不同栽培基质配方对银耳胞外酶活性的影响
2021-04-19马运龙林占熺陈超前黄暖云林兴生
马运龙 林占熺 陈超前 黄暖云 林兴生

摘 要:为探明不同栽培基质配方对银耳胞外酶活性的影响,以棉籽壳和木屑原料为对照,利用鲜绿洲1号、鲜巨菌草等菌草为主要原料工厂化栽培银耳,比较不同配方银耳不同生長期胞外酶活性。结果表明:银耳在不同栽培基质下羧甲基纤维素酶、滤纸纤维素酶、淀粉酶、漆酶和木聚糖酶等胞外酶活力大小为棉籽壳(C1)>鲜绿洲1号(L1)>鲜巨菌草(J1)>木屑(C2),以鲜菌草为原料工厂化栽培银耳的营养基质利用能力强于木屑,鲜菌草可以替代木屑作为原料工厂化栽培银耳。
关键词:银耳;鲜菌草;胞外酶
中图分类号:S 646.14 文献标志码:A 文章编号:0253-2301(2021)11-0023-05
DOI: 10.13651/j.cnki.fjnykj.2021.11.004
Effects of Different Cultivation Substrate Formulations on theExtracellular Enzyme Activities of Tremella Fuciformis
MA Yun-long1, LIN Zhan-xi1, CHEN Chao-qian1, HUANG Nuan-yun2, LIN Xing-sheng1*
(1. National Engineering Research Center of JUNCAO Technology, Fujian Agriculture and
Forestry University, Fuzhou, Fujian 350002, China; 2. Fujian Xiangyun Biotechnology
Development Co., Ltd., Sanming, Fujian 365106, China)
Abstract: In order to explore the effects of different cultivation substrate formulations on the activities of extracellular enzymes of Tremella fuciformis, the cottonseed shell and sawdust were used as the control, and the fungi such as fresh Lvzhou No.1 and fresh Pennisetum giganteum were used as the main raw materials for the industrial cultivation of Tremella fuciformis, thus to compare the activities of extracellular enzymes of Tremella fuciformis in different growth stages. The results showed that the activities