燕麦籽粒皮裸性相关AP2/ERF 转录因子基因的克隆
2020-05-19张鸣凡陈志伟贾举庆张晓军张春来张美俊杨武德
张鸣凡,陈志伟,贾举庆,张晓军,李 欣,张春来,张美俊,杨武德
(1.山西农业大学农学院,山西太谷030801;2.山西省农业科学院作物科学研究所,作物遗传与分子改良山西省重点实验室,农业部黄土高原作物基因资源与种质创制重点实验室,山西太原030031)
燕麦属禾本科燕麦属一年生草本植物,是世界各地广泛栽培种植的一种粮、经、饲、药、草多用途作物[1],具有耐寒抗旱、耐瘠薄、耐适度盐碱、不与水稻小麦等其他禾谷类作物争夺耕地以及农业风险系数低等优良性状[2]。燕麦品种可根据外稃形态学差异分为皮燕麦与裸燕麦2 种类型[3]。皮燕麦和裸燕麦的区别主要在于成熟后籽粒外包裹的谷壳是否容易与籽粒分离,其中,皮燕麦的外稃为革质,质地坚硬,紧贴颖果;而裸燕麦的外稃为膜质,质地柔软,容易与颖果分离。与皮燕麦相比,裸燕麦籽粒成熟时的小穗外颖薄且脆,与护颖类似,手搓易与颖果分开[4]。
籽粒皮裸性即为籽粒带壳与籽粒不带壳(裸粒)的一对相对性状,研究该性状对作物育种、起源驯化具有重要作用。尽管大麦等禾本科植物都有籽粒带壳与裸粒2 种类型,但控制籽粒皮裸性的机制不完全一致[4]。在大麦中,根据图位克隆确定ERF转录因子家族基因(Nud)控制大麦籽粒皮裸性。在皮、裸大麦中扩增该目的基因发现,裸大麦的扩增片段比皮大麦的扩增片段缺失17 kb,并确认缺失的17 kb 包含了整个ERF 基因;Nud 基因与拟南芥调控脂质生物合成途径的WIN1/SHN1 转录因子基因同源[5]。
燕麦尤其是六倍体裸燕麦(Avena nuda)是我国特有的物种,是世界上重要的粮食作物之一[6]。皮燕麦较裸燕麦有一定的生长优势,尤其在抵抗外界不良环境及抗病性等方面[7]。由于二倍体燕麦较六倍体燕麦的基因组简单,因此,本研究克隆了二倍体燕麦中的大麦Nud 同源基因的AsNud,以及分析了一个二倍体皮燕麦(CIav2921)和一个二倍体裸燕麦(CIav9008)的转录组数据,旨在为寻找控制燕麦籽粒皮裸性的基因提供一定参考。
1 材料和方法
1.1 试验材料
二倍体皮燕麦(CIav2921)、二倍体裸燕麦(CIav9008)的试验材料来自美国种质资源库,这2 个材料于2019 年5 月底种植在24 cm×19 cm 的花盆里。
1.2 皮、裸燕麦外稃形态观察
在开花期取下附着于花序梗上的穗状花序进行表型分析。从成熟的小穗中取下外颖,并用镊子分解第一小花,将外颖、外稃和内稃从左至右摆好,置于体视显微镜下,选取合适的视野进行拍照观察,每个样品设3 个生物学重复,并应用cellSens Dimension 拍照软件量化开花期外稃的面积。
1.3 PCR 引物设计、扩增和测序
参照天根植物DNA 提取试剂盒说明书从试验材料的幼叶中提取基因组DNA。参照大麦Nud 基因序列,在NCBI 燕麦EST 数据库、燕麦基因组测序数据库中以及本试验的转录组数据库中比对燕麦基因组中已有的同源性最高的基因序列。利用Primer 3 在线软件设计合适的PCR 引物。参照表1所示的试验条件,在CIav2921 和CIav9008 基因组DNA 中进行目的基因扩增。扩增产物经TAE 缓冲液琼脂糖凝胶电泳、溴化乙锭染色观察后,利用康为世纪Gel Extraction kit 试剂盒对扩增产物进行纯化,然后采用TaKaRa pMDTM18-T Vector 试剂盒将纯化后的目的基因片段与T 载体连接。将反应产物转化到大肠杆菌JM109 感受态细胞,涂布于含有Amp 抗性的LB 固体平板上,对长出的单克隆进行阳性克隆验证,挑选正确的单克隆菌液送至华大公司进行测序。利用BioEdit 和DNAMAN 软件对得到的测序序列数据进行比对分析。
