黑龙江三江国家级自然保护区赤狐的生境选择
2021-01-13李路云刘振生滕丽微
朱 倩 李路云 刘振生,2 滕丽微,2*
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.国家林业局野生动物保护学重点开放实验室,哈尔滨,150040)
生境是动物赖以生存的场所,是保证其能完成生命过程的空间[1-2],了解动物如何选择生境对生态学至关重要[3]。除本身特性以外,野生动物食物的空间分布和生存策略等因素都会影响生境的选择[4-5]。为了满足不同时期的生存需求,动物在不同生活阶段、不同季节对生境的选择机制会发生改变。因此,为了能更好地揭示其生存策略,制定合理的保护措施和资源管理方案,对野生动物各季节生境选择和利用的研究非常必要[5-6]。
食肉动物处于食物网的顶端[7],其生境选择、分布和种群动态等信息对生态保护至关重要[8-9]。国内有关食肉动物生境选择的研究主要集中在鼬科(Mustelidae)、熊科(Ursidae)和猫科(Felidae)等动物。对犬科(Canidae)狐属(Vulpes)动物的研究尚不完善,多集中在活动节律和洞穴选择等方面,对其不同季节生境选择机制研究较少。周文扬等[10-12]发现狐属动物常利用啮齿动物的洞穴,繁殖期赤狐洞穴选择的主要因子是水源、隐蔽性和安全性,其次是坡向、坡位和坡度[13];从3种犬科动物春季洞穴选择的研究得出,柳灌丛、高草坡地分别是赤狐(Vulpesvulpes)和沙狐(Vulpescorsac)春季偏爱的生境类型[14],且通过对春季沙狐的洞穴选择的研究得出,不同生境中影响沙狐洞穴选择的主要生态因子存在差异[14-15]。
赤狐,分布广泛,繁殖率高,是适宜性极强的食肉动物[16],典型的机会主义者捕食者[17],在调节鼠类数量和控制鼠害方面起着重要的作用[18]。国内外学者对赤狐家域和食性的研究已有深入了解,而对生境特征和选择机制的研究甚少[19]。本文在前人研究的基础之上,对黑龙江三江国家级自然保护区赤狐4个季节的生境选择进行比较,揭示其不同季节生境选择偏好和影响因子的差异,以期更好地了解赤狐各时期的生物学需求,为保护区生态环境及赤狐的保护和管理提供参考依据。
1 研究地区自然概况