of extracellular enzymes such as carboxymethyl cellulase, filter paper cellulase, amylase, laccase and xylanase under different cultivation substrates of Tremella fuciformis were as follows: cottonseed shell (C1) > fresh Lvzhou No.1 (L1) > fresh Pennisetum giganteum (J1) > sawdust (C2). The utilization ability of nutrient matrix in the industrial cultivation of Tremella fuciformis by using fresh fungi as the raw material was stronger than that of sawdust, and fresh fungi could replace sawdust as raw material for the industrial cultivation of Tremella fuciformis.
Key words: Tremella fuciformis; Fresh fungi; Extracellular enzyme
银耳Tremella fuciformis Berk,是一种食药同源的大型真菌[1-2],可段木栽培和代料栽培。以棉籽壳为原料栽培银耳,虽然产量高,其质量不如段木或木屑银耳[3-5],可能产生农药残留[6]及棉酚危害[7-9]等问题,导致银耳价格较低;以木屑为原料栽培银耳,品质较好,产量比棉籽壳栽培的低,但由于林木的再生能力弱,多地限制砍伐树木用于栽培菌类,严重制约了银耳产业的可持续发展[5]。1986年福建农林大学发明了菌草技术[10],利用五节芒、芦苇、巨菌草、类芦等干菌草栽培食药用菌,并初步探索出菌草栽培银耳技术[11-13]。传统食用菌理论认为,腐生菌类所需的营养物质来自已经死亡的及无生活力的有机物,只能从枯死的木本和草本植物中吸收营养并形成子实体,活性植物细胞具有抗性,且含水量高达90%以上,菌丝无法生长,原料干燥目的是杀死植物细胞,使其生理含水量降低。本研究旨在探讨利用未经干燥的鲜菌草栽培银耳[1,4]。
银耳的生长离不开对栽培基质中营养物质的利用,木质纤维素、淀粉等物质是食用菌生长的主要能量来源[14],但它们不能直接被菌丝吸收利用。食用菌通过分泌胞外酶,把木质纤维素分解成可被菌丝吸收的小分子碳水化合物,进而为菌丝的生长提供能量[15]。胞外酶的活力可以反映出菌丝在不同时期对栽培基质的利用情况,本研究以棉籽壳和木屑原料作为对照,以鲜绿洲1号Arundo Donax Lvzhou No.1、鲜巨菌草Pennisetumgiganteumz.x.lin为主要原料工厂化栽培银耳,通过比较不同配方工厂化栽培银耳的胞外酶在不同时期的变化规律,分析银耳生长过程中对不同栽培基质的利用情况,了解鲜菌草配方与棉籽壳或木屑配方对营养物质利用的差异,为鲜菌草工厂化栽培银耳配方优化及其生产工艺提供依据。
