耳鸣相关基因研究进展
2020-08-26于澜王秋菊
于澜 王秋菊
1解放军总医院耳鼻咽喉头颈外科医学部(北京100853)
2国家耳鼻咽喉疾病临床医学研究中心(北京100853)
1 引言
耳鸣是指在没有外界声源或电刺激存在的情况下,患者自觉耳内或颅内有声音的一种主观症状,是一种内在声音的感知而不是外界存在的声源[1]。据统计,人群中约10%-30%有耳鸣感受,其中多数可耐受耳鸣,但少数患者的生活受到严重影响[2]。《耳鸣临床应用指南》中提到,美国有超过5000万人抱怨耳鸣,发生率为10%~15%[3]。耳鸣易影响睡眠、注意力、社会交往和工作[4],还易使患者产生神经衰弱、焦虑、忧郁的现象[5]。耳鸣作为一种常见的主观症状困扰着人们的学习、工作和生活,深入了解耳鸣的病因和病理机制有助于对耳鸣进行诊断和干预。
耳鸣是耳鼻喉科最常见的主诉之一,其病因包括环境因素和遗传因素[6]。随着遗传学研究的重大突破和快速发展,耳鸣的遗传因素也逐渐受到学术界的关注。遗传易感性可能对某些患者耳鸣的起源有着重要作用,而非遗传因素可能在决定有遗传易感的受试者是否会发展为耳鸣方面起主导作用[7]。耳鸣受心理、耳部疾病、遗传因素等综合影响,本文主要介绍与耳鸣有关的遗传基因研究进展。
2 流行病学
2004年,文献报道与听力下降相关联的耳鸣提示可能为家族性耳鸣[8]。2007年,1147例比利时耳鸣患者的研究发现家族性耳鸣与平坦型听力图相关[9]。2007年,Hendrickx等[10]在一项涉及7个欧洲国家、198个家庭的多中心研究中报道,来自于有耳鸣家庭的个体的耳鸣发生率是来自于无耳鸣家庭个体的1.7倍。但Kvestad等[11]在2010年对一个挪威大家庭的研究,显示耳鸣只有0.11的遗传率。2017年比利时的研究小组利用复杂性状遗传分析(Genetic Analysis of Complex Traits,GACT)软件,估计全基因组关联分析(Genome Wide Association Study,GWAS)中所有单核苷酸多态性(singlenucleotide polymorphisms,SNPs)可解释3.2%的变异。这项GWAS研究还发现了一些可能与耳鸣有关的通路(如氧化应激、内质网应激和5-羟色胺受体介导的信号传递)[12]。如果一种性状或疾病在同卵双胞胎中,同时发生的概率高于异卵双胞胎,那么这种性状或疾病是可遗传的。2017年瑞典一项多中心研究选取了10,000对双胞胎登记数据进行研究,发现耳鸣在同卵双胞胎中发生率高于异卵,说明了遗传基因可能在耳鸣中起着重要作用,尤其强调了双侧(0.56)耳鸣的遗传性,男性双侧耳鸣的遗传性为0.68[7]。
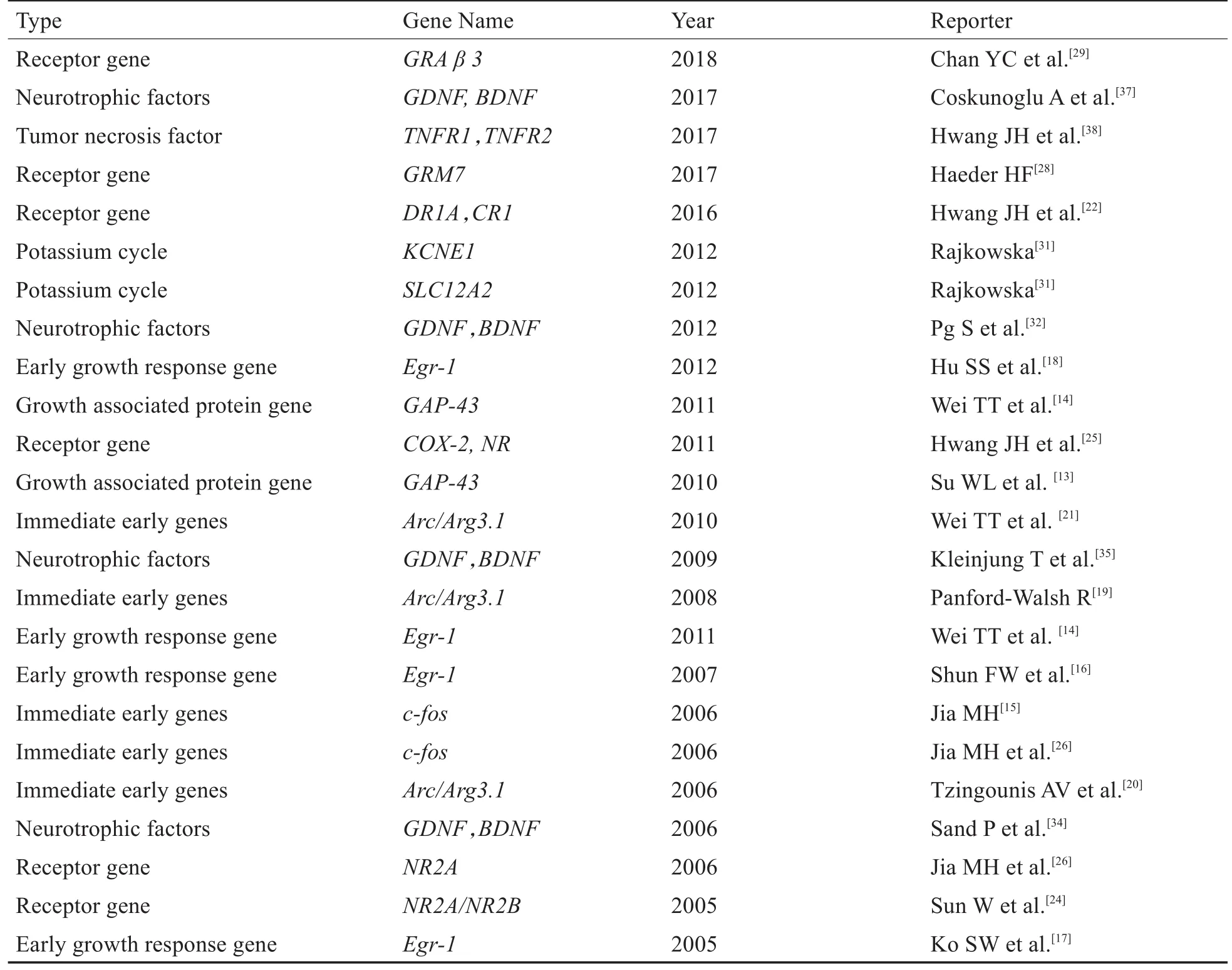