黑龙江三江国家级自然保护区位于三江平原的东北角,地跨抚远、同江两县市,133°43′—134°47′E,47°26′—48°23′N。保护区总面积198 089 hm2,属于温带湿润半湿润大陆性季风气候,年平均气温较低,冬季长而严寒,夏季短而炎热,水量充沛,阳光充足。保护区集山地、平原、沼泽为一体,属比较完整的森林、湿地、海洋生态系统。区内野生动植物资源丰富,分布有植物992种,记录到兽类38种,鸟类259种,两栖爬行类12种,鱼类77种。其中国家Ⅰ级和Ⅱ级保护动物共53种,黑龙江省地方重点保护动物53种。
2 研究方法
2.1 野外数据采集
于2013—2014年,分冬(2013年12月—2014年1月)、春(2014年5—6月)、夏(2014年7—8月)、秋(2014年10—11月)4个季节,采用痕迹检验法和样方法进行野外数据采集。根据当地护林员的经验,在赤狐出没或发现赤狐的粪便、足迹、洞穴等痕迹的区域,设置1个10 m×10 m的样方,用全球定位仪(GPS)定位。同时,在未发现赤狐痕迹的区域设置对照样方,结合研究地区地形地貌及相关研究工作,测定样方内的10个生态因子。测定方法参照张洪海等[20-21]的方法。
(1)植被类型:按样方内植物生长类型和外貌分为岛状林、草甸、湿地和农田;同时根据植被类型将生境类型划分为岛状林、草甸、湿地和农田。
(2)郁闭度:林冠覆盖面积与地表面积的比例,采用抬头垂直昂视的方法,判断天空被树冠覆盖的面积比例。
(3)灌木密度:统计样方内每100 m2灌木的丛数。
(4)灌木高度:测算样方内灌木的平均高度。
(5)灌木距离:测算样方中心点距最近的灌木的距离。
(6)植被盖度:测算样方内草本的覆盖面积占整个样方面积的百分比。
(7)食物丰富度:样方内可被赤狐利用的食物资源,如小型啮齿类、鸟类等动物或足迹,其中高>8只,中6—8只,低<6只。
(8)水源距离:估算样方中心距河流、湖泊、沼泽等的距离。
(9)人为干扰距离:估算样方中心距居民点等人为干扰源的距离。
(10)隐蔽级:在每个样方的中央放一赤狐身高平均标杆,在样方对角线上距离标杆20 m从东南西北4个方位观测,记录标尺的可见程度,求其平均值。
2.2 数据处理和分析
(1)运用降维的思想,进行主成分分析,取特征值大于1的成分作为主成分,以了解赤狐利用生境的特征值,确定生境选择中的主要因子。
(2)利用拟合优度卡方检验检测赤狐在各个季节对10种生态因子是否有选择性,再利用Bonferroni不等式分析赤狐对生态因子的偏好或回避。
