西部乳源乳酸菌胆盐解离能力筛选及其性能评价
2020-11-12郗梦露张兴吉葛武鹏
郗梦露 张兴吉 葛武鹏
(西北农林科技大学食品科与工程学院 陕西咸阳712100)
我国传统发酵乳品产区主要集中在甘肃、青海、新疆、西藏以及内蒙古等地,其特殊的地理气候环境以及牧民的生活习惯造就了独特的微生物资源[1-3]。生活在牧区的牧民利用丰富的乳品资源,以马乳、牛乳、骆驼乳和羊乳等为原料,通过传统的发酵方式制成酸牦牛奶、酸马奶、酸驼奶、奶渣等乳制品[4-5]。在发酵乳品制作过程中,伴随着以乳酸菌为主的微生物生长代谢活动[6]。相关研究表明,长期饮用酸马奶、酸驼奶等乳酸菌发酵制品可以降低人体内血清胆固醇的含量,降低人体患心血管疾病、肥胖等风险[7-8]。从不同牧区发酵乳中筛选出具有开发应用潜力的乳酸菌,是从大自然获取可利用资源的有效方式之一。
近年来,随着我国经济的快速发展,人们的生活节奏不断加快,不良生活习惯的人群比例越来越高,从而导致患冠心病、高血脂和高血压等心血管疾病的人群日益增多[9]。人体内血清胆固醇含量高是诱发心血管疾病的重要原因[10]。大量研究发现,部分乳酸菌具有降低人体内血清胆固醇的能力,关键在于其生长代谢过程中产生胆盐水解酶的活力优劣(Bile salt hydrolase,BSH)[11]。胆盐水解酶在机体内可将结合态胆盐分解为游离胆盐和相应的氨基酸产物,在酸性条件下,游离胆盐和胆固醇发生共沉淀效应;亦或是结合态胆盐,经胆盐水解酶分解后生成的游离胆汁酸不利于肠道吸收,被排出体外,导致其在肠肝系统循环中的胆汁酸量减少,肝脏利用胆固醇重新合成胆汁酸,加速了胆固醇的分解代谢,因此胆盐水解酶被认为是降解胆固醇的关键因素[12-16]。如郭春锋等[17]对乳酸菌降解胆固醇的可能机理进行探究,结果表明:在体外具有胆盐解离能力的乳酸菌,在体内亦具有潜在的降解胆固醇能力。γ-氨基丁酸(GABA)作为一类非蛋白质氨基酸,广泛存在于自然界中,是一种重要的神经递质,其在哺乳动物体内起重要作用,参与体内的代谢与生理功能的正常表达。
现有研究中,还没有对青海、甘肃、新疆、西藏和内蒙古等5 个西部牧区的传统发酵乳品中乳酸菌的胆盐解离能力的比较分析。本文综合分析上述五大西部牧区传统发酵乳品中乳酸菌胆盐解离能力,探究筛选出的优良乳酸菌产γ-氨基丁酸的能力,为乳酸菌新资源发掘,以及γ-氨基丁酸工业化生产提供参考,也为西部牧区微生物新资源的开发及合理化利用提供理论依据。
1 材料与方法
1.1 材料与试剂
1.1.1 样品来源 分离自青海、甘肃、新疆、内蒙古、西藏牧区传统发酵乳品(酸牦牛奶、酸马奶、酸驼奶和奶渣)的275 株乳酸菌。
1.1.2 培养基与试剂 培养基:MRS 肉汤培养基,北京陆桥技术有限公司。
试剂:乙腈、乙酸乙酯,美国TEDIA 有限公司;甘氨胆酸钠(GCA)、甘氨脱氧胆酸钠(GDCA)、甘氨鹅脱氧胆酸钠(GCDCA)、牛磺胆酸钠(TCA)、牛磺脱氧胆酸钠 (TDCA)、牛磺鹅脱氧胆酸钠(TCDCA)、脱氧胆酸钠标品、甲酸、磷酸、2,4-对溴苯乙酮、N,N-二异丙基乙胺,美国Sigma-Aldrich公司。