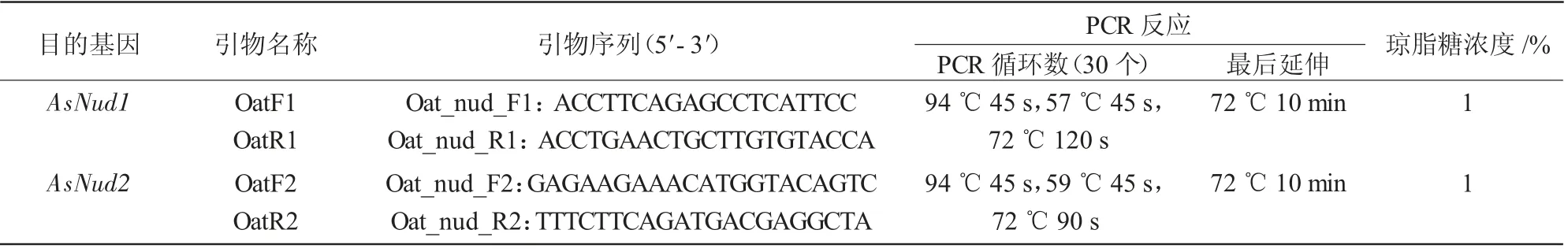
表1 PCR 引物序列和PCR 反应条件
1.4 系统发育分析
将目的基因所编码的蛋白质氨基酸序列与大麦[5]、拟南芥[8]、水稻和玉米[9]中该基因编码的同源程度较高的蛋白质氨基酸序列,在MEGA5.05[10]软件中的ClustalW 模块下进行多重序列比对,并利用Neighbor-Joining 算法(执行参数:Bootstrap method 1 000;Poissonmodel;Pairwise deletion)构建系统进化树。
1.5 RNA 提取、cDNA 文库构建及转录组测序分析
采用RNA 提取试剂盒提取CIav2921 和CIav9008 种子蜡熟期时花序的总RNA,检测合格后交由诺禾致源公司进行测序分析。cDNA 文库采用cDNA 合成试剂盒(Illumina 美国)构建,检测合格的文库利用HiSequTM2000(Illumina 美国)进行测序。RPKM值(每百万reads 中来自某一基因每千碱基长度的reads 数目)>1 且P 值<0.05 的基因被认为是可表达的基因。每个样品设3 个生物学重复。
1.6 数据分析
采用SPSS 和Microsoft office 2010 软件进行数据处理及方差分析,采用Origin 软件作图。
2 结果与分析
2.1 二倍体皮、裸燕麦外稃的形态学差异分析
开花期时的CIav2921 与CIav9008 的小穗相比,颖片(Glume)的形态学没有差异,而外稃(Lemma)却有明显差异,其中,裸燕麦的外稃与颖片类似,能看到清晰的叶脉,表面无光泽;皮燕麦的外稃只在近上部1/3 处能看到叶脉,表面通体泛有光泽(图1-A、C)。成熟期皮、裸燕麦的外颖和内稃形态没有差异,而外稃有差异,其中,裸燕麦的外稃舒展;皮燕麦的外稃弯曲,与籽粒紧密相连(图1-B、D)。比较皮、裸燕麦外稃面积发现,裸燕麦的面积显著大于皮燕麦的面积(图1-E)。
2.2 AsNud 基因的分离与结构分析
将大麦Nud 基因序列放入NCBI 燕麦EST 数据库、燕麦基因组测序数据库以及本试验的转录组数据库中进行比对,结果在已有的燕麦基因组中找到同源性较高的4 条基因序列(GO593063.1、BMK_Unigene_060579、lcl|CTG24048、lcl|CTG76330)。其中GO593063.1 序列不完整,为一片段;将序列BMK_Unigene_060579 和GO593063.1 利用BioEdit 软件比对后发现,二者在共有的碱基处相似性极高,遂将BMK_Unigene_060579 之后的序列拼接到序列GO593063.1 之后,成为一条拼接序列。利用Primer 3在线软件设计出的引物扩增这5 条目的基因,经切胶回收、T 载体连接、感受态细胞转化、阳性克隆验证、菌液测序后,将得到的序列利用BioEdit 软件进行比对,结果认为,序列BMK_Unigene_060579、序列GO593063.1 和拼接序列为同一条序列,记作AsNud1;序列lcl|CTG24048 和序列lcl|CTG76330 为同一条序列,记作AsNud2。
