GP73参与肝癌索拉菲尼获得性耐药的初步研究
2020-11-16杨欢彭雨蒙李辉龙韦猛陈智炜唐置鸿孟维达韦滔魏从文钟辉吴飞翔
杨欢,彭雨蒙,李辉龙,韦猛,陈智炜,唐置鸿,孟维达,韦滔,魏从文,钟辉,吴飞翔,3,4
1.广西医科大学附属肿瘤医院 肝胆外科,广西 南宁 5300021;2.军事医学研究院 生物工程研究所,北京 100850;3.广西肝癌诊疗工程技术研究中心,广西 南宁 530021;4.区域性高发肿瘤早期防治研究教育部重点实验室,广西 南宁 530021
原发性肝细胞癌(hepatocellular carcinoma,HCC)是常见的恶性肿瘤之一,每年新发病例和死亡病例均超过80万例[1-2]。目前,中晚期肝癌患者以局部治疗、系统治疗等为主[3]。目前各种常见治疗手段都有局限性,所以现在肝癌治疗领域的共识是多学科协作和多种治疗方法联用的综合治疗[4]。综合治疗方法包括介入治疗、消融治疗、靶向治疗、免疫治疗和放化疗等,由于分子靶向治疗的选择性、灵敏性和效率都很高,靶向治疗以成为传统肝癌治疗手段之外极其重要的治疗方法[5]。作为第一个被美国FDA批准用于治疗肝癌的分子靶向治疗药物,索拉菲尼是肝癌系统性治疗中惟一能够改善患者存活率、显著延长晚期患者总体生存时间的化疗药物。
索拉菲尼是一种新型多靶点抗肿瘤药物,通过与细胞内ATP结合位点上的三磷酸腺苷竞争性结合,阻断酪氨酸激酶和丝/苏氨酸激酶的活性。大量临床研究表明,绝大部分HCC患者通过一段时间的索拉菲尼药物治疗后,表现出明显的药物耐受特征,导致索拉菲尼对HCC的疗效欠佳。多种机制与索拉菲尼的获得性耐药相关,有报道显示肿瘤细胞SIRT1、EGFR或其配体等分子的过表达,PI3K/Akt、JAK/STAT3信号通路的异常激活,以及Ras/Raf/MEK/ERK通路的活性下调均降低HCC对索拉菲尼的敏感性[6-7]。
高尔基蛋白73(Golgi protein 73,GP73)又名GOLM1(Golgimembraneprotein1)或 GOLPH2(Golgi phosphorprotein 2),是2000年发现的一种高尔基膜蛋白,在正常肝脏的胆管上皮细胞中稳定表达[8]。70%以上的肝癌病人,GP73蛋白在血清及肝组织中的表达水平均显著上调。最近的研究已证实GP73在肝癌发生发展和转移中的重要作用,张宏冰等研究发现mTORC1能够通过调节GP73促进肝癌的发生发展[9]。我们前期研究发现高表达的GP73抑制了由饥饿诱导的细胞自噬的激活,而GP73敲除细胞的自噬水平明显升高。自噬在肝癌索拉菲尼的原发性耐药中具有双向作用。mTOR抑制剂雷帕霉素(rapamycin)并不能完全恢复耐药细胞株对索拉菲尼的敏感性。这就提示我们,在索拉菲尼获得性耐药的HCC细胞中还有别的机制参与自噬的负调控。因此,研究GP73与索拉菲尼获得性耐药的关系对原发性肝癌的分子靶向治疗具有重要意义。