(3)利用Kendall相关系数对赤狐偏好和回避的生境分别与各生态因子进行相关性分析,得到各类生境中生态因子的特征值。Kendall相关系数用非参数检验方法来度量定序变量间的线性相关关系。
(4)对赤狐生境选择的生态因子进行卡方差异性检验。当χ2值越大时说明两个季节利用生境中的生态因子差异性越显著,P<0.01表示极显著;P<0.05表示显著[22]。
3 结果
研究区域共设置实验样方 475个(春季105个、夏季116个、秋季114个、冬季140个),对照样方229个(春季88个、夏季41个、秋季57个、冬季43个),逐一测定样方内的10种生境因子。
3.1 赤狐冬季的生境选择
3.1.1 赤狐冬季生态因子的主成分分析
主成分分析结果表明,赤狐冬季利用生境的前4个因子特征值大于1,累计贡献率为59.376%。第1主成分贡献率达到21.531%,载荷系数绝对值较大的为植被类型(0.760)和灌木高度(-0.767),第2主成分中载荷系数绝对值较大的为郁闭度(0.623),第3主成分中载荷系数绝对值较大的为食物丰富度(0.665),第4主成分中载荷系数绝对值较大的为隐蔽级(0.596),说明植被类型、灌木高度、郁闭度食物丰度以及隐蔽级是赤狐冬季生境选择的主要因子(表1)。
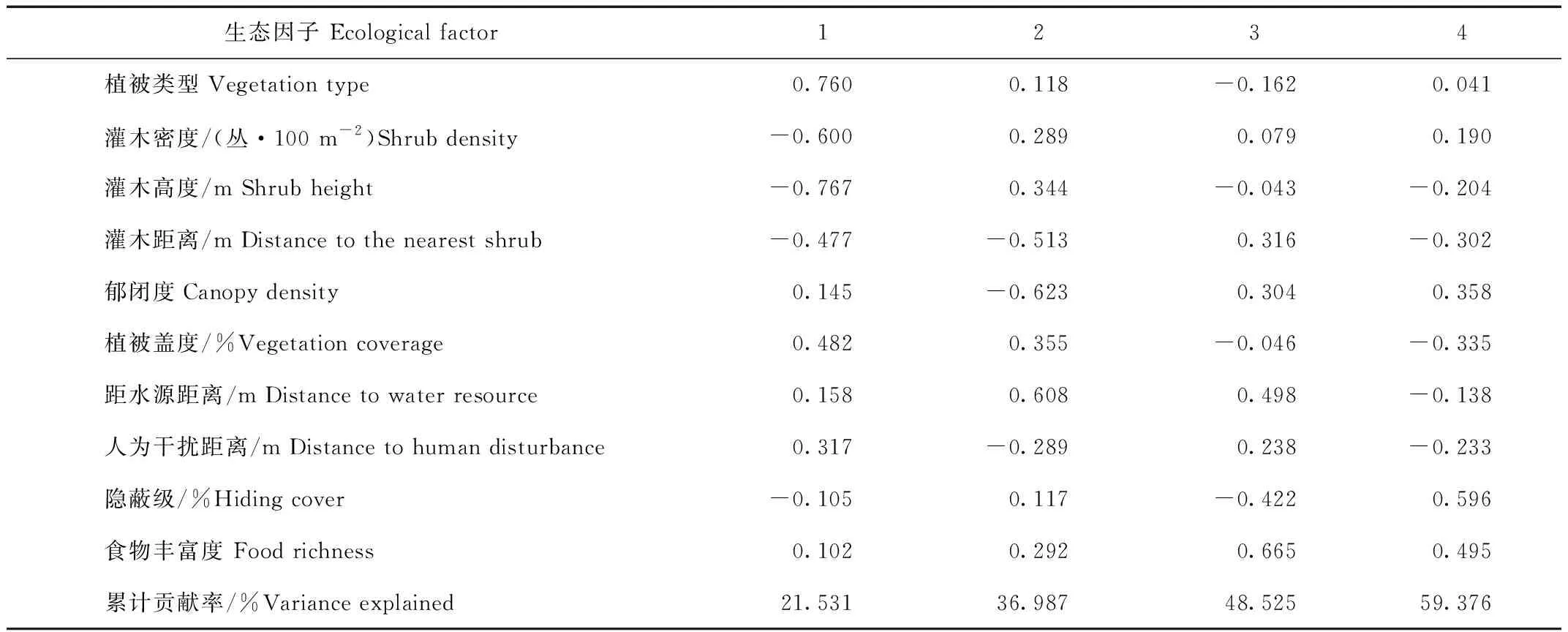
表1 赤狐冬季的10种生态因子的主成分分析Tab.1 Principal components of 10 ecological factors of red fox in winter
3.1.2 赤狐冬季对生境的利用和选择
利用Boferroni不等式对赤狐各种生境进行“可利用-利用”判别分析得出,赤狐在冬季偏好选择农田和岛状林生境,避免选择草甸和湿地,说明农田和岛状林为赤狐冬季的最适宜生境;同时,赤狐对这两种偏好生境内的各生态因子也具有选择性,更偏好灌木密度>10丛、灌木高度1—2 m、灌木距离>3 m、郁闭度<50%、植被盖度<50%、距水源距离<500 m、人为干扰距离<500 m和食物丰富度高的农田和岛状林,避免选择隐蔽级、郁闭度以及植被盖度过高的农田和岛状林生境(表2)。

表2 赤狐冬季对10种生态因子的选择性Tab.2 Utilization and selection of 10 factors in habitat used by red fox during winter