1.2 仪器与设备
1100 高效液相色谱仪,美国安捷伦公司;MD200-1 氮吹仪,山东莱索科技有限公司;DRP-9162 型电热恒温培养箱,上海森信实验仪器有限公司;YXQ-LS-50SII-01-00 立式压力蒸汽灭菌器,上海博迅实业有限公司;HC3018 型高速离心机,安徽科大中佳有限公司。
1.3 试验方法
1.3.1 乳酸菌胆盐解离能力定性分析 将待测菌株接种至MRS 肉汤培养基中于37 ℃活化2 代后,以体积分数1%的量接种到2 mL 含有0.4 mmol/L GDCA 和TDCA 的MRS 肉汤培养基中,36 ℃静置培养24 h。
待测菌株培养24 h 后,向培养基中加入甲酸,调节pH 值至2.0,混匀,再向培养基中加入6 mL 乙酸乙酯,混匀,室温静置20 min,于8 000×g离心5 min,取3 mL 上清液于氮吹管,50 ℃氮气吹干,然后用2.5 mL 4.5 g/L 2,4-对溴苯乙酮的乙腈-甲醇(体积比9∶1)溶液进行复溶,向氮吹管中加入23 μL N,N-二异丙基乙胺溶液,60 ℃水浴30 min,水浴结束后将溶液混匀冷却至25 ℃室温,取1 mL 溶液经0.45 μm 尼龙膜过滤,利用高效液相色谱法对溶液中的胆酸钠进行分析。本试验以不接菌的含GDCA 和TDCA 的MRS 肉汤培养基作为空白对照[18]。
高效液相色谱条件:色谱柱为安捷伦TC-C18(25 cm×0.46 cm×5 μm);流动相A 为乙腈-水(70∶30,体积比,磷酸调节pH 值至3.1);流动相B 为乙腈。梯度洗脱:流动相A 首先运行5 min,然后流动相B 在25 min 内由0%升高到80%,维持15 min。样品进样量20 μL,流速1 mL/min,柱温40℃,检测器为二极管阵列检测器,检测器波长为254 nm。
1.3.2 乳酸菌胆盐解离能力定量分析 对275 株乳酸菌进行胆盐解离能力定性分析,筛选出具有胆盐解离能力的阳性株,并对阳性株的胆盐解离能力进行定量分析。
将待测阳性株接种至MRS 肉汤培养基中于37 ℃活化2 代后,以体积分数1%的量接种到2 mL 含有0.4 mmol/L GCA,GDCA,GCDCA,TCA,TDCA,TCDCA 的MRS 肉汤培养基中,36 ℃静置培养24 h。
待测菌株培养24 h 后,向培养基中加入2 mL 甲醇终止反应,用1 mol/L NaOH 溶液将培养基pH 值调至6.0,混匀,于8 000×g 离心5 min,取1.0 mL 上清液经0.45 μm 尼龙膜过滤,利用高效液相色谱法对溶液中的胆盐进行分析。本试验以不接菌的含TCA,TDCA,TCDCA,GCA,GDCA,GCDCA 的MRS 肉汤培养基作为空白对照[19]。