在本研究中,我们探讨了GP73在肝癌索拉菲尼耐药细胞株中的表达情况,以及GP73对索拉菲尼耐药性的影响,为GP73参与肝癌索拉菲尼获得性耐药的分子机制研究提供实验基础。
1 材料与方法
1.1 材料
肝癌细胞HepG2(本实验室保存);DMEM高糖培养基、胎牛血清(Gibco公司);索拉菲尼(Sigma-Aldrih公司);转染试剂VigoFect(Qbiogene公司);MTT检测试剂盒(碧云天生物技术公司);Flag-GP73、Flag-Vector(本实验室构建);si-GP73、si-scramble(吉玛基因公司);anti-GP73抗体、anti-Tubulin抗体(Abcam公司)。
1.2 细胞培养与转染
HepG2细胞用含10%胎牛血清的DMEM高糖培养基常规培养,构建耐药细胞株时,在培养基中加入不同浓度的索拉菲尼。细胞转染前1 h更换一半新鲜的DMEM培养基,按说明书要求配制小干扰RNA(siRNA)和转染试剂混合液,滴加入细胞,常规培养,4~6 h后补充另一半培养基。
1.3 MTT法检测细胞存活率
细胞接种于96孔细胞培养板,每孔加入10 μL MTT溶液(5 mg/mL),继续培养4 h,酶联免疫检测仪测定各孔的D490nm值,计算细胞存活率。
1.4 蛋白免疫印迹实验
收集细胞,1×PBS冲洗3次,3000 r/min离心3 min后弃上清。加入适量1×PBS重悬细胞,加入等量4×SDS缓冲液[10%甘油,2%十二烷基苯磺酸钠,50 mmol/L Tris-HCl(pH6.8),2.5% β巯基乙醇,0.1%溴酚蓝],沸水浴15 min,12 000 r/min低温离心10 min,取上清进行SDS-PAGE,半干转到PVDF膜上,用5%脱脂牛奶液室温封闭1 h,一抗和二抗分别室温孵育1 h,TBST洗3次,每次5 min,将Perkin Elmer的ECL发光液均匀涂在PVDF膜上,暗室显影。
1.5 数据分析处理
采用Graphpad prism 8.0进行统计学分析;计量资料采用x±s表示,组间比较采用双因素方差分析;P<0.05表示差异有统计学意义。
2 结果
2.1 HepG2细胞索拉菲尼耐药株的构建