续表2
3.1.3 赤狐冬季生境选择与各生态因子的相关性分析
冬季,赤狐对植被类型的选择与植被盖度(R=0.377,P=0.000)和隐蔽级(R=0.231,P=0.001)呈极显著的正相关,与灌木密度(R=-0.340,P=0.000)、灌木高度(R=-0.489,P=0.000)和灌木距离(R=-0.442,P=0.000)存在极显著的负相关,说明这几个因子在赤狐生境选择中起到非常重要作用;人为干扰距离(R=0.038,P=0.593)、距水源距离(R=0.014,P=0.841)和食物丰富度(R=0.074,P=0.295)与生境类型选择的相关性不显著,说明这些因子在生境选择中作用较弱。
3.2 赤狐春季的生境选择
3.2.1 赤狐春季生态因子的主成分分析
赤狐春季生境选择的前4个因子特征值大于1,累计贡献率为63.004%。第1主成分贡献率达到20.452%,载荷系数绝对值较大的为食物丰富度(0.744),第2主成分中载荷系数绝对值较大的是灌木密度(0.772),第3主成分中载荷系数绝对值较大的是植被类型(0.792),第4主成分中载荷系数绝对值较大的是灌木距离(0.731),说明食物丰度、灌木密度、植被类型和灌木距离为赤狐春季生境选择的主要因子(表3)。

表3 赤狐春季的10种生态因子的主成分分析Tab.3 Principal components of 10 ecological factors of red fox in spring
3.2.2 赤狐春季对生境的利用和选择
赤狐在春季偏好选择农田和湿地、随机选择岛状林,避免选择草甸生境,说明在春季赤狐的最适宜生境类型为农田和湿地。同时,赤狐对这两种生境内的生态因子也具有选择性,偏好灌木密度<10丛、灌木高度<1 m、郁闭度<50%、植被盖度<50%、水源距离<300 m、人为干扰距离>500 m的农田和湿地,避免食物丰度低的生境(表4)。
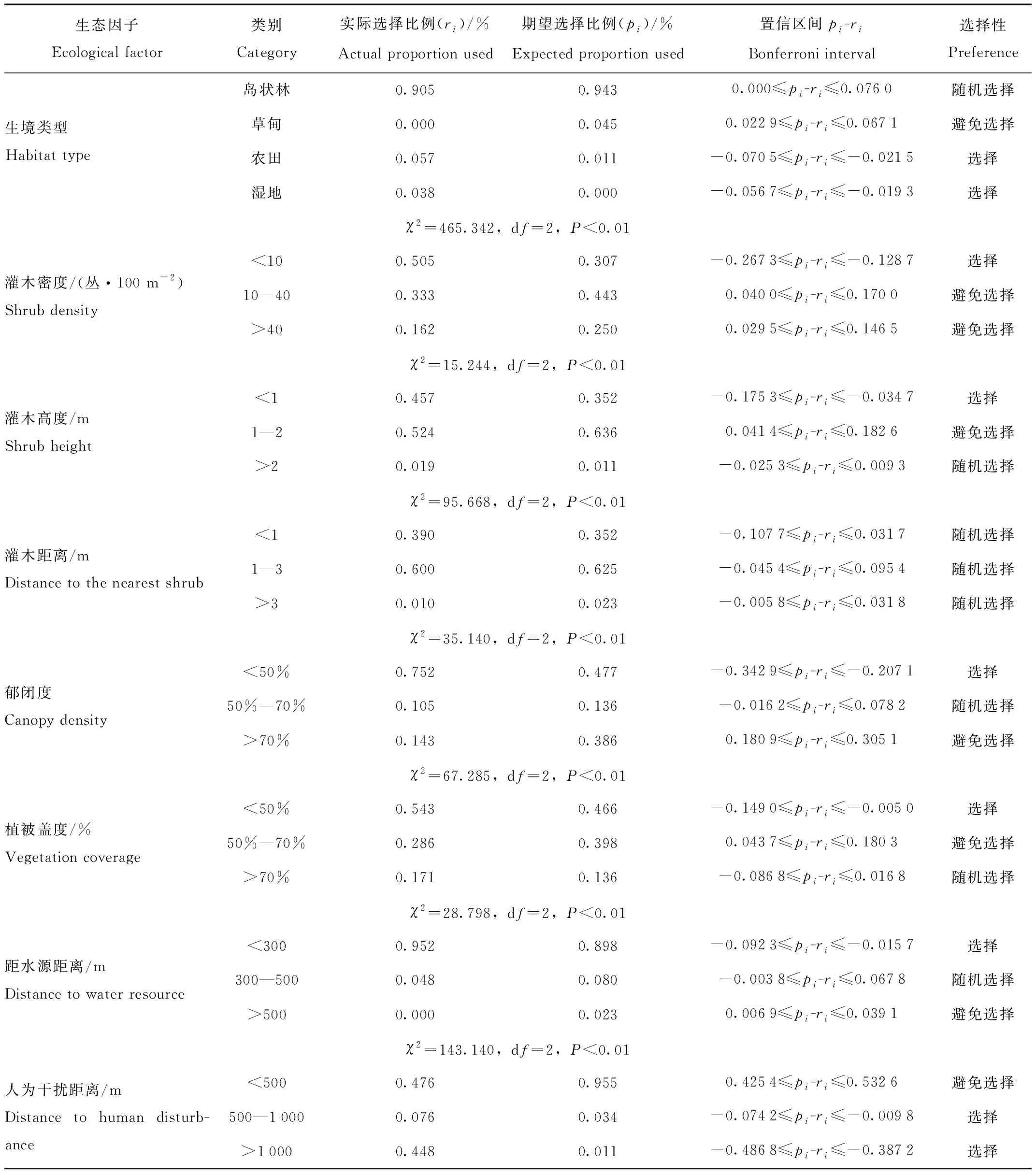
表4 赤狐春季对10种生态因子的选择性Tab.4 Utilization and selection of 10 factors in habitat used by red fox during spring