高效液相色谱条件:色谱柱为安捷伦TC-C18(25 cm×0.46 cm×5 μm);流动相A 为含0.0075 mol/L 四丁基硫酸氢铵的乙腈-水溶液(体积比60∶40);流动相B 为含0.0075 mol/L 四丁基硫酸氢铵的乙腈-水溶液(体积比30∶70)。梯度洗脱:流动相B 的比例在30 min 内由85%下降至30%,然后在1 min 内上升至85%,维持9 min。样品进样量为40 μL,流速1 mL/min,柱温40 ℃,检测器为二极管阵列检测器,检测器波长200 nm。
待测菌株胆盐解离能力评价基于菌株培养基中TCA,TDCA,TCDCA,GCA,GDCA,GCDCA 的降解率按式(1)计算。
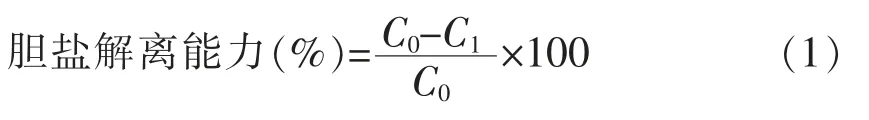
式中,C0——不接种菌株的含0.4 mmol/L TCA,TDCA,TCDCA,GCA,GDCA,GCDCA 的MRS肉汤培养基中各胆盐的浓度(mmol/L);C1——待测样品培养基中TCA,TDCA,TCDCA,GCA,GDCA,GCDCA 各胆盐的浓度(mmol/L)。
1.3.3 乳酸菌产γ-氨基丁酸能力分析 将筛选出的优势胆盐解离能力的乳酸菌阳性株于50 mmol/L L-谷氨酸钠的MRS 肉汤培养基中培养12,24,36,48 h 后,置于离心机中8 000×g 离心10 min。取100 μL 上清液于5 mL 具塞试管中,向具塞试管中加入0.45 mL 40 g/L NaHCO3溶液,充分混匀,加入0.5 mL 5 g/L 衍生试剂丹磺酰氯(用丙酮溶解丹酰氯,配制得到丹磺酰氯),充分混匀,60 ℃水浴反应30 min,水浴结束后加入100 μL 氨水,充分混匀,60 ℃水浴反应5 min 以终止衍生反应,水浴结束后加入1 mL 甲醇,振荡混匀,取1 mL 上层溶液经0.45 μm 尼龙膜过滤,通过高效液相色谱法测定溶液中的γ-氨基丁酸含量。本试验空白对照为不接菌的含有50 mmol/L L-谷氨酸钠的MRS 肉汤培养基,试验重复3 次。
高效液相色谱条件:色谱柱为安捷伦TC-C18(25 cm×0.46 cm×5 μm);检测器为紫外检测器;流动相A 为四氢呋喃-甲醇-三水合乙酸钠溶液(三水合乙酸钠浓度为0.05 mol/L,磷酸调节pH 值至6.2,流动相A 中各溶液体积比为10∶150∶840);流动相B 为甲醇。梯度洗脱为流动相B,在0~25 min内,从10%升高到45%,维持2 min,然后从27 min 到28 min 升高到90%,维持5 min,然后在1 min 内降到10%,维持7 min。梯度洗脱程序持续41 min,样品进样量15 μL,流速1 mL/min,柱温35 ℃,紫外检测器波长254 nm。
1.4 数据处理
试验重复3 次,使用SPSS 19.0 和Excel 软件对试验数据进行处理分析,结果采用平均值±标准差表示,显著性水平设定为P<0.05。
2 结果与分析
2.1 胆盐解离能力阳性株筛选