在HepG2细胞培养基中加入索拉菲尼,使其终浓度为0.5 μmol/L,连续作用1周;换成新鲜培养基,镜下观察细胞正常生长后,增加0.25 μmol/L溶液继续培养1周。如此反复换液、传代,逐步提高索拉菲尼浓度(0.5~5 μmol/L)间歇诱导,最终获得HepG2细胞的索拉菲尼耐药株(HepG2-SR)。为了验证构建的索拉菲尼耐药细胞株对索拉菲尼的敏感性降低,在野生型HepG2细胞及索拉菲尼获得性耐药细胞株中加入不同浓度的索拉菲尼,MTT法检测细胞的存活率。结果显示,培养基中索拉菲尼浓度越高,细胞存活率越低;索拉菲尼耐药细胞株对索拉菲尼的敏感性比野生型细胞大幅降低(P<0.05)(图1)。HepG2索拉菲尼耐药细胞株建立成功。
2.2 GP73在肝癌索拉菲尼耐药细胞株中的表达明显升高
收集野生型HepG2细胞和HepG2索拉菲尼耐药株细胞,分别加入等量的PBS和4×SDS缓冲液重悬,沸水浴15 min使细胞充分裂解。12 000 r/min低温离心10 min,取等量上清进行SDSPAGE。半干转到PVDF膜上,用5%脱脂牛奶液室温封闭 1 h,anti-GP73抗体(1∶1000 TBST溶解)、anti-Tubulin抗体(1∶5000 TBST溶解)室温摇床孵育1 h,用5%脱脂奶粉以1∶2000的比例稀释羊抗鼠HRP-conjugated二抗,室温摇床孵育1 h,TBST洗3次,以1∶1的比例配制ECL发光液显影。结果显示,GP73在肝癌索拉菲尼耐药细胞株中的表达比野生型细胞株高(图2)。
2.3 过表达GP73的HepG2细胞对索拉菲尼敏感性降低
GP73在肝癌索拉菲尼耐药细胞株中高表达,接下来将GP73在野生型HepG2细胞中高表达,研究GP73在肝癌细胞中对索拉菲尼敏感性的影响。在野生型HepG2细胞中转染Flag-GP73,对照组转染等量Flag-Vector,24 h后加入不同浓度的索拉菲尼(0、5、10、15、20 μmol/L)处理细胞,48 h后收取细胞,MTT法检测细胞存活。结果表明,随着索拉菲尼的浓度升高,细胞存活率明显越低;而过表达GP73的HepG2细胞(Flag-GP73)对索拉菲尼的敏感性较对照组(Flag-Vector)降低(P<0.05)(图 3)。
2.4 耐药细胞株敲低GP73能够升高索拉菲尼的敏感性
GP73过表达后细胞对索拉菲尼的敏感性降低,接下来探讨在肝癌索拉菲尼耐药细胞株中敲低GP73后对索拉菲尼敏感性的影响。在已构建的索拉菲尼耐药细胞株中转染si-GP73敲低细胞中的GP73水平,对照组转染等量的si-scramble。24 h后加入不同浓度的索拉菲尼(0、5、10、15、20 μmol/L)处理细胞,48 h后收取细胞,MTT法检测细胞存活。结果表明,随着索拉菲尼的浓度升高,细胞存活率明显越低;而在索拉菲尼耐药细胞中敲低GP73后,耐药细胞株(si-GP73)对索拉菲尼的敏感性高于对照组(si-scramble)(P<0.05)(图4),表明耐药细胞株敲低GP73能够升高索拉菲尼的敏感性。
