切花菊优香不同生长阶段光合作用的变化
2020-05-19卢珍红韩福贵李绅崇李进昆杨春梅
卢珍红,韩福贵,李绅崇,黎 霞,桂 敏,李进昆,杨春梅
(1.云南省农业科学院花卉研究所,云南昆明650100;2.中国科学院植物研究所,江苏南京210014)
菊花(Chrysanthemum grandiflora)是我国十大传统名花之一,也是世界四大鲜切花之一,全世界每年切花菊的产量约占切花总量的1/3[1]。虽然菊花起源于我国,但是菊花的切花生产却在欧美等发达国家率先得到普及和发展,直到21 世纪初我国切花菊产业才开始飞速发展,目前已成为增长最快的出口创汇花卉产品之一[2]。
我国切花菊产品的输出国主要是日本和韩国。尤其是日本,每年切花菊的消费量稳定在30 亿支以上[3]。每年8 月(日本盂兰盆节)和9 月(日本彼岸节)则是日本切花菊消费量的高峰。但在夏季,高温影响了切花菊的营养生长及花芽分化和发育,造成花朵畸形[4]。切花菊品种优香来源于日本,是深受日本、韩国消费者喜爱的白菊品种,花径大、花型饱满、花期持久,尤其是抗病性强、耐运输的优点使其在夏季切花菊市场上一直独秀[5],也是近几年我国推广面积种植较大的白菊品种。优香的自然花期在7 月下旬,通过花期调控技术可使其在需求高峰期7—9 月开花。因此,目前关于优香的研究主要集中在促控栽培[3-8]和花期调控[9-10]等方面。但在促控栽培和花期调控中所涉及的补光、遮阳的措施势必会影响其光合作用,进而影响其生长发育。
本研究以切花菊优香为试验材料,选取营养生长中期、营养生长后期和营养生长向生殖生长的过渡期,研究植株不同生长阶段、不同部位叶片光合作用的特点和规律,为优香的优质高效栽培提供试验依据。
1 材料和方法
1.1 试验材料
以切花菊优香3 个生长阶段的扦插苗为试验对象。所有的供试植株均来源于云南省农业科学院花卉研究所试验苗圃。其中,营养生长中期植株质量为18.80 g,高36.83 cm,叶片平均质量为10.80 g,茎平均质量为6.30 g,根平均质量为1.70 g;营养生长末期植株质量为28.50 g,高42.67 cm,叶片平均质量为16.10 g,茎平均质量为9.70 g,根平均质量为1.70 g;营养生长向生殖生长过渡期质量为32.70 g,高52.37 cm,叶片平均质量为18.70 g,茎平均质量为10.20 g,根平均质量为3.80 g。
1.2 试验方法
试验于2014 年4 月进行。营养生长中期植株测定10.00 cm(上部叶)和26.00 cm(下部叶)处叶片的光合参数;营养生长后期植株测定14.00 cm(上部叶)和28.00 cm(下部叶)处叶片的光合参数;营养生长向生殖生长过渡期植株测定17.00 cm(上部叶)和34.00 cm(下部叶)处叶片的光合参数。测量当天最高温度25 ℃,最低温度19 ℃。采用GFS-3000 型高级光合作用测量系统(Heinz Walz GmbH,Germany)的开放式气路自动测定光合- 光响应曲线。测定过程中,利用自带光源先给叶片提供700 μmol/(m2·s)光强,适应30 min 后,测定优香不同叶位叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)等光合生理参数。
在晴朗天气9:30—11:30,随机选择不同叶位的健康叶片,使用GFS-3000 型便携式光合作用测量仪,采用3040-L 红蓝光源,设定气体流流速为750 μmol/s,控制叶温(25±1)℃,CO2浓度为(380±5)μmol/(m2·s),设置光照强度(PAR)梯度为0、50、100、150、200、400、600、800、1 100、1 400、1 700、2 000 μmol/(m2·s)。测定前使用1 500 μmol/(m2·s)的PAR 对叶片进行光诱导,使叶片活化。测定优香不同叶位的Pn 值,每个光强下稳定2~3 min,待稳定时记录数据,每个叶位测定3 片叶片,用于光响应曲线的拟合。
1.3 数据分析