对275 株乳酸菌胆盐解离能力进行定性分析,筛选出83 株具有胆盐解离能力的乳酸菌,分为12 个乳酸菌种,分别分离自青海酸牦牛奶、新疆酸马奶、内蒙古酸马奶、新疆酸驼奶和西藏奶渣样品。具有胆盐解离能力乳酸菌菌株信息如表1所示。
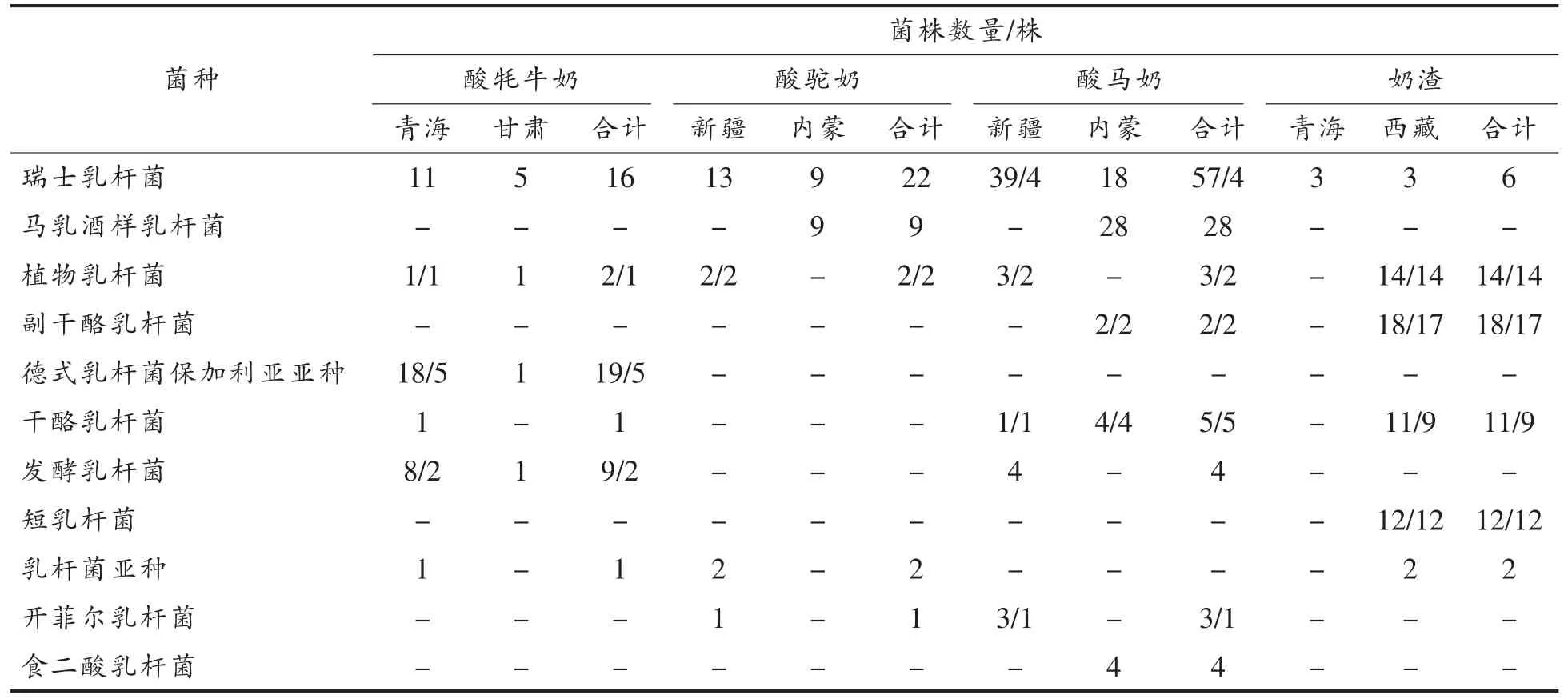
表1 解离胆盐阳性株分布情况Table 1 The information of bile salt deconjugation ability strains
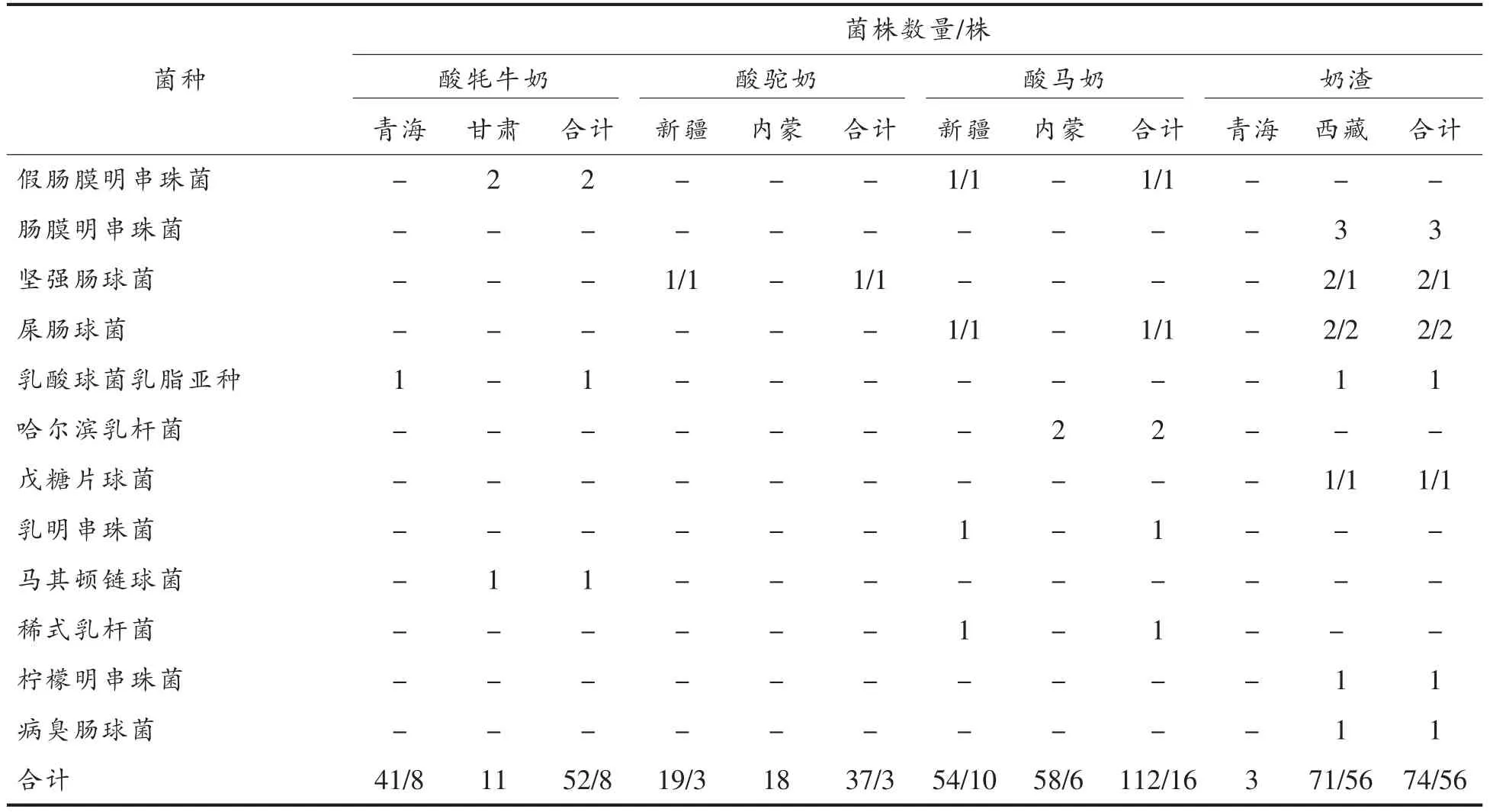
(续表1)
在23 种乳酸菌中,有12 种乳酸菌具有胆盐解离能力,分别为瑞士乳杆菌(Lb.helveticus)、植物乳杆菌 (Lb.plantarum)、副干酪乳杆菌(Lb.paracasei)、德式乳杆菌保加利亚亚种 (Lb.delbrueckii ssp.Bulgaricus)、干酪乳杆菌(Lb.casei)、发酵乳杆菌(Lb.fermentum)、短乳杆菌(Lb.breris)、开菲尔乳杆菌(Lb.kefiri)、假肠膜明串珠菌(Leuc.pseudomesenteroides)、坚强肠球菌(Ec.durans)、屎肠球菌(Ec.faecium)、戊糖片球菌(P.pentosaceus)。在275 株乳酸菌中不同菌种之间阳性株筛出率存在差异,瑞士乳杆菌阳性株筛出率为4%,副干酪乳杆菌阳性株筛出率为95%,植物乳杆菌阳性株筛出率为90.5%,屎肠球菌阳性株筛出率为100%,德式乳杆菌保加利亚亚种阳性株筛出率为26.3%,干酪乳杆菌阳性株筛出率为82.4%,短乳杆菌阳性株筛出率为100%,发酵乳杆菌阳性株筛出率为15.4%,开菲尔乳杆菌阳性株筛出率为25%,假肠膜明串珠菌阳性株筛出率为33.3%,坚强肠球菌阳性株筛出率为66.6%,戊糖片球菌阳性株筛出率为100%,故副干酪乳杆菌、植物乳杆菌、屎肠球菌、短乳杆菌、干酪乳杆菌和戊糖片球菌呈现出较高的阳性株筛出率。本研究的筛选结果与先前研究相一致,根据之前的文献报道显示,乳杆菌、肠球菌以及片球菌均具有胆盐水解酶合成能力[20-24]。
不同地区、不同传统发酵乳品分离出的乳酸菌中,阳性株筛出率存在差异。地区间比较结果显示,青海地区、甘肃地区、新疆地区、内蒙古地区、西藏地区分离出的乳酸菌中阳性株筛出率分别为18.2%,0,17.8%,7.9%,78.9%,故西藏地区分离出的乳酸菌中阳性株筛出率最高,而甘肃地区分离出的乳酸菌中未筛选出阳性株。发酵乳品品种间比较结果显示,酸牦牛奶、酸马奶、酸驼奶、奶渣分离出的乳酸菌中阳性株筛出率为15.4%,8.1%,14.3%,75.7%,故奶渣分离出的乳酸菌中阳性株筛出率最高,而酸马奶分离出的乳酸菌中阳性株筛出率最低。导致上述差异的原因可能与乳种成分之间的差异、菌种的聚类、当地独特的地理气候环境、发酵乳的制作工艺以及采样点的不同有关。
2.2 阳性乳酸菌胆盐解离能力定量分析
2.2.1 阳性株对6 种胆盐解离能力比较 将筛选出的阳性株对6 种胆盐的解离率进行方差分析,比较阳性株对6 种胆盐解离能力的差异。