3 讨论
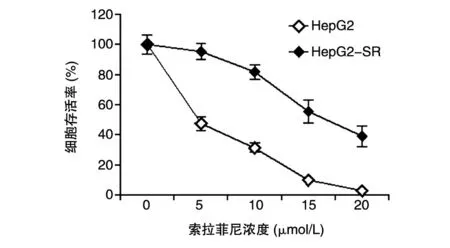
图1 索拉菲尼耐药细胞株对索拉非尼的敏感性降低
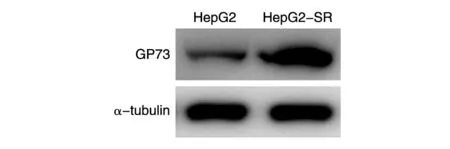
图2 GP73在肝癌索拉菲尼耐药细胞株中高表达
原发性肝细胞癌作为发病率和病死率极高的癌症之一,一直都是科研人员及临床工作者努力攻克的难题。肝癌具有生长速度快、恶性程度高、易复发转移及药物耐受等特点,因此肝癌患者的预后较差[10]。索拉菲尼作为多靶点的分子药物,在肝癌治疗中能够改善患者存活率,但其疗效仍然有限,耐药性的产生可能是导致其化疗失败的主要原因[11]。因此,研究肝细胞癌的耐药机制,对于提高其化疗疗效,指导临床用药有着重要的价值。
GP73不仅可以作为肝癌的肿瘤标志物,也参与肝癌的发生发展过程[12]。我们研究发现,在索拉菲尼获得性耐药的肝癌细胞株中,位于高尔基体的糖蛋白GP73的表达水平显著升高。此外,GP73过表达细胞对索拉菲尼的敏感性降低,GP73敲低恢复了耐药细胞对索拉菲尼的敏感性。因此,GP73很可能参与到原发性肝癌索拉菲尼耐药的机制当中。
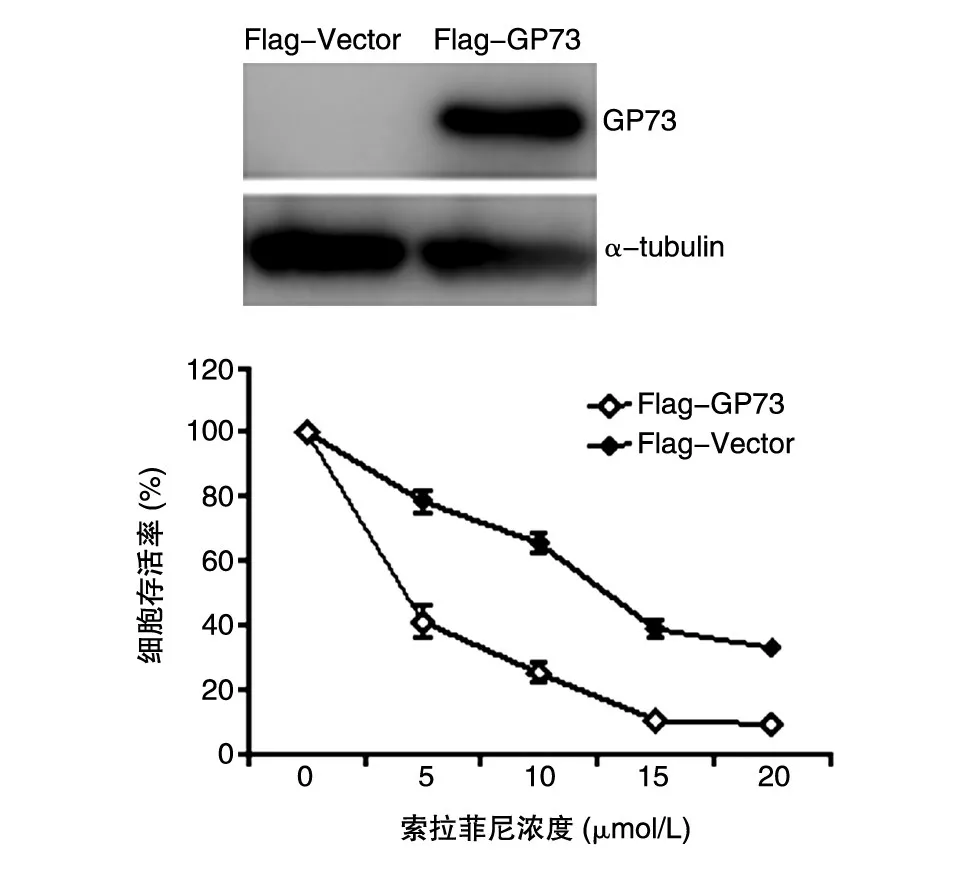
图3 GP73过表达细胞对索拉菲尼的敏感性降低
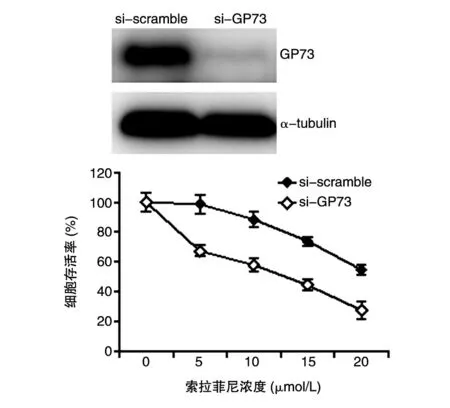
图4 敲低GP73能升高索拉菲尼耐药细胞株对索拉菲尼的敏感性
此前有研究发现自噬与肿瘤的发生密切相关[13]。Shimizu等发现,索拉菲尼可以激活HCC细胞的自噬,诱导自噬小体的形成,增强自噬的活性;而将自噬抑制剂与索拉菲尼联用后,细胞凋亡增多,生存力下降,肿瘤生长受到明显抑制。然而,Bareford等用索拉菲尼与自噬诱导剂培美曲塞联合加入HCC细胞,发现培美曲塞与索拉菲尼可起协同作用[14]。可见,自噬在肝癌索拉菲尼的原发性耐药中具有双向作用。早前我们实验室发现高表达的GP73抑制了由饥饿诱导的细胞自噬的激活,而GP73敲除细胞的自噬水平明显升高。因而,GP73很可能通过负调控自噬,进而影响肝癌索拉菲尼获得性耐药。
综上所述,我们发现GP73在肝癌索拉菲尼获得性耐药的过程中有重要作用,为深入研究索拉菲尼耐药的分子机制奠定了实验基础。