表1 耳鸣遗传基因相关的研究Table 1 Studies on correlated gene of tinnitus
3 遗传相关性研究
耳鸣可作为唯一主诉出现,也可能伴随其他症状和疾病,例如梅尼埃病和老年性聋等。在一些个体中,耳鸣逐渐发展,而在另一些个体中耳鸣突然发作,这种个体对耳鸣的易感性可以通过不同环境和内在因素的影响来解释,包括遗传因素[10]。耳鸣不论是产生于外周还是中枢,异常信号必须上传到大脑皮层才能被感知为耳鸣,即听觉中枢特别是大脑皮层参与了耳鸣的产生与维持。本文列举了近些年与耳鸣相关的基因研究,见表1。
3.1 生长相关蛋白基因
检测神经元功能可塑性基因GAP-43属于生长相关蛋白(growth associated protein,)中的一种,广泛分布于大脑、小脑、海马以及脊髓、背根神经节和自主神经系统。体外培养脊髓神经元的结果表明,GAP-43的表达与神经元的发育和可塑性有直接关系[13]。对水杨酸钠致耳鸣的大鼠研究中显示[14],耳鸣大鼠听皮层中GAP-43蛋白阳性神经元的表达明显高于非耳鸣大鼠,说明水杨酸钠组大鼠听皮层产生了新的连接,在大鼠耳鸣的维持中起到了重要作用。
3.2 即刻早期基因
即刻早期基因(Immediate early genes,IEGs)cfos,能在静止细胞受到各种刺激时迅速表达。研究显示[15],耳鸣大鼠的c-fos基因表达增多,且FOS蛋白在耳鸣模型大鼠听皮层中的表达显著增高,说明耳鸣大鼠出现神经元的异常电活动以及神经元功能活动增加。从研究结果看来,FOS蛋白不仅参与了耳鸣的发生,也反映了耳鸣动物听皮层中神经电活动的异常和功能可塑性改变。
早期生长反应基因-1(Early growth response-1,Egr-1)也是一种即刻早期基因。研究表明,突触可塑性常表现为长时程增强(long-term potentiation,LTP),而早期生长反应基因Egr-1的表达能作为引发LTP的分子标识(LTP常常伴随着Egr-1表达的升高)[16]。研究发现[17],用低强度的短阵脉冲刺激(theta burst stimulation,TBS)正常小鼠听皮层后可引发持续40分钟的突触反映的长时程增强,而在Egr-1基因敲除小鼠,给予相同刺激时听觉皮层所引起的LTP被明显减弱或完全阻断,说明Egr-1在神经元活动中维持着可塑性的改变。赵德安[14]等对耳鸣大鼠模型的研究中显示听皮层和下丘Egr-1的表达水平显著增高,而耳蜗核及上橄榄核中,随着水杨酸钠的注射次数的增多,Egr-1的mRNA表达水平总体出现下降趋势。黄治物[18]等研究显示,慢性注射水杨酸后大鼠听皮层Egr-1表达显著下降。
3.3 细胞骨架活性调节基因