图1 83 株阳性菌对6 种胆盐解离能力比较Fig.1 The comparison of six human bile salts deconjugation rate for 83 BSH-positive LAB
结果如图1所示,阳性株对6 种胆盐解离能力存在显著性差异(P<0.05),其中对GDCA 的解离率最高,对TDCA 的解离率最低。同时阳性株对GDCA 和GCDCA 的解离能力差异不显著(P>0.05),对TCA,TDCA 和TCDCA 的解离能力差异不显著(P>0.05),对GDCA 和GCDCA 的解离能力显著高于GCA,TCA,TDCA 和TCDCA (P<0.05),对GCA 的解离能力显著高于TCA,TDCA 和TCDCA(P<0.05)。综上可知,阳性乳酸菌对甘氨酸结合态胆盐的解离能力高于牛磺酸结合态胆盐。Mcauliffe 等[25]研究表明嗜酸乳杆菌NCFM 中含有bshA 和bshB 两个胆盐水解酶基因,而这两种基因对应合成的胆盐水解酶对结合态胆盐降解偏好不同,故导致上述差异的原因可能是乳源中筛分出的阳性株,所含胆盐水解酶基因表达出的胆盐水解酶对甘氨酸结合态胆盐的解离能力高于牛磺酸结合态胆盐所致。
2.2.2 不同地区发酵乳品中阳性株胆盐解离能力比较 将青海、新疆、内蒙古、西藏4 个地区传统发酵乳品中筛分出阳性株的胆盐解离率进行方差分析,比较不同地区阳性株在胆盐解离能力上的差异。
结果如图2所示,不同地区阳性株胆盐解离能力存在显著性差异(P<0.05)。从总胆盐解离结果分析可知,来源于西藏和新疆的阳性株解离能力显著高于青海和内蒙古的阳性株 (P<0.05),尤其以来源于新疆地区的阳性株对总胆盐解离能力最强,内蒙古地区的阳性株解离能力最差。分别从6 种胆盐解离结果分析可知,4 个地区的阳性株分别对TCA,TDCA 和TCDCA 的解离能力差异不显著(P>0.05),然而对GCA,GDCA 和GCDCA 的解离能力各自呈显著性差异(P<0.05)。来源于新疆的阳性株对GCA 和GCDCA 的解离能力显著高于青海与内蒙古的阳性株(P<0.05);来源于西藏的阳性株对GCA 和GCDCA 的解离能力显著高于内蒙古的阳性株(P<0.05),而与新疆和青海的阳性株解离能力差异不显著(P>0.05);来源于新疆的阳性株对GDCA 的解离能力显著高于青海与内蒙古的阳性株 (P<0.05);来源于西藏的阳性株对GDCA 的解离能力显著高于青海的阳性株(P<0.05),而与新疆和内蒙古的阳性株解离能力差异不显著(P>0.05)。不同地区的传统发酵乳品由于当地独特的地理气候环境,使其存在着复杂多样的菌群微生态结构以及特定的微生物类群,发酵乳品体系中的菌群之间存在互生现象,使得不同发酵乳品中乳酸菌的丰度存在差异,从而可能导致乳酸菌胆盐解离能力的不同。乳酸菌的遗传多样性与其分离地区之间也存在直接关系,研究表明,每个地区乳酸菌均有属于其独特的基因型,从而解释了乳酸菌解离胆盐特性因地区不同而存在差异的推论[26]。