通过对AsNud 基因序列特征进行分析,研究AsNud 基因序列的遗传多样性,结果显示(表2),CIav2921AsNud1 基因组的DNA 长度为744 bp、CIav9008AsNud1 基因组为744 bp、HvNud 为884 bp;CIav2921AsNud1 开放阅读框(ORF)长度为48 bp、CIav9008AsNud1 开放阅读框为639 bp、HvNud 为684 bp;CIav2921AsNud1 预测的基因产物包含15 个氨基酸残基,CIav9008AsNud1 为212 个氨基酸、HvNud 为227 个氨基酸;CIav2921AsNud1 估计分子 质 量 为1.9 ku、CIav9008AsNud1 为22.8 ku、HvNud 为24.1 ku;CIav2921AsNud1 等电点为12.83、CIav9008AsNud1 为10.03、HvNud 为9.74。CIav2921 AsNud1 ORF 的GC 含量(54.17%)低于大麦HvNud ORF(65.64%)及全基因组DNA(61.20%),CIav9008 AsNud1 ORF 的GC 含量(61.19%)低于大麦HvNud ORF(65.64%) 及大麦基因组DNA(61.20%)。CIav2921AsNud1 与大麦HvNud 在基因组DNA 中核苷酸相似性为65.64%,而在ORF 与肽水平上,相似性分别为6.14%和6.17%;CIav9008AsNud1 与大麦HvNud 在基因组DNA、ORF 与肽水平上,其相似性分别为71.37%、73.68%和69.60%。
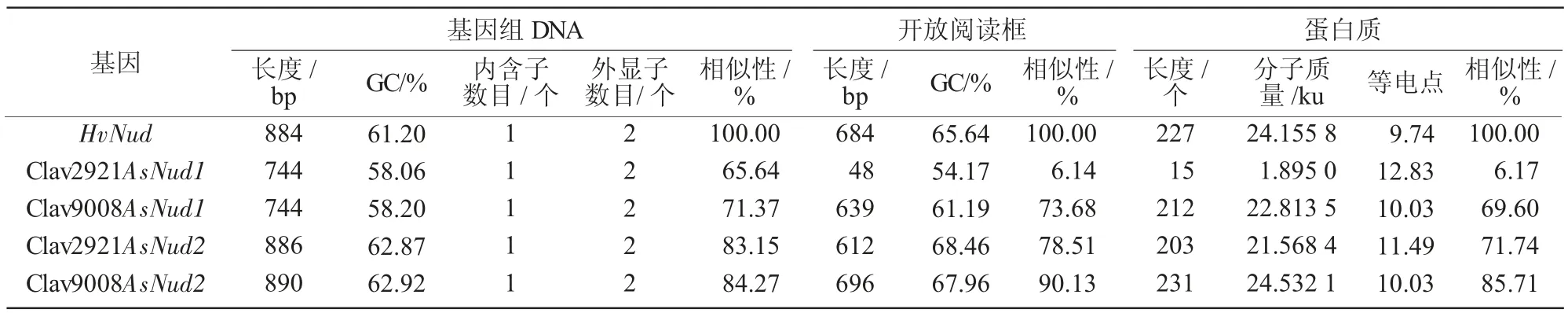
表2 HvNud 与燕麦同源基因AsNud 的同源性
CIav2921AsNud2 基因组DNA 长度为886 bp、CIav9008AsNud2 基因组为890 bp、HvNud 为884 bp,CIav2921AsNud2 开放阅读框(ORF)长度为612 bp、CIav9008AsNud2 开放阅读框为696 bp、HvNud 为684bp;CIav2921AsNud2 预测的基因产物包含203 个氨基酸残基、CIav9008AsNud2 为231 个氨基酸、HvNud 为227 个氨基酸;CIav2921AsNud2 估计分子质量为21.5 ku、CIav9008AsNud2 为24.5 ku、HvNud为 24.1 ku;CIav2921AsNud2 等 电 点 为 11.49、CIav9008AsNud2 为10.03、HvNud 为9.74。CIav2921 AsNud2 ORF 的GC 含量(68.46%)高于大麦HvNud ORF(65.64%)及全基因组DNA(61.20%);CIav9008 AsNud1 ORF 的GC 含量(67.96%)高于大麦HvNud ORF (65.64%) 及 全 基 因 组DNA (61.20%)。CIav2921AsNud2 与大麦HvNud 在基因组DNA 中核苷酸相似性为83.15%,而在ORF 与肽水平上,相似性分别为78.51%和71.74%;CIav9008AsNud2 与大麦HvNud 在基因组DNA、ORF 与肽水平上,其相似性分别为84.27%、90.13%和85.71%。