1 材料与方法
1.1 供试菌株
银耳XY04(A4)菌株,由福建省祥云生物科技发展有限公司提供。
1.2 栽培配方
L1:鲜绿洲1号54%,棕榈仁粕20%,莲子壳10%,麦麸14%,石膏1%,石灰1%;J1:鲜巨菌草54%,棕榈仁粕20%,莲子壳10%,麦麸14%,石膏1%,石灰1%;C1:棉籽壳71%,麦麸28%,石膏1%;C2:木屑71%,麦麸28%,石膏1%。
按比例称取原料,投入搅拌机中加入适量水,搅拌40 min,控制含水率在58%左右,拌匀后装瓶,每瓶装料560 g,打孔封口,于灭菌锅中灭菌6 h,移入无菌房,冷却后接种。
1.3 工厂化栽培条件
接种后的栽培瓶移入养菌房中,保持温度20~23℃,相对湿度80%~85%,25 d后,栽培瓶开盖,移入出菇房中,控制温度23~25℃,相对湿度80%,待耳片舒展时湿度控制在90%,于采收前5 d降低室内湿度,42 d时进行采收。
1.4 样品收集
接种后,各配方每6 d取样一次,每次取3瓶作为生物学重复,共取样7次。取出栽培料后,搓碎拌匀,取100 g栽培料,用液氮速冻后于-80℃冰箱中保存备用。
1.5 胞外酶活性测定
1.5.1 粗酶液制备 称取10 g解冻后的栽培料,研磨后移入三角瓶中,加入50 mL 蒸馏水,于25℃恒温振荡培养箱中150 r·min-1浸提90 min,浸提液用双层纱布进行过滤,滤液用蒸馏水定容至50 mL,9000 r·min-1离心15 min,上清液即为粗酶液。
1.5.2 羧甲基纤维素酶活性测定 向试管中依次加入预热的粗酶液0.5 mL和1% C8H11O5Na溶液1 mL,以煮沸灭活的粗酶液为对照组,于50℃水浴30 min。迅速加入DNS溶液1.5 mL,沸水浴反应7 min,立即用流水冷却,用蒸馏水定容至10 mL,于540 nm处测定吸光度值。酶活力单位的定义为:在50℃、pH 4.5的条件下,1 min水解底物生成1 μg葡萄糖为1个酶活力单位
U[16]。
1.5.3 滤纸纤维素酶活性测定 向试管中加入预热的粗酶液0.5 mL和CH3COONa缓冲液(pH 4.5)1 mL,再加入(50±0.5)mg滤纸1条(1 cm×6 cm),以高温灭活的粗酶液为对照组,于50℃水浴1 h。立即加入DNS溶液1.5 mL,沸水浴反应7 min,立即用流水冷却,用蒸馏水定容至10 mL,于540 nm处测定吸光度值。酶活力单位的定义为:在50℃,pH 4.5条件下,1 mL酶液1 min水解底物生成1 μg葡萄糖为1个酶活力单位U[17]。
1.5.4 淀粉酶活力测定 取可溶性淀粉溶液4 mL于试管中,加入磷酸缓冲液1 mL,再加入0.5 mL粗酶液,以煮沸灭活的粗酶液为对照组,于50℃水浴30 min。取反应液1 mL,迅速加入盛有0.5 mL稀盐酸和5 mL稀碘液的试管中,于660 nm处测定吸光度值。酶活力单位的定义为:在60℃、pH 6.0条件下,1 min水解1 μg可溶性淀粉,定义为1个淀粉酶活力单位U[18]。