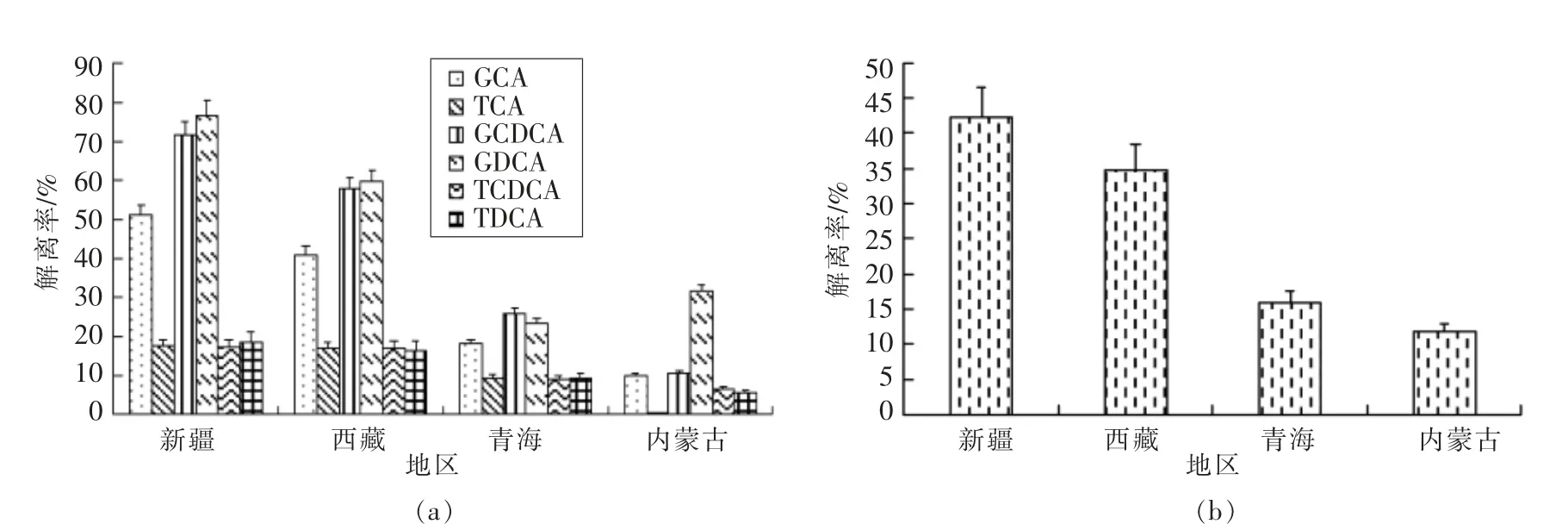
图2 不同地区阳性株胆盐解离能力比较Fig.2 Comparison of bile salt dissociation ability of positive strains in different regions
2.2.3 不同发酵乳品中阳性株胆盐解离能力比较 将酸牦牛奶、酸马奶和奶渣3 种传统发酵乳品筛分出阳性株的胆盐解离率进行方差分析,分析不同传统发酵乳品中阳性株在胆盐解离能力上的差异。
结果如图3所示,不同传统发酵乳品筛分出的阳性株胆盐解离能力存在显著性差异 (P<0.05)。从总胆盐解离结果分析可知,奶渣中的阳性株解离能力显著高于酸牦牛奶中的阳性株(P<0.05),酸马奶中的阳性株解离能力与酸牦牛奶和奶渣中的阳性株无显著性差异(P>0.05),其中奶渣中的阳性株对总胆盐解离能力最强,酸牦牛奶中的阳性株解离能力最差。从6 种胆盐解离结果分析可知,来源于3 种传统发酵乳品的阳性株对GCA,TCA,TDCA 和TCDCA 的解离能力差异不显著(P>0.05),对GDCA 和GCDCA 的解离能力呈显著性差异 (P<0.05)。来源于奶渣中的阳性株对GCDCA 的解离能力显著高于酸牦牛奶中的阳性株(P<0.05),而来源于酸马奶中的阳性株对GCDCA 解离能力与酸牦牛奶和奶渣中的阳性株无显著性差异(P>0.05);来源于酸马奶和奶渣中的阳性株对GDCA 解离能力显著高于酸牦牛奶中的阳性株(P<0.05)。不同传统发酵乳品中乳酸菌胆盐解离能力存在差异,可能由于不同物种之间各自生存条件、牧民的喂养方式以及生长环境不同,导致其乳中干物质等营养成分存在差异,使得乳中乳酸菌群落结构和所表达出的胆盐解离能力不同。Sun 等[27]采用多位点序列分型(Multilocus sequence typing,MLST)技术对分离自我国和蒙古国的瑞士乳杆菌遗传多样性进行了系统分析,指出菌株的聚类与乳源有关,即酸马奶、酸牛奶和酸牦牛奶中分离出的乳酸菌形成单独的类群使其菌株的特性存在普遍的差异;Song 等[26]利用MLST 技术对中国、俄罗斯及蒙古国等地的传统发酵乳品中德式乳杆菌保加利亚亚种进行微进化分析,研究证实了乳酸菌会因为适应其所处环境而发生某种特定进化的假说,显示出分离菌株为适应特定的生存环境而发生某种进化的历程。除此之外,乳的发酵前处理以及发酵乳的制作方式不同也会导致乳酸菌胆盐解离能力的差异。
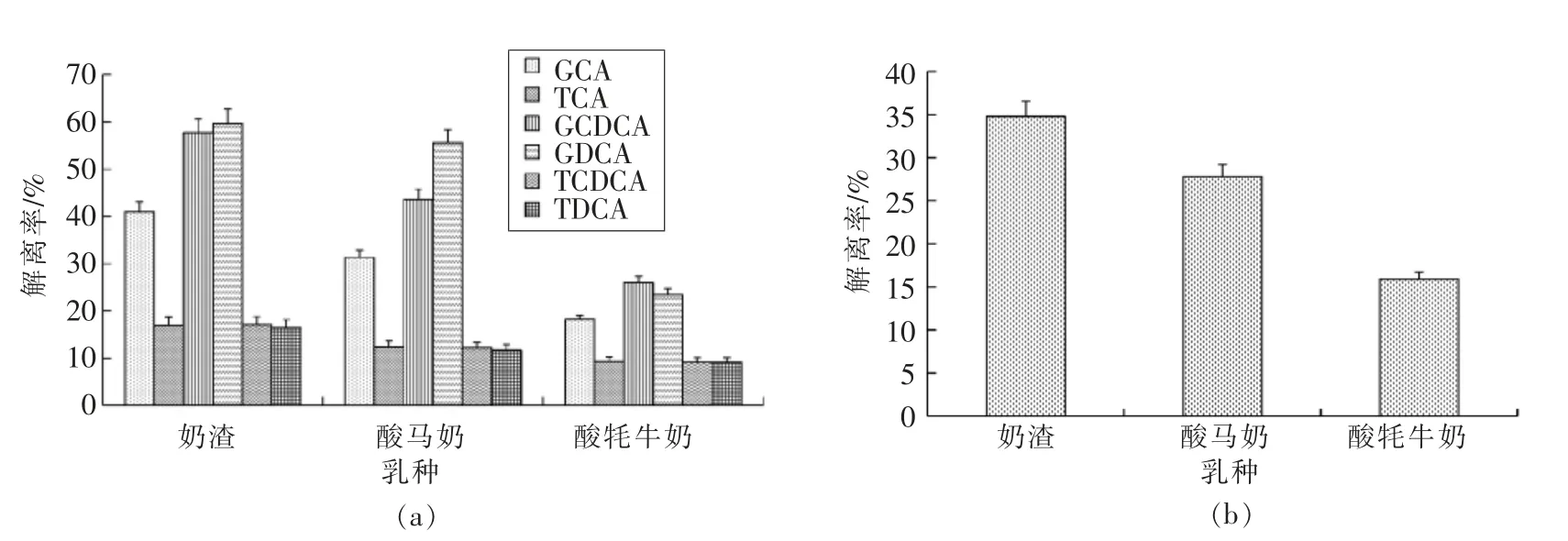
图3 不同发酵乳品中阳性株胆盐解离能力比较Fig.3 Comparison of bile salt dissociation ability of positive strains in different fermented dairy products