对 预 测 的CIav2921、CIav9008AsNud 和 大 麦HvNud 编码的氨基酸序列分析发现,AsNud1 与AsNud2 基因同HvNud 一样,都具有一些保守的序列特征,包括AP2/ERF 结构域、保守的中间基序(mm)和保守的C 末端基序(cm);除了这几个保守结构域外,AsNud1、AsNud2 与HvNud 又具有程度不一的差异。对比AsNud1 和AsNud2 基因在CIav2921 和CIav9008 中的差异发现,AsNud1 在二者中差异不大,而AsNud2 在二者中差异较大,特别是在C 末端。值得一提的是,CIav2921AsNud1 和CIav2921AsNud2 编码的氨基酸有提前终止现象(AsNud1 基因在第17 位氨基酸处,AsNud2 基因在第204 位氨基酸处),而CIav9008 中则没有此现象。(图2-A、B)。
2.3 AsNud 同源基因的系统发育分析
大麦HvNud 基因与拟南芥WIN1/SHN1 转录因子基因同源,其中,拟南芥有3 个(AtWIN1/SHN1、AtSHN2 和AtSHN3)同源程度很高且参与蜡质和角质合成调控过程的基因,均属于AP2/ERF 转录因子家族。将该转录因子下的6 个拟南芥基因(At1g15360、At5g11190、At5g25390、At5g25190、At5g67010 和At-2g20350)、同源程度很高的7 个水稻基因(Os02g1-0760、Os06g40150、Os06g08340、Os02g55380、Os04g-56150、Os02g32140 和Os04g32620)、3 个玉米基因(Zm2g085678、Zm2g111415 和Zm2g104260)、1 个西红柿基因(LeERF1)以及根据大麦HvNud 基因同源克隆得到的2 个二倍体皮、裸燕麦基因(AsNud1和AsNud2)编码的氨基酸序列在MEGA 5.05 软件上经ClustalW 多序列比对,采用邻接法(Neighborjoining,NJ)构建系统进化树(图3)。
从图3 可以看出,AtWIN1/SHN1(At1g15360)、AtSHN2(At5g11190)和AtSHN3(At5g25390)在进化树上属于同一个分支,其中,AtWIN1/SHN1 与水稻中的Os02ERF(Os02g10760)和Os06ERF(Os06g40150)、与皮、裸燕麦中的AsNud1 和AsNud2、与玉米中的Zm2g085678 和大麦HvNud 的同源程度比较高。另外,玉米的Zm2g085678 与Os02ERF 和AsNud1 同源程度很高,同属于一个小进化分支。Os06ERF 与HvNud 和AsNud2 同源程度很高,同属于一个小进化分支,在该分支中,AsNud2(CIav2921)与大麦HvNud 的同源性更高。
2.4 AsNud 基因的转录分析
将CIav2921 和CIav9008 种子蜡熟期时花序进行转录组测序,基于RNA-Seq 数据,分析了AsNud1与AsNud2 在2 个材料中的表达情况,结果显示(图4),AsNud1 在2 个材料中都有表达,在CIav2921中的表达量(RPKM 值0.76)低于在CIav9008 中的表达量(RPKM 值1.34),但差异未达显著水平。AsNud2 在2 个材料中也都有表达,在CIav2921 中的表达量(RPKM 值4.46)显著高于在CIav9008 中的表达量(RPKM值0.65)。
3 结论与讨论