细胞骨架活性调节(activity regulated cytoskeleton,ARC)基因是一种可被不同形式的神经活性迅速诱导的即刻早期基因,ARC基因的动态表达对于听觉神经元持续产生并精确调控突触强度和细胞兴奋性有着重要的作用[19],因此,ARC表达水平的降低对于听觉脑干神经元突触可塑性具有不利影响[20]。研究显示[13,21],耳鸣大鼠下丘神经元中ARC的mRNA表达水平显著上升,说明ARC蛋白参与了耳鸣大鼠听皮层的重塑。
3.4 受体基因
耳鸣最常见的是源于脑干水平、皮质下区域(包括边缘系统)和皮层区域的中枢神经系统功能障碍。研究显示很多神经递质和受体也都与耳鸣相关。有学者[22,23]对水杨酸诱导耳鸣的大鼠行多巴胺受体(dopamine receptor,DR)和大麻素受体(cannabinoid receptor,CR)的研究,结果显示,水杨酸诱导耳鸣可能与耳蜗和许多耳鸣相关脑区DR1A基因mRNA表达增加有关,但与CR1基因mRNA表达减少有关。研究表明,N-甲基-D-天冬氨酸受体(N-methyl-D-aspartate Receptor,NMDAR)的活化在控制或影响突触可塑性中起重要作用[24]。关于环氧合酶(COX)和NMDAR的基因表达,Hwang等研究表明[25],耳鸣小鼠在耳蜗和中脑中,COX-2的mRNA表达略有下降,但NR2B的mRNA表达增加。贾明辉等报道,大鼠听皮层的正常发育与NMDAR的亚型NR2A的表达升高有关,而耳鸣大鼠听皮层中NR2A的表达无论在数量上还是染色体强度上都显著提高[15]。Jia等[26]发现耳鸣小鼠听觉皮层的NR2A基因表达增加,认为高剂量水杨酸引起耳鸣并且NMDAR基因表达沿着听觉途径增加。Puel等[27]发现耳鸣患者内耳的NMDAR功能和表达增强。谷氨酸代谢受体7(Glutamate metabotropic receptor 7,GRM7)可能与耳鸣严重程度有关[28]。γ-氨基丁酸(GABA)受体的功能与耳鸣有关,Chan YC等[29]研究结果显示,钝顶螺旋藻(Spirulina platensis,SP)可以增加GRAβ3的基因表达来减轻水杨酸盐导致的耳鸣。螺旋藻可降低水杨酸诱导耳鸣中KCC2基因的表达[30]。
3.5 钾循环途径相关基因
关于钾循环途径与耳鸣的相关性,Rajkowska等[31]对128例耳鸣患者和498例正常人研究显示,钾循环途径中的相关基因是引起耳鸣的潜在性因素,其中KCNE1和SLC12A2基因的2个变体存在显著关联:KCNE1与独立于听力损失的耳鸣相关,SLC12A2与噪声诱发的听力损失有关。水杨酸诱导耳鸣可能与KCC2基因mRNA表达增加有关,但与耳蜗及部分耳鸣相关脑区NKCC1基因mRNA表达无关。