1.5.5 漆酶活力测定 取1 mmol·L-1 ABTS溶液0.1 mL于96孔板中,加入CH3COONa缓冲液(pH 4.5)0.15 mL,于30℃预热5 min,加入粗酶液50 μL,立即记录吸光度值,每隔30 s记录1次,共记录3 min。酶活力单位的定义为:在30℃、pH 4.5条件下1 min氧化1 μmolABTS所需酶量定义为1个酶活力单位U[19-20]。
1.5.6 木聚糖酶活力测定 取1% 木聚糖溶液0.75 mL于试管中,加入粗酶液0.25 mL,以煮沸灭活的粗酶液作为对照组,50℃保温30 min,立即向试管中加入DNS試剂1.5 mL,沸水浴7 min,流水冷却至室温,用蒸馏水定容至10 mL,于540 nm处测定吸光度值。酶活力单位的定义为:在50℃、pH 5.2条件下,1 min水解底物生成1 μg木糖所需的酶量定义为1个酶活力单位U[21]。
1.6 数据处理
使用SPSS 25软件进行数据分析,使用Origin 2021软件作图。
2 结果与分析
2.1 羧甲基纤维素酶活力变化
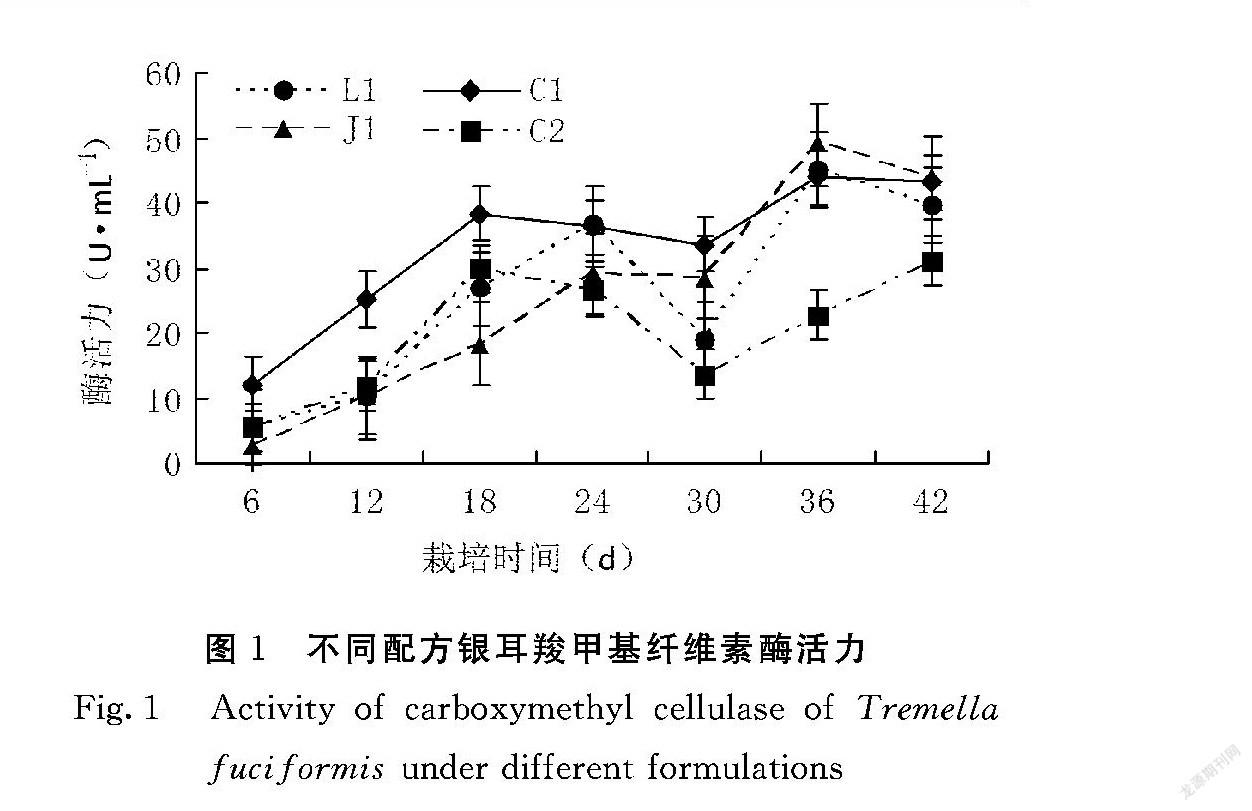
羧甲基纤维素酶是纤维素降解酶系的代表之一,该酶活力的变化能反映出银耳对栽培基质中纤维素的利用情况。由图1可知,不同配方随着栽培时间的增加,羧甲基纤维素酶活性呈现出先升高,后降低,再升高的过程,其中,第1次峰值,L1和J1出现在第24 d,且L1显著高于J1,C1和C2出现在第18 d,C1显著高于C2,但它们均在第30 d下降至最低值,C1显著高于J1,J1显著高于L1,L1显著高于C2。结合银耳的生长特性分析,在第24 d至30 d羧甲基纤维素酶活力的降低可能与银耳原基形成有关,在第30 d之后,子实体的生长需要消耗大量营养物质,因此酶活力迅速升高,为子实体提供足够的养分,从原基形成(第24 d)至子实体成熟(第42 d),C1、 L1 、J1的羧甲基纤维素酶活力均显著高于C2。
2.2 滤纸纤维素酶活力变化
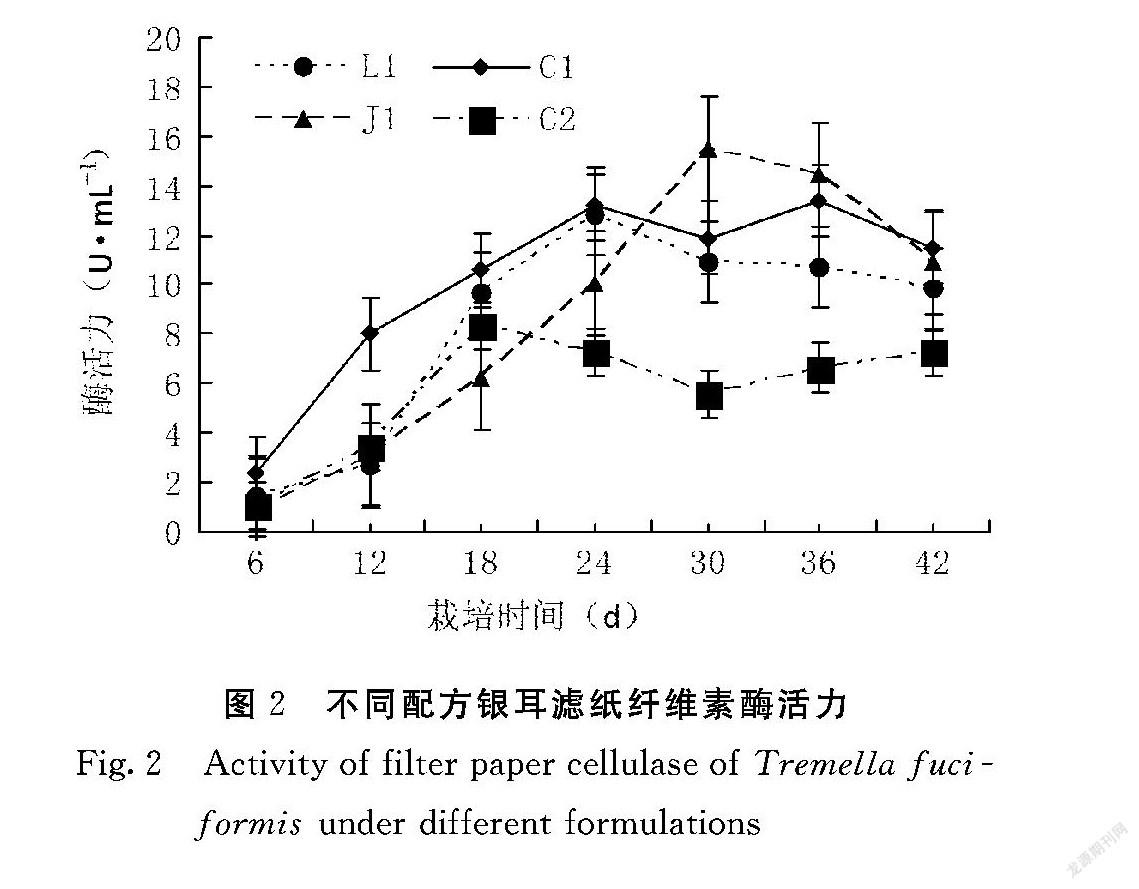
滤纸纤维素酶活力也是反应纤维素降解酶系的重要标志。由图2可知,滤纸纤维素酶活力随着栽培时间的延长,总体上呈现上升之后保持平稳的趋势。在菌丝生长第12 d,C1的酶活力显著高于其他配方,L1、J1、C2差异不显著。C2的滤纸纤维素酶活力在24 d后低于其他配方且差异显著,J1的酶活力波动较大,其他配方酶活力升高之后较为平稳。L1的酶活力均低于C1,但变化趋势与之相同。综合羧甲基纤维素酶和滤纸纤维素酶的数据来看,C1、L1、J1的纤维素酶活力均高于C2。
2.3 淀粉酶活力变化
淀粉酶可以分解栽培基质中的淀粉等多糖类物质,使其水解成可供菌丝吸收利用的单糖等小分子物质,是菌丝生长过程中参与碳水化合物代谢的关键酶之一。由图3可知,随着栽培时间的延长,L1和J1的淀粉酶活力逐渐升高,至子实体成熟期开始达到稳定状态,而C1和C2则是先升高后缓慢降低,在原基形成阶段达到峰值。在菌丝生长期,C1的淀粉酶活力显著高于其他配方,C2次之,在第30 d,不同配方淀粉酶活力无显著差异,在子实体生长后期,L1和J1的酶活力较接近,且显著高于C1和C2。菌草配方L1和J1的淀粉酶活力在菌丝生长和原基形成期(30 d前)低于棉籽壳和木屑配方,而在子实体生长期,菌草配方的淀粉酶活力继续升高,显著高于棉籽壳和木屑配方,并保持平稳。