AP2/ERF 转录因子是一个数目庞大的家族,因含有高度保守的AP2/ERF DNA 结合域而得名[11]。AP2/ERF 转录因子至少含有一个名为AP2 结构域的DNA 结合结构域,是由约60 个氨基酸残基按照3 个β 折叠和1 个α 螺旋的方式所构成的一个典型的三维结构[12]。本研究利用大麦中的AP2/ERF 转录因子家族中的HvNud 基因为种子序列,采用电子克隆的方式,从二倍体皮、裸燕麦中克隆到2 条同源序列,序列分析发现,AsNud1、AsNud2 和HvNud 一样,都存在由大约68 个保守残基组成的被称为AP2 结构域的DNA 结合基序[13],因此,本试验克隆到的AsNud1 和AsNud2 都属于AP2/ERF 转录因子家族。此外,AsNud1 和AsNud2 还存在另外2 个保守的中间基序(mm)、C 末端基序(cm),这2个保守基序也同样存在于HvNud 基因中[5]。结合转录组的数据,对基因的结构分析发现,AsNud 基因与大麦的HvNud 基因具有相同的基因结构,都是由2 个外显子和1 个内含子组成。系统发育分析发现,AsNud 基因与大麦的HvNud 基因、拟南芥的WIN1/SHN1、水稻的Os02ERF 和Os06ERF、玉米的Zm2g085678 亲缘关系最近。相比AsNud1,AsNud2与HvNud 的关系更近。对AsNud1 和AsNud2 序列比对发现,二者的相似性达到77%,因此,二者为旁系同源基因。
AP2/ERF 转录因子是植物中广泛存在且特有的一类转录因子,参与调控植物生长发育及抗逆反应等生命过程。第一个AP2/ERF 转录因子基因在模式植物拟南芥中被分离出来,且被证实参与了花发育过程的调控[13]。其他植物中的AP2/ERF 转录因子也有陆续报道。本研究中克隆到的AsNud 基因,其同源基因在水稻中影响水稻的木质素含量,是转录抑制子,该基因可能参与植物次生细胞壁的生物学过程[9],这与皮燕麦外稃坚硬且木质素含量高、裸燕麦外稃柔软且木质素含量低对应一致;该基因还与拟南芥SHN/WIN 转录因子家族基因同源,拟南芥SHN/WIN 转录因子家族与蜡质和角质合成调控有关[8],这与皮燕麦外稃平滑有光泽、裸燕麦外稃不平滑无光泽对应一致;该基因在大麦中控制大麦的皮裸性[5]。燕麦籽粒的皮裸性与大麦有相似之处又有所不同。成熟后的皮燕麦外稃质地坚硬,与籽粒紧密相连,不易与籽粒分离,而裸燕麦外稃质地柔软,容易与籽粒分离,这点与大麦皮裸性相似,即裸大麦外稃较皮大麦外稃更易与籽粒分离。大麦开花10 d 后,皮大麦的颖果表面会出现一种黏性物质,将颖果与外稃黏附在一起[14]。大麦皮裸性由位于染色体7HL 上的单个基因位点(nud)控制[15],裸大麦与皮大麦相比,缺失固定的17 kb ERF 基因。而在燕麦中,皮、裸燕麦虽然不存在这种缺失,但皮燕麦(CIav2921)较裸燕麦(CIav9008)会提前终止编码。本研究表明,这种差别可能与籽粒的皮裸性相关,同时可以推测大麦与燕麦中控制皮裸性的机制不完全一致。对AsNud 基因的表达研究发现,AsNud1和AsNud2 在CIav2921(皮)和CIav9008(裸)的花器官中都有表达,但表达模式不同,其中,AsNud1在CIav2921 中的表达量低于在CIav9008 中的表达量,但差异未达显著水平;而AsNud2 在CIav2921中的表达量显著高于在CIav9008 中的表达量。这种旁系同源基因表达模式的差异在多种植物中都有报道[16-19]。此外,AsNud2 在CIav2921 和CIav9008 燕麦中的表达有明显的差异,说明AsNud2 基因的调控区或上游的调控因子在该皮、裸燕麦中出现了差异,这可能是导致这2 个材料籽粒皮裸性不同的另一个原因。
