免疫也可能对生态位产生影响。假结核耶尔森菌(Y.pseudotuberoulosis,假结核菌)与小肠结肠炎耶尔森菌(Y.enterocolitica)与鼠疫菌均属于耶尔森菌属。三者的基因组分析表明,其在遗传水平上亲缘关系很近[50],均含有共同携带的毒力质粒(pYV,在鼠疫菌中被称为pCD1)编码的V抗原(LcrV)及外膜蛋白A(outer membrance protein A,OmpA)等多组一致的抗原结构[51]。因而疫源地动物在感染某种致病性耶尔森菌后可能获得保护性免疫。Harrison等[52]发现,鼠疫菌在自然疫源地中通常与假结核菌、小肠结肠炎耶尔森菌交替出现,随后鼠疫疫情由流行转为静息。高少坤和杜国义[50]比较了鼠疫菌与假结核菌的基因片段和历史上曾发生的3次世界范围的人间鼠疫大流行,也发现鼠疫菌与假结核菌在鼠疫流行和静息期曾交互出现。同一疫源地内动物感染鼠疫菌、假结核菌或小肠结肠炎耶尔森菌的分布此消彼长呈“同心圆”现象(concentric circle distribution)[53]。这一现象间接解释了在鼠疫自然疫源地的啮齿类动物中通常无法分离出其他致病性耶尔森菌的现象;反之也提示耶尔森菌属其他物种的入侵可能是引起生态位变化、疫源地静息的原因之一。
5 人类活动与生态位
人类活动也会引起动物间鼠疫疫情流行及静息的交替转换,比如以灭鼠、灭蚤为主的疫区处理工作,会造成主要宿主动物及蚤类媒介的密度急剧降低。由于疫源地内宿主动物恢复慢,减缓了动物间鼠疫传播速度,短期内无法发生大规模动物间鼠疫疫情,鼠疫趋于静息[54]。当疫源地经过多年的自身调节,宿主动物逐渐恢复,达到一定程度后会再次引起鼠疫流行。若动物间鼠疫流行时没有人为干预(疫区处理),鼠疫可能会波及至人导致人间鼠疫的流行[20]。除此之外,水库修建等大型施工活动也会破坏鼠疫的生态平衡,我国广西南盘江流域就曾因修建天生桥水库而引起动物及人间鼠疫的流行。在建设水库的十年间,当地未出现过动物间疫情。但随着水库的蓄水将周围农田和植被淹没,使得原本分散的啮齿类动物被集中在一个没有水的地方,鼠疫菌经跳蚤叮咬人后最终引发人间疫情。随着水库成功修建以及人类居住环境和动物、植被的逐渐恢复,鼠疫自然疫源地转入静息期[3,9]。
6 鼠疫静息机理的认识
鼠疫菌是生态位中关键的一环,因此,了解其在静息期内如何保存,又以何种机制引起动物间流行对于鼠疫研究而言十分重要。鼠疫菌不仅可以在动物及媒介体内保存,还能在土壤、植物等生态因子中呈非寄生相保存。原生动物是许多病原体的宿主和传播者[55],其对所吞噬的细菌有选择作用,研究证实鼠疫菌也可寄生在土壤原生动物中[56]。在冬季或干旱等不利环境下,原生动物通过吞噬土壤中的鼠疫菌形成孢囊,以“休眠”状态贮藏,保护细菌不受外界环境侵害[56-57]。有些阿米巴个体吞噬鼠疫菌后,被吞噬细菌可在其细胞内大量繁殖,最终使细胞破裂而释放到外环境中[58]。宋志忠等[59]提出鼠疫菌还可能以植物根系正常菌群的形式保存在根系环境中。当环境内某些因素发生变化时,细菌可通过根部进入植物地上部分,通过摄食途径感染啮齿类动物,造成动物间鼠疫的传播[59-60],但这一推论目前尚缺乏强有力的实验室和自然条件证据。
7 结 语
动物间鼠疫流行、静息期交替转换的现象普遍存在于各鼠疫自然疫源地中。除上文总结的影响因素之外,自然灾害、气候变化等也会对当地动物及媒介生物的栖息环境产生影响。适宜的生态环境使得啮齿类动物和媒介种群扩增,为鼠疫在动物间的定植、传播提供机会。而洪涝、干旱、地震等自然灾害的发生能短时间内破坏生态位的动态平衡,对鼠疫自然疫源地造成影响,鼠疫可能局部灭绝或进入静息期,也可能迁徙溢出到其他地区建立新的鼠疫生态位[7-8]。此外,气温的季节性变化与宿主动物的活动强度、生殖腺发育、受孕和繁殖等密切相关。温度过高或过低都能使动物活动强度降低,出现发育迟缓等现象[61-62],间接对鼠疫疫情造成干扰。
尽管动物间鼠疫流行和静息期转换研究一直是自然疫源性传染病领域的重要课题,但目前已知的鼠疫生态位影响因素依然有限,鼠疫流行周期性转化规律和详细机制仍不明确。我国作为鼠疫自然疫源地分布和类型最为丰富的国家之一,在面临鼠疫防控巨大挑战的同时,也拥有独特的研究资源。随着生态学、生物组学、生物信息学等数据密集型学科的飞速发展,将会为鼠疫流行与静息转换机制的研究方法和策略带来新的思路。尤其是将生态学与进化理论(Eco-Evo)相结合的先进研究方法,将使我们从多个维度更加充分的认识鼠疫,进一步揭示鼠疫周期性流行的转换机制,提高预测预警能力,为鼠疫防控工作提供支撑。
利益冲突:无