试验数据利用Excel 2007 进行计算汇总,获得各部位叶片的光合作用日变化参数;利用SPSS 软件,采用直角双曲线修正模型拟合光响应曲线,根据曲线方程计算出最大净光合速率(Pnmax)、暗呼吸速率(Rd)、光补偿点(LCP)以及光饱和点(LSP);表观量子效率(AQE)是光合有效辐射(PAR)(<200 μmol/(m2·s))与Pn 拟合直线的斜率。用SPSS软件进行光响应曲线的拟合。R 语言作图,作图过程参考了Plantecophys 包的部分代码[11]。
2 结果与分析
2.1 切花菊优香不同生长阶段光合作用气体交换参数的比较
从表1 可以看出,优香不同生长阶段上部叶的光合作用参数高于下部叶。上部叶的蒸腾速率(Tr)随株龄的增加而增强,即营养生长向生殖生长过渡期的蒸腾作用最强,蒸腾速率达3.74 mmol/(m2·s),为营养生长中期的2.58 倍;气孔导度(Gs)、净光合速率(Pn)和胞间CO2浓度(Ci)则均为营养生长后期最高,此时的Gs、Pn 和Ci 分别达409.97 mmol/(m2·s)、11.05 μmol/(m2·s)和394.03 μmol/L,分别是营养生长中期的4.22 倍、3.95 倍和1.28 倍,是营养生长向生殖生长过渡期的2.18 倍、1.99 倍和1.09 倍。与上部叶略有不同,优香3 个生长阶段下部叶的4 个光合参数值均为营养生长后期最大,Tr、Gs、Pn 和Ci 分别达到1.89 mmol/(m2·s)、163.09 mmol/(m2·s)、4.53 μmol/(m2·s)和431.92 μmol/L,分别为营养生长中期的1.29 倍、2.07 倍、2.43 倍和1.31 倍,是营养生长向生殖生长过渡期的1.07 倍、1.87 倍、1.78 倍和1.17 倍。
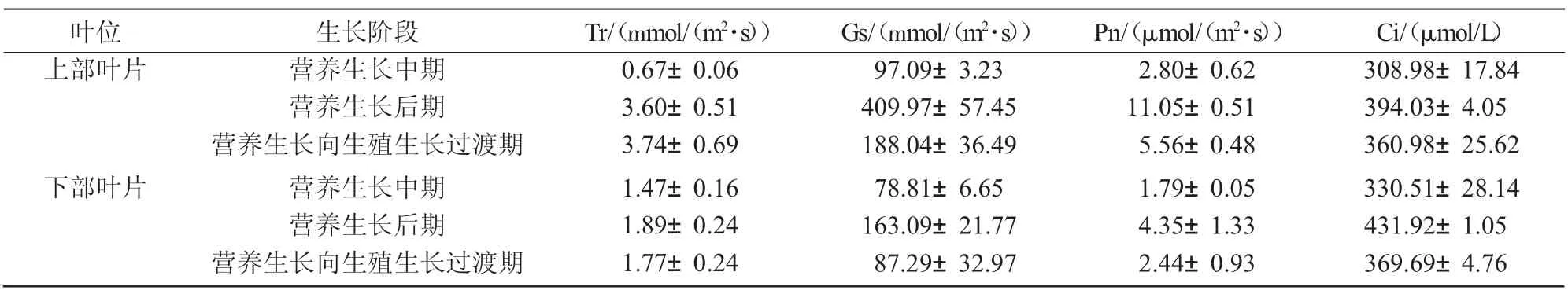
表1 切花菊优香不同生长阶段不同叶位叶片光合参数比较
2.2 切花菊优香不同生长阶段光响应曲线的比较
从图1 可以看出,优香上部叶和下部叶不同生长阶段的光响应曲线基本一致。在营养生长中期,当光照强度(PAR)小于250 μmol/(m2·s)时,Pn 随着PAR 的增强迅速增加,此时PAR 是叶片光合作用的主要限制因子;当PAR>250 μmol/(m2·s)时,随着PAR 的增大,Pn 上升速度开始减慢,并逐渐趋于平缓,达到光饱和状态。在营养生长后期,Pn 随着PAR 快速增加的范围扩大到0~600 μmol/(m2·s);当优香植株发育到营养生长向生殖生长的过渡期时,Pn 随PAR 快速增加的范围缩小为0~400 μmol/(m2·s)。可见,在不同的生长阶段,优香的光合能力变化显著。
2.3 切花菊优香不同生长阶段光响应曲线参数的比较
从表2 可以看出,不同生长阶段、不同叶位叶片光响应曲线拟合的R2均达到了0.90 以上,拟合效果较好。不同生长阶段上部叶的最大净光合速率(Pnmax)差异较大,下部叶的Pnmax 差异相对较小;上部叶营养生长后期Pnmax 达20.83 μmol/(m2·s),约为营养生长中期的4 倍,是营养生长向生殖生长过渡期的1.5 倍;下部叶营养生长后期Pnmax 为13.02 μmol/(m2·s),是营养生长中期的2.76 倍,是营养生长向生殖生长过渡期的2.47 倍。
与Pnmax一样,优香上部叶表观量子效率(AEQ)也高于下部叶。上部叶中,营养生长向生殖生长过渡期的AEQ 值最大,为0.04,营养生长中期和后期分别为0.02 和0.03。下部叶中,营养生长后期和营养生长向生殖生长过渡期的AEQ 均为0.02,营养生长中期为0.01。

表2 切花菊优香不同生长阶段不同叶位叶片光响应参数比较
优香上部叶营养生长后期的暗呼吸速率(Rd)达3.91 μmol/(m2·s),明显高于营养生长中期和营养生长向生殖生长过渡期,分别为二者的2.38 倍和1.98 倍。但下部叶暗呼吸速率随着株龄的增加呈现增加的趋势,因此,营养生长向生殖生长过渡期的Rd 最大,为1.20 μmol/(m2·s),营养生长中期和后期Rd 值分别为0.20、0.82 μmol/(m2·s)。