续表4
3.2.3 赤狐春季生境选择与各生态因子的相关性分析
赤狐春季生境类型的选择与灌木高度(R=-0.225,P=0.001)和灌木距离(R=-0.204,P=0.003)存在极显著负相关,说明灌木高度和灌木距离在赤狐春季生境选择中具有非常重要的作用;生境类型和隐蔽级(R=-0.161,P=0.024)成显著负相关;郁闭度(R=0.053,P=0.430)、植被盖度(R=0.023,P=0.738)、人为干扰距离(R=0.129,P=0.067)、食物丰富度(R=-0.050,P=0.121)和距水源距离(R=-0.026,P=0.710)相关性较差,说明这些因子在生境选择中作用较弱。
3.3 赤狐夏季的生境选择
3.3.1 赤狐夏季生态因子的主成分分析
夏季赤狐生境选择的前4个因子特征值大于1,累计贡献率为64.745%。第1主成分贡献率达到24.862%,载荷系数绝对值较大的为距水源距离(0.706),第2主成分中载荷系数绝对值较大的是植被盖度(0.746)和隐蔽级(0.734),第3主成分中载荷系数绝对值较大的是距水源距离(0.627),第4主成分中载荷系数绝对值较大的是人为干扰距离(0.565),说明植被盖度、隐蔽级、距水源距离及人为干扰距离是赤狐夏季生境选择的主要因子(表5)。
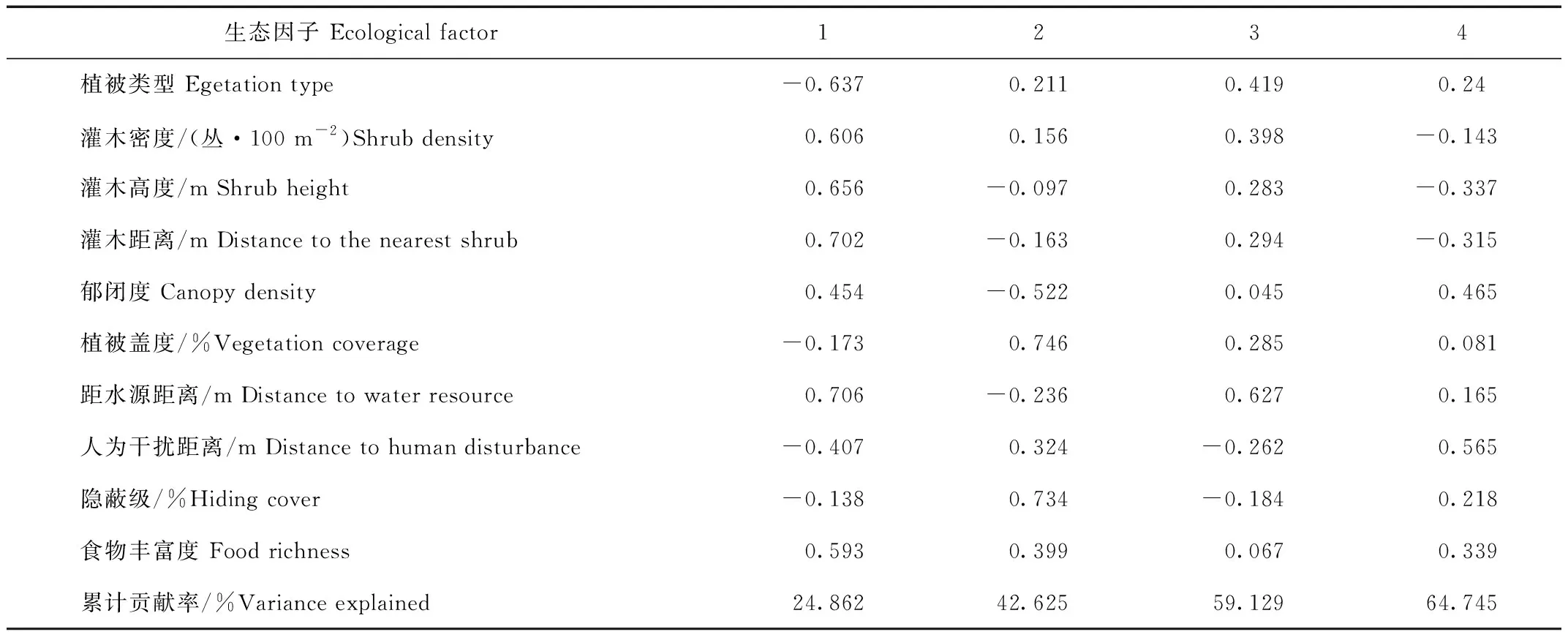
表5 赤狐夏季的10种生态因子的主成分分析Tab.5 Principal components of 10 ecological factors of red fox in summer
3.3.2 赤狐夏季对生境的利用和选择
赤狐夏季偏好选择岛状林、农田,随机选择草甸,避免选择湿地生境,说明岛状林和农田为赤狐夏季最适宜生境。同时,赤狐对偏好生境内的各生态因子也具有选择性,更喜欢选择灌木密度10—40丛、灌木高度>2 m、郁闭度>70%、植被盖度50%—70%、水源距离>500 m、人为干扰距离>500 m、隐蔽级<50%和食物丰富度高的岛状林和农田,对生境内灌木距离没有严格的选择,避免选择距水源距离近<500 m、人为干扰距离近和食物丰富度低的生境。(表6)。
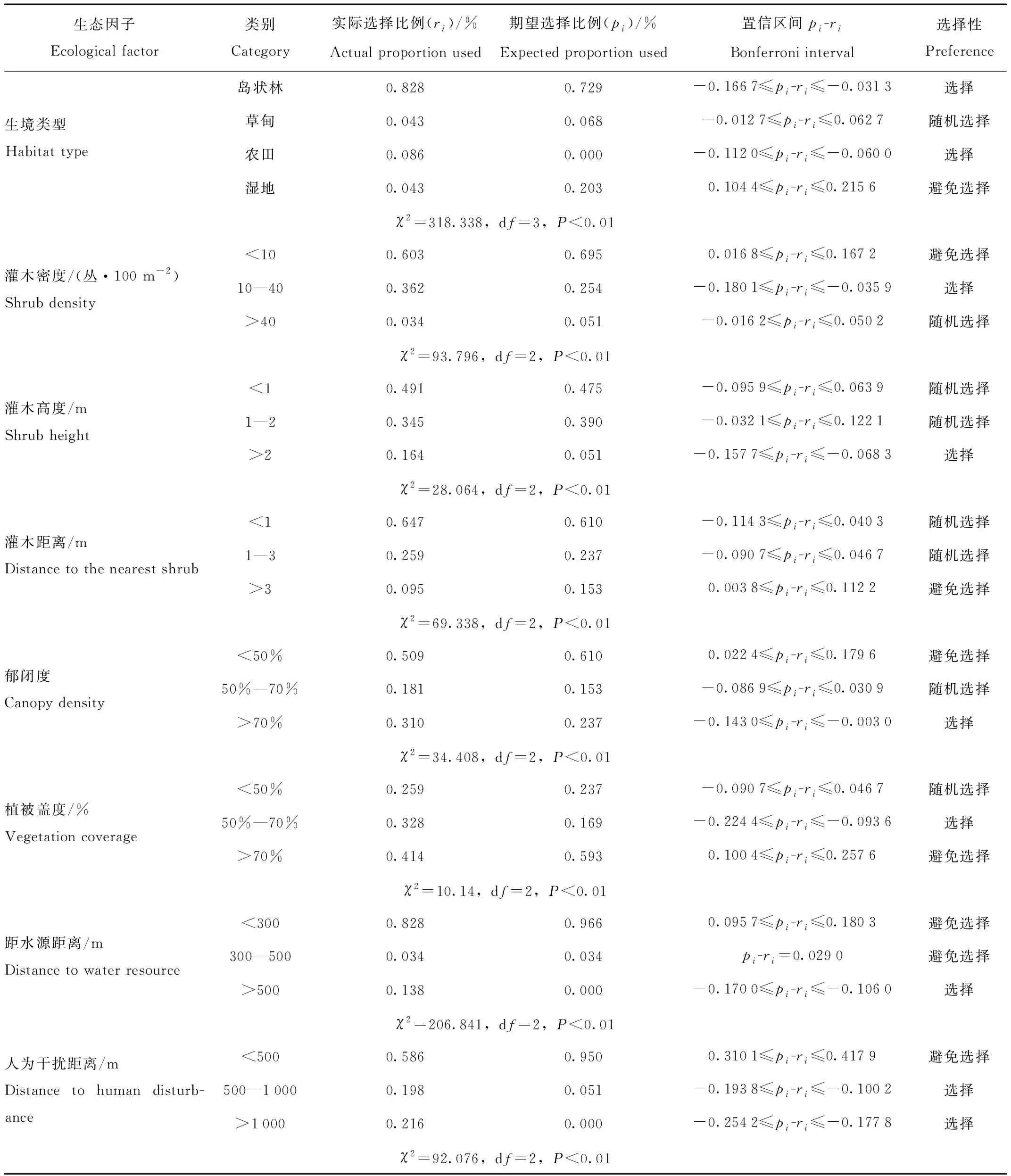