2.2.4 不同种类优势阳性株胆盐解离能力比较 对4 种优势阳性乳酸菌的胆盐解离率进行方差分析,分析不同优势阳性菌种在胆盐解离能力上的差异。
结果如图4所示,4 种不同优势阳性菌种之间胆盐解离能力存在显著性差异(P<0.05)。从总胆盐解离结果分析可知,植物乳杆菌和短乳杆菌的胆盐解离能力显著高于副干酪乳杆菌和干酪乳杆菌(P<0.05),短乳杆菌对总胆盐解离能力最强,干酪乳杆菌解离能力最差。从6 种胆盐解离结果分析可知,4 种阳性菌种分别对TCA 和TCDCA 的解离能力差异不显著(P>0.05),然而对GCA,GDCA,TDCA 和GCDCA 的解离能力呈显著性差异(P<0.05)。植物乳杆菌和短乳杆菌对GDCA 和GCDCA 的解离能力显著高于副干酪乳杆菌和干酪乳杆菌(P<0.05);短乳杆菌对GCA 的解离能力显著高于植物乳杆菌、副干酪乳杆菌和干酪乳杆菌(P<0.05),植物乳杆菌对GCA 的解离能力显著高于副干酪乳杆菌和干酪乳杆菌(P<0.05);短乳杆菌和副干酪乳杆菌对TDCA 的解离能力显著高于干酪乳杆菌(P<0.05),而植物乳杆菌对TDCA的解离能力与短乳杆菌、副干酪乳杆菌和干酪乳杆菌差异不显著(P>0.05)。不同菌种在胆盐解离能力上的差异可能由于菌种自身所具有的特性间存在相互影响,不同种类乳酸菌产胆盐水解酶能力不同,导致其胆盐解离能力存在差异;不同种类乳酸菌在基因水平上的不稳定性以及其表达出胆盐水解酶活力的不稳定,都可能导致其胆盐解离能力不同。除此之外,不同种类乳酸菌自身所携带的胆盐水解酶基因具有明显差异,研究表明嗜酸乳杆菌 NCFM 中含有bshA 和bshB 两个胆盐水解酶基因,而植物乳杆菌中含有bsh1,bsh2,bsh3,bsh4 共4 个胆盐水解酶基因[28-30],不同基因所表达出的胆盐水解酶活性存在差异,表现出对6 种胆盐的解离能力不同,故不同菌种间所含胆盐水解酶基因的不同也是导致其胆盐解离能力差异的原因之一。
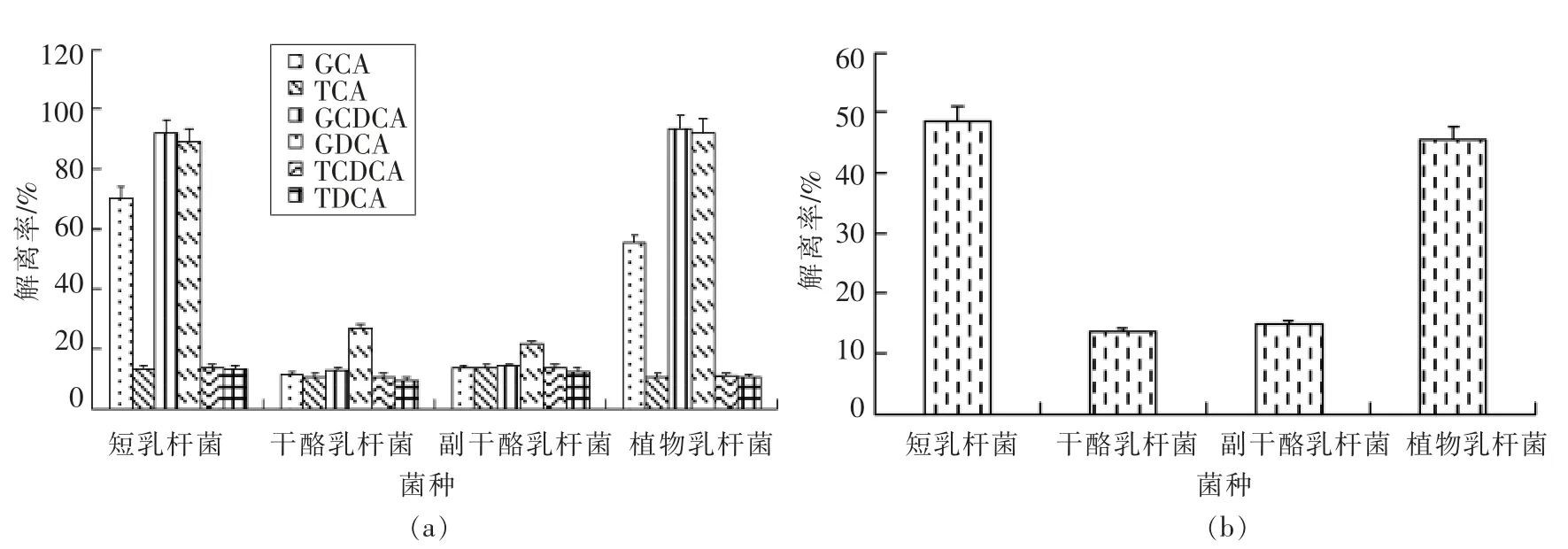
图4 不同种类优势阳性株胆盐解离能力比较Fig.4 Comparison of bile salt dissociation ability of different dominant positive strains
2.2.5 对筛选出不同优势阳性株产γ-氨基丁酸能力研究 对筛选出4 种优势阳性乳酸菌的产γ-氨基丁酸能力进行产量测定,通过结果分析不同优势阳性菌种在产γ-氨基丁酸能力上的差异。
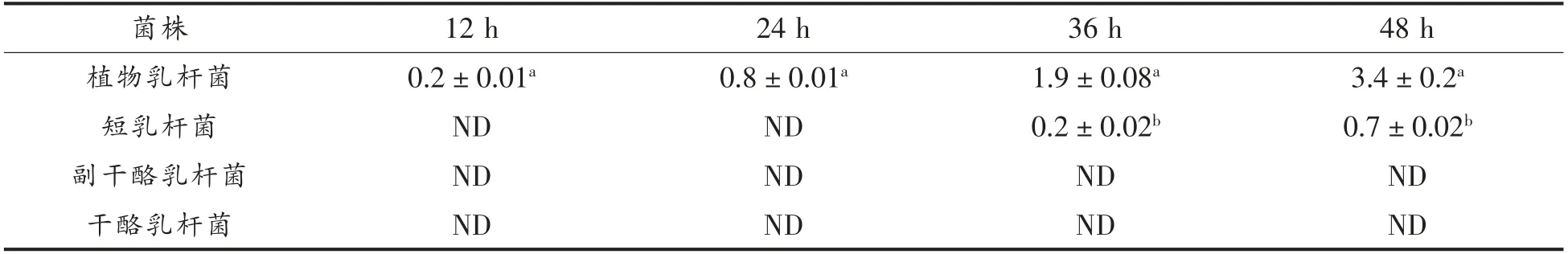
表2 乳酸菌的γ-氨基丁酸产量测定结果(mmol/L)Table 2 Determination of γ-aminobutyric acid production of lactic acid bacteria (mmol/L)