2.4 漆酶活力变化
漆酶是一种多酚氧化酶,在栽培料中主要降解木质素。由图4可知,4个配方的漆酶活力均呈现出先升高后降低的趋势,但达到峰值的时间不同。C1和C2在菌丝生长期的活力高于L1和J1,在第24 d达到峰值,之后迅速下降,而L1和J1则分别在第30 d和第36 d达到峰值,且下降较缓慢。在菌丝生长24 d,漆酶活力C1显著高于C2,C2显著高于L1和J1。在第36 d至42 d时,L1、J1菌草配方的漆酶活力显著高于C1棉籽壳和C2木屑配方。
2.5 木聚糖酶活力变化
木聚糖酶活力可以显示出银耳菌丝对半纤维素的降解能力。由图5可知,不同配方木聚糖酶活力随栽培时间延长逐渐升高。在菌丝生长18 d,木聚糖酶活力C1 显著高于L1、C2和J1,L1和C2差异不显著,J1最低,与其他差异显著。各配方的木聚糖酶活力均在出菇期达到峰值,从第30 d至42 d(出菇期),C2显著低于J1、C1和L1。
3 讨论与结论
3.1 不同配方工厂化栽培银耳的胞外酶变化趋势
胞外酶活性的高低决定了银耳菌丝在不同时期对栽培基质中营养物质的利用情况。综合各组酶活数据分析,在菌丝生长期,C1的胞外酶活力均高于其他配方,而J1则最低,但是在子实体成熟期,则是J1最高,C2的酶活力最低。从变化趋势分析,C1和C2的变化趋势相同,这两个配方无论是升高和下降的趋势,还是达到峰值的时间,均显示出强的同步性。两个鲜菌草配方与两个对照配方相比,变化的趋势不同。
3.2 不同配方栽培银耳与前人研究结果的对比
在菌丝生长期,C1对木质纤维素和淀粉的利用能力高于其他配方,这可能是C1产量高的原因,赵超等[22]研究表明,胞外酶的活力与子实体产量显著相关。酶活力的变化趋势与王长文[17]的研究结果相同,纤维素和木质素酶活力均是先升高后降低的趋势;各配方半纤维素酶活力呈现出持续升高的趋势,这与王庆福[23]的研究结果类似;淀粉酶的变化趋势则与陈冈等[24]的研究结果不同,他认为银耳的淀粉酶活力是升高降低再升高再降低的趋势,但是在本研究中,棉籽壳配方的淀粉酶活力只有一个峰值,这可能与栽培配方和培养条件不同有关。刘欣怡等[25]研究了鲜菌草和干菌草栽培平菇的胞外酶活性,以木屑配方为对照,发现从菌丝满袋期开始,漆酶的活力先升高后降低,而半纤维素酶则是菌草配方呈逐渐降低趋势,木屑配方呈先升高后降低的趋势,其变化趋势与银耳中的酶活力变化趋势不同,这可能与不同食用菌品种有关。
总体上,不同配方工厂化栽培银耳的羧甲基纤维素酶、滤纸纤维素酶、淀粉酶、漆酶和木聚糖酶等胞外酶活力为棉籽壳(C1)>鲜绿洲1号(L1)>鲜巨菌草(J1)>木屑(C2),说明鲜绿洲1号、鲜巨菌草等鲜菌草工厂化栽培银耳的营养基质利用能力强于木屑,鲜菌草可替代木屑栽培银耳。
参考文献:
[1]黄年来.中国食药用菌学[M].上海:上海科学技术文献出版社,2010.