表6 赤狐夏季对10种生态因子的选择性Tab.6 Utilization and selection of 10 factors in habitat used by red fox during summer
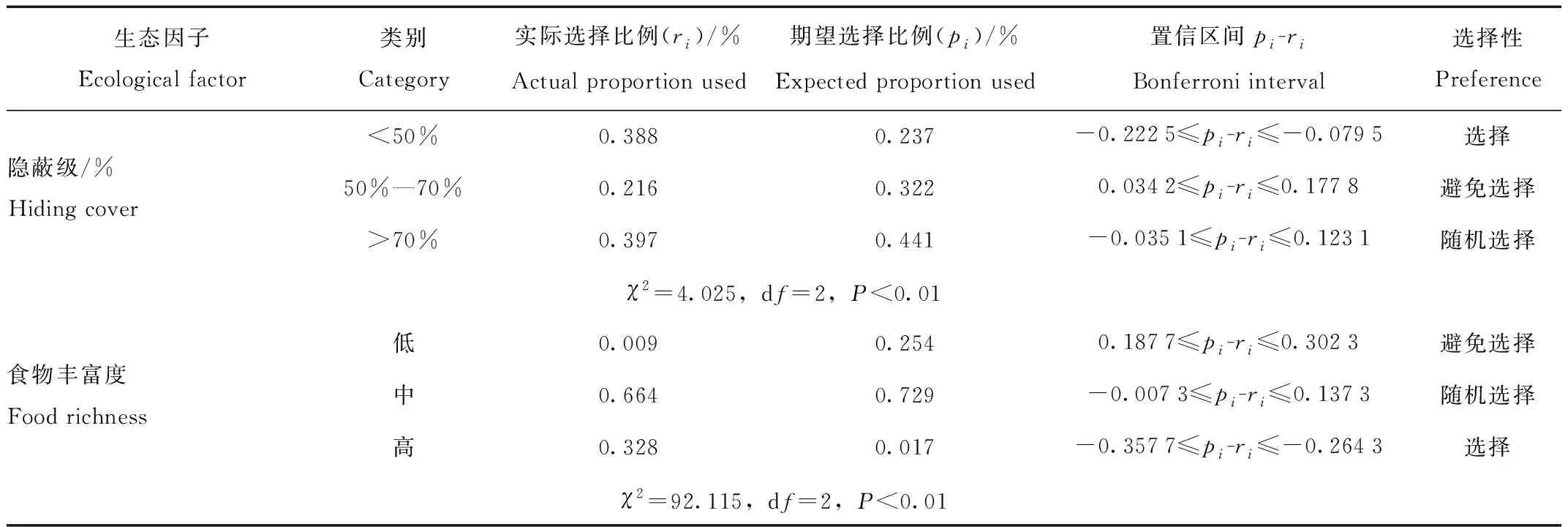
续表6
3.3.3 赤狐夏季生境选择与各生态因子的相关性分析
赤狐夏季生境类型的选择与水源距离(R=0.504,P=0.000)存在极显著的正相关,与灌木密度(R=-0.232,P=0.003)、人为干扰距离(R=-0.210,P=0.006)和食物丰富度(R=-0.247,P=0.001)存在极显著的负相关,说明这些因子在赤狐夏季生境选择中起到非常重要的作用;生境类型与灌木高度(R=-0.174,P=0.020)、灌木距离(R=-0.177,P=0.020)和郁闭度(R=-0.184,P=0.014)存在显著的负相关;植被盖度(R=0.145,P=0.051)和隐蔽级(R=-0.008,P=0.919)相关性较差,说明这两个生态因子在赤狐生境选择中作用较弱。
3.4 赤狐秋季的生境选择
3.4.1 赤狐秋季生态因子的主成分分析
秋季赤狐生境选择的前4个因子特征值大于1,累计贡献率为68.622%。第1主成分贡献率达到26.682%,载荷系数绝对值较大的为植被类型(-0.737)和灌木距离(0.737),第2主成分中载荷系数绝对值较大的是植被盖度(0.783)和隐蔽级(0.732),第3主成分中载荷系数绝对值较大的是距水源距离(0.727),第4主成分中载荷系数绝对值较大的是郁闭度(0.565),说明植被类型、灌木距离、植被盖度、距水源距离及郁闭度是赤狐秋季生境选择的主要因子(表7)。
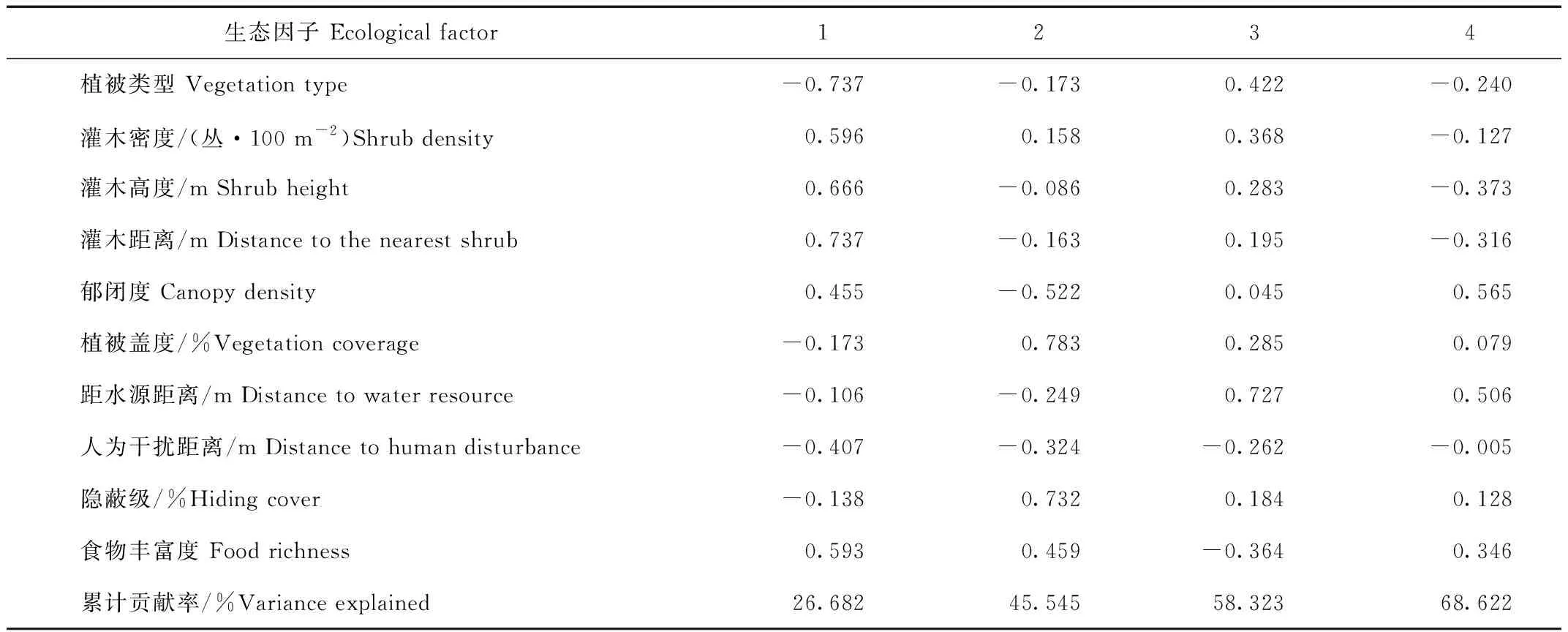
表7 赤狐秋季的10种生态因子的主成分分析Tab.7 Principal components of 10 ecological factors of red fox in autumn
3.4.2 赤狐秋季对生境的利用和选择
在秋季,赤狐偏好选择农田和湿地、随机选择草甸,避免选择岛状林生境,说明秋季赤狐的最适宜生境为农田和湿地。同时,赤狐对偏好生境内的各生态因子也具有选择性,更喜欢选择灌木密度>10丛、灌木高度<2 m、郁闭度<50%、植被盖度<70%、距水源距离<500 m、人为干扰距离<500 m、隐蔽级<70%且食物丰富度高的农田和湿地生境,对植被距离选择性不明显,避免选择灌木密度<10丛、灌木高度>2 m、郁闭度>50%、植被盖度>70%、距水源距离>500、隐蔽级<70%和食物丰富度一般的生境(表8)。