除营养生长中期外,优香上部叶营养生长后期和营养生长向生殖生长过渡期的光饱和点(LSP)高于下部叶。其中,营养生长后期上部叶的LSP 高达1 057.99 μmol/(m2·s),是营养生长中期和营养生长向生殖生长过渡期的3.34 倍和2.87 倍;下部叶的营养生长后期的LSP 为565.18 μmol/(m2·s),明显高于营养生长中期的476.63 μmol/(m2·s)和营养生长向生殖生长过渡期的353.93 μmol/(m2·s)。但优香不同生长阶段、不同叶位光补偿点(LCP)的差异与LSP 不同。营养生长向生殖生长过渡期的上部叶的LCP 低于下部叶;营养生长中期和后期则为上部叶的LCP 高于下部叶。但总体来说,上部叶中,营养生长向生殖生长过渡期的LCP 最低,为20.97 μmol/(m2·s),约为营养生长后期LCP 的1/4;下部叶中,营养生长中期的LCP 最低,仅为11.52 μmol/(m2·s),不足营养生长后期的1/4。
α 快速光曲线的初始斜率,代表了叶片的光化学效率。由于拟合模型的原因,营养生长向生殖生长过渡期的叶片,未获得α 值。下部叶营养生长中期和后期的α 值相同,均为0.02;但上部叶营养生长中期的α 值为0.08,略高于营养生长后期。
3 结论与讨论
植物在不同的生长阶段、不同成熟度、不同叶位的叶片光合作用参数差异显著[12-14]。同时,由于所处的位置不同,冠层的微环境也会影响叶片的光合作用[15]。在本试验选取的优香3 个生长阶段中,无论上部叶还是下部叶,营养生长后期的各光合参数(Tr、Gs、Pn 和Ci)均高于其他2 个时期。其中,上部叶Gs 表现最为明显,营养生长后期的Gs 为营养生长中期的4.22 倍;Tr 值的差异最小,下部叶营养生长后期Tr 值为营养生长向生殖生长过渡期的1.07 倍。这与优香生长发育的特点相吻合。在营养生长后期,营养生长基本完成,光合效率也达到最大值,为进入生殖生长即开花提供了充足的物质储备。
直角双曲线修正模型拟合光响应曲线结果表明,不同生长阶段、不同叶位叶片光响应曲线拟合的R2均达到了0.90 以上,拟合效果较好。从光响应曲线参数的比较结果来看,不同生长阶段上部叶的最大净光合速率(Pnmax)差异较大,下部叶的Pnmax差异相对较小,营养生长后期的Pnmax 最大。郑玉红等[16]对神马花前期和蕾后期不同叶位叶片的光合作用特点进行了研究,结果表明,神马花前期植株上部叶的Pnmax 明显高于蕾后期;下部叶片的P nmax 值则十分接近。这与本研究的结果相似。余倩花等[17]于9 月至次年3 月测定了神马、三色白等25个切花菊品种的光合速率,结果表明,夏菊品种八月黄(中大菊)6 个月Pn 的平均值为5.03 μmol/(m2·s),低于优香上部叶3 个生长阶段的Pn 值平均值6.47 μmol/(m2·s),表明从光合作用的角度看,优香确为优良的夏菊品种。
本研究中,优香营养生长后期的暗呼吸速率(Rd)和光饱和点(LSP)远远高于其他2 个时期,说明随着生长发育的进行,优香植株能够更好地利用强光进行光合作用,积累更多的光合产物,满足生长发育的需要,但同时,较高的Rd 值表明其在黑暗条件下消耗有机物也较多,这可能是优香适应生长的机制。但下部叶暗呼吸速率随着株龄的增加呈现增加的趋势,因此,营养生长向生殖生长过渡期的Rd 最大。下部叶营养生长中期的LCP 为11.52 μmol/(m2·s),远远低于其他时期的叶片,表明此时叶片利用弱光的能力最强。从3 个生长时期的LSP 和LCP 的差值比较发现,营养生长后期对光照条件的适应范围更广,且能有效地利用弱光[18]。日光温室进行切花菊的栽培,一般都需要对其进行补光,补光的时间通常为21:00—3:00,目的是抑制花芽分化,调控花期[19],有效的利用弱光,也是对补光适应性的体现,同时也能加速植株生长。
从光响应曲线来看,除营养生长后期的上部叶外,供试3 个生长阶段、2 个叶位叶片的光饱和点介于300~600 μmol/(m2·s)。这与神马花前期和蕾后期的LSP 相近[14]。此为天气晴好时日光温室内的光照强度。如考虑到设施栽培中高密度种植的特点,植株叶片能接收到的光强远远低于这个值[20]。而在切花菊设施栽培时,为了调控花期,通常还会进行遮光处理。因此,为保证切花菊的品质和经济效益,在阳光大棚种植时,合理的确定种植密度,保证光合作用的高效性,加快植株的生长速度,避免植物徒长至关重要。