由表2可知,优势阳性株的γ-氨基丁酸的产量随着培养时间的延长而不断增加,在培养48 h后产量达到峰值。4 种阳性菌中γ-氨基丁酸的产量最高为新疆酸马奶的植物乳杆菌,产量达(3.4±0.2)mmol/L,其次为西藏奶渣中的短乳杆菌,产量为(0.7±0.02)mmol/L,而副干酪乳杆菌和干酪乳杆菌不能产γ-氨基丁酸。乳酸菌具备产γ-氨基丁酸能力主要因其在生长代谢过程中,会产生谷氨酸脱羧酶与谷氨酸发生催化脱羧反应产生γ-氨基丁酸,故不同乳酸菌种之间产谷氨酸脱羧酶能力不同,也可导致其产量的差异;不同来源的乳酸菌因其地区、乳源以及发酵乳品制作工艺的不同,使得菌种在基因表达方面上存在差异,从而导致菌株产量不同。
大量研究表明短乳杆菌、植物乳杆菌、副干酪乳杆菌等均具有产γ-氨基丁酸的能力,且产量较高。本研究中筛选出的植物乳杆菌产γ-氨基丁酸能力相比于其它纯培养条件下处于较高水平。Di等[31]从干酪中分离出一株植物乳杆菌,在MRS 纯培养条件下,γ-氨基丁酸的产量为498.1 mg/L。本试验筛选出4 种优良菌株产γ-氨基丁酸能力分析:短乳杆菌和植物乳杆菌有较高产γ-氨基丁酸能力,而副干酪乳杆菌和干酪乳杆菌不产γ-氨基丁酸,同时基于本实验室之前的研究发现[32],短乳杆菌和植物乳杆菌呈优势胆盐解离能力、高产γ-氨基丁酸能力的同时也具备高降解亚硝酸盐能力,可进行下一步性能研究[33],为优良乳酸菌种资源的开发提供理论依据。
3 结论
1)从275 株乳酸菌中筛选出83 株具有胆盐解离能力的乳酸菌,分为12 个乳酸菌种,分离自青海酸牦牛奶、新疆酸马奶、内蒙古酸马奶、新疆酸驼奶和西藏奶渣样品。菌种中副干酪乳杆菌、植物乳杆菌、短乳杆菌、屎肠球菌、戊糖片球菌和干酪乳杆菌呈现出较高的阳性株筛出率,菌株来源中西藏奶渣呈现出较高的阳性株筛出率。
2)阳性株对6 种结合态胆盐的解离能力存在显著性差异(P<0.05),其中对GDCA 解离率最高,对TDCA 解离率最低。阳性株对甘氨酸结合态胆盐的解离能力高于牛磺酸结合态胆盐。
3)区域间比较显示:不同地区阳性株对GCA,GCDCA,GDCA 和总胆盐解离能力存在显著性差异(P<0.05);乳种间比较显示:不同乳种阳性株对GCDCA,GDCA 和总胆盐解离能力存在显著性差异(P<0.05);优势菌种间比较显示:不同种类阳性株对GCA,GCDCA,GDCA,TDCA 和总胆盐解离能力存在显著性差异(P<0.05)。由此可见,胆盐解离能力因菌株来源和种类不同存在差异性,以新疆酸马奶、酸驼奶和西藏奶渣中的阳性株胆盐解离能力表现最优,其中短乳杆菌、植物乳杆菌显示了较强的胆盐解离能力。
4)4 种优良胆盐解离能力菌株产γ-氨基丁酸能力分析:短乳杆菌和植物乳杆菌有较高产γ-氨基丁酸能力,而副干酪乳杆菌和干酪乳杆菌不产γ-氨基丁酸。综上所述,胆盐解离能力因菌株来源和种类不同存在差异性,以新疆酸马奶、酸驼奶和西藏奶渣中的阳性株胆盐解离能力表现最优,其中短乳杆菌、植物乳杆菌显示了较强的胆盐解离能力,同时具有高产γ-氨基丁酸能力,可为优良乳酸菌种资源的开发提供理论依据。