续表8
3.4.3 赤狐秋季生境选择与各生态因子的相关性分析
秋季赤狐生境选择的类型与人为干扰距离(R=0.218,P=0.002)呈极显著的正相关,说明人为干扰在赤狐生境选择中有非常重要作用;与灌木密度(R=-0.286,P=0.000)、灌木高度(R=-0.346,P=0.000)、灌木距离(R=-0.367,P=0.000)、郁闭度(R=-0.275,P=0.000)和食物丰富度(R=-0.591,P=0.000)呈极显著负相关,即这些因子在赤狐生境选择中同样起到非常重要的作用;与植被盖度(R=0.125,P=0.077)、距水源距离(R=0.042,P=0.563)和隐蔽级(R=0.065,P=0.361)相关性较差,说明这些因子在生境选择中作用较弱。
3.5 赤狐全年生境选择的比较
利用卡方检验对赤狐利用生境生态因子进行比较,赤狐在植被类型(χ2=839.771,df=3,P<0.01)和食物丰富度(χ2=237.996,df=2,P<0.01)选择上存在极显著差异,利用Kruskal-WallisH检验对赤狐其余的生态因子进行比较,结果表明:赤狐在冬夏季偏好选择岛状林和农田生境,春秋季偏好选择湿地和农田;不同季节生境的选择在灌木密度、灌木高度、灌木距离、植被盖度、距水源距离、人为干扰距离和隐蔽级上差异极显著(表9)。

表9 赤狐全年利用生境7种生态因子比较Tab.9 K Independent samples Kruskal-Wallis H test was used to test the differences on used habitat among four seasons
4 讨论
已有研究表明,赤狐喜欢生活在食物丰富、距水源较近、隐蔽性较好且远离居民点或人为干扰较弱的灌丛、草地或林地中[23],食物、水源和隐蔽条件是影响其生境选择的重要因素[24]。本研究发现,赤狐在黑龙江三江保护区偏好选择食物丰富度高的农田生境,但不同季节在生境选择时对各生态因子的重视程度出现了差异。各季节赤狐在生境选择时对灌木密度、灌木距离、灌木高度、植被盖度、水源距离、人为干扰距离和隐蔽级的选择差异显著。我们猜测不同季节生境选择的差异是赤狐对不同季节生理需求的响应,是为了能够在不同时段获得更大的收益[25]。