3.6 神经营养因子和肿瘤坏死因子
胶质细胞源性神经营养因子(Glial cell line-Derived Neurotrophic Factor,GDNF)和脑源性神经营养因子(Brain Derived Neurotrophic Factor,BDNF)在中枢听觉通路和内耳的早期发展中起着关键作用[32,33]。研究显示[34,35],GDNF和BDNF与耳鸣存在正相关,在BDNF发生错义突变的携带者中,与听力损失关联的耳鸣发生风险显著降低。神经营养因子与皮质和海马的神经再生缺陷有关系[36]。在耳鸣病因学中,BDNF等神经营养因子扮演着重要角色。但据Coskunoglu A等报道,BDNF基因多态性与耳鸣无相关性[37]。关于肿瘤坏死因子(Tumor Necrosis Factor,TNF)阻滞剂对耳鸣动物模型耳蜗炎性细胞因子基因表达的影响,Hwang JH等发现治疗组TNFR1和TNFR2的mRNA水平均明显低于对照组,提出神经炎症可能是水杨酸诱导耳鸣的一种新的机制[38]。
4 小结
耳鸣作为耳科学研究热点一直备受人们关注。由于耳鸣是一种主观症状,目前缺乏客观有效的评估方法,再加上其病因及病理机制的复杂性,耳鸣患者的治疗效果并不十分理想。近十余年来,随着遗传学的发展,各国研究学者陆续开展了耳鸣与基因相关的研究。本文将目前报道的与耳鸣相关的基因研究进行了总结回顾,望为之后的耳鸣研究提供一些参考。本文列举了近些年与耳鸣相关的基因研究,研究主要涉及耳鸣与生长相关蛋白、即刻早期基因、细胞骨架活性调节基因、受体基因、钾循环途径相关基因和神经营养因子等。然而,尽管近几年的数据强调了遗传因素在耳鸣中的重要作用,但缺乏与任何特定遗传位点的强相关性,并且缺少具有统计学意义的遗传学研究[39]。有些基因被认为是耳鸣的候选基因,但后续研究没有发现或未能证实先前报道的相关性,缺乏可重复性的主要原因可能是群体分层或病例与对照之间的差异。研究对象的选择可能产生选择偏倚、假阳性或无意义的结果。耳鸣常被作为单一临床症状进行研究,而这些耳鸣患者可能属于不同亚型,其潜在机制可能不尽相同。耳鸣相关的基因变异可能也会导致听力下降或焦虑等,即基因的多效性[40]。
综上,耳鸣易受多种因素影响,且致病机制并未形成完善统一的认识,相关基因导致耳鸣的证据还不足,耳鸣的致病基因和遗传模式需进一步研究和探讨,人类耳鸣相关基因的研究目前仍处于起步阶段。已有流行病学研究通过同卵和异卵双胞胎多年的随访结果支持耳鸣具有遗传性。下一步需要根据表型对受试者进行仔细精确的选择,制定规范的纳排标准和选择合适的评估量表,将有助于识别耳鸣基因或与耳鸣相关合并症的基因[41]。耳鸣候选基因的鉴定可为耳鸣的诊断和治疗提供依据,使耳鸣的个性化干预做到精准有效。