[2]ZHANG Y, ZHANG Q, LU J, et al.Physicochemical properties of Tremella fuciformis polysaccharide and its interactions with myofibrillar protein[J].Bioactive Carbohydrates and Dietary Fibre,2017, 11:18-25.
[3]LIU X Z, WANG Q M, THEELEN B, et al.Phylogeny of tremellomycetous yeasts and related dimorphic and filamentous basidiomycetes reconstructed from multiple gene sequence analyses[J].Studies in mycology,2015, 81:1-26.
[4]黃年来等.中国银耳生产[M].北京:中国农业出版社, 2000.
[5]上官端琳,龚凤萍,竹玮,等.我国银耳段木栽培技术研究现状[J].食用菌, 2020, 42(2):1-3.
[6]温志强,陈丽芳,李兵兵.农药对银耳生长发育的影响及残留情况分析[J].食用菌学报, 2014, 21(3):70-76.
[7]EL-SHARAKY A S, NEWAIRY A A, ELGUINDY N M, et al.Spermatotoxicity, biochemical changes and histological alteration induced by gossypol in testicular and hepatic tissues of male rats[J].Food and Chemical Toxicology,2010,48(12):3354-3361.
[8]JANERO D R, BURGHARDT B.Protection of rat myocardial phospholipid against peroxidative injury through superoxide-(xanthine oxidase)-dependent, iron-promoted Fenton chemistry by the male contraceptive gossypol[J].Biochem Pharmacol,1988, 37(17): 3335-3342.
[9]RANDEL R D, CHASE C J, WYSE S J.Effects of gossypol and cottonseed products on reproduction of mammals[J].J Anim Sci,1992, 70(5): 1628-1638.
[10]林占熺.菌草学概论[M].北京:中国农业出版社, 2019.
[11]黄玉琴.菌草栽培银耳工艺的研究与应用[D].福州:福建农林大学, 2012.
[12]项丽娟.菌草银耳品质及其主要活性成分的研究[D].福州:福建农林大学, 2012.
[13]王泽辉,林占熺,马运龙,等.鲜菌草工厂化栽培银耳的营养成分分析与评价[J].中国食用菌, 2020, 39(12):73-77.
[14]赖春芬,杨羽茜,张海洋,等.5种食用菌废菌糠的营养成分及胞外酶活性[J].贵州农业科学, 2017, 45(12): 82-85.
[15]陈岗,詹永,杨勇,等.温度对银耳胞外酶活力及营养品质特性的影响[J].食品科学, 2017, 38(23): 113-120.
[16]张职视,林辉,王丽芬,等.斑玉蕈培养过程中酶活性与菌包成熟度的关系[J].中国食用菌, 2016, 35(1): 53-58.
[17]王长文.袋栽银耳栽培关键技术研究[D].福州:福建农林大学,2017.
[18]孙静,耿慧莉,莫德馨.中温α淀粉酶活性的定量测定[J].教学仪器与实验, 2009, 25(11): 44-45.
[19]郭艳艳,阮玲云,冯宏昌,等.不同营养条件下斑玉蕈菌丝生长及产酶特性[J].菌物学报, 2014, 33(3): 697-705.
[20]刘健鹏.灵芝漆酶及部分胞外酶研究[D].北京:中国协和医科大学, 2009.
[21]周晨妍,张金华,马雪婷,等.黑曲霉产木聚糖酶的固态发酵条件优化[J].安徽农业科学, 2010, 38(21): 11052-11054.
[22]赵超,高兆银,刘美珍.不同配方蔗渣培养基对平菇纤维素酶活性和产量的影响[J].浙江农业科学, 2010(1): 31-34.
[23]王慶福.银耳与香灰菌CAZymes差异性研究[D].福州:福建农林大学, 2013.
[24]陈岗,詹永,杨勇,等.温度对银耳胞外酶活力及营养品质特性的影响[J].食品科学, 2017, 38(23): 113-120.
[25]刘欣怡,陈敏,雷雅婷,等.新鲜巨菌草对平菇胞外酶活性以及栽培料营养成分变化的影响[J].江苏农业科学, 2020, 48(2): 159-163.
(责任编辑:柯文辉)