赤狐在冬季偏好选择农田和岛状林,郁闭度、灌木高度、食物丰富度和隐蔽级是其生境选择的主要因子。冬季的三江保护区风雪较大,寒冷的天气使得赤狐耗能增加[26]。因此,赤狐偏好选择具有较好隐蔽性和保温性,植被较稠密的岛状林,或岛状林与农田交错区,这些区域不仅雪兔(Lepustimidus)和鸟类的数量相对较多,还可减少由于长距离迁徙寻找食物而带来过多的能量消耗。且由于冬季积雪深厚,人为活动弱,人为干扰距离和水源距离这两生态因子在赤狐生境选择时的作用较弱。
张洪海等[14]对赤狐春季洞穴的选择研究表明,影响赤狐春季洞穴选择的前3位主要因子是地形、水源和人为干扰,与本研究结果基本相似。赤狐在春季偏好选择农田和湿地,此类生境能满足赤狐对水源的需求。春季的天气开始回暖,植被开始萌芽,各类动物活动增加,农耕活动陆续地开展,赤狐也开始进入繁殖期。赤狐偏好选择距居民点远且隐蔽性较高的生境,从而避免受到外界的干扰[12]。灌木的分布情况实际上决定了生境的隐蔽性和抗干扰程度,是春季赤狐生境选择的主要因子。
三江平原地区夏季炎热、蚊虫较多。赤狐夏季偏好选择岛状林、农田。距水源距离、植被盖度及距居民点距离是其生境选择的主要因子,在生境选择时起到重要作用。夏季是赤狐生产哺育时期,产仔洞穴多选择在稀疏灌丛或开阔的岛状林,这类生境既具备较好的隐蔽性,减少被捕食的风险,同时能兼顾幼兽对阳光的需求,这与周文扬等[12]和Nakazono等[27]的研究结果一致。
秋季,赤狐偏好选择农田和湿地。距居民点距离、灌木距离、距水源距离、植被盖度和郁闭度是其生境选择的主要因子,可以总的归纳为隐蔽级和食物丰度在其生境选择中起到重要作用。我们研究时观察到,赤狐秋季喜欢在鼠洞较多,野兔和鸟类经常出没的灌丛中筑巢。秋季树叶开始枯萎,环境内视野逐渐开阔,隐蔽级下降,且保护区内人类活动频繁,此时赤狐会更为注重生境的隐蔽性。但有研究表明动物在选择生境时会对干扰距离和食物做出权衡[28-29]。所以,赤狐秋季生境选择会重点考虑隐蔽级,灌木的生长情况和人为干扰距离这3个生态因子。
总的来说,赤狐在进行生境选择时会受诸多因素的影响,且存在季节性差异,因此需根据季节的变化对其进行针对性保护。
