雨生红球藻HaeDGAT2-3 的分子克隆和生物信息学分析
2020-05-19张宏江赵春超许文鑫崔红利李润植
张宏江,赵春超,许文鑫,杭 伟,崔红利,李润植
(山西农业大学分子农业与生物能源研究所,山西太谷030801)
化石能源的过度开采以及煤炭、石油等的大量使用是造成化石能源枯竭、全球气候变暖的主要原因。温室效应的“罪魁祸首”CO2是植物光合作用所必需的底物材料,光合作用过程中,绿色植物可以在光能作用下,将所吸收的CO2和空气中的水分转化成碳水化合物并且释放出O2。微藻是一种诞生于亿万年前并且普遍存在于地球上的光合自养绿色植物[1],它可以利用光合作用高效吸收CO2,并将其转化为对人类有益的高附加值活性成分,如高蛋白、多糖、虾青素和生物柴油等[2-4]。近年来,微藻由于其具有光合效率高、繁殖速度快、生长周期短等优点,被认为是解决能源危机和治理环境污染问题的最佳选择。
三酰甘油(TAGs)是真核生物能量储存的主要形式,同时也是生物柴油原材料的理想来源[5]。植物体内的TAGs 合成主要有2 种途径:一种是乙酰辅酶A(acyl-CoA)依赖型,也称肯尼迪途径;另一种是不依赖于乙酰辅酶A 的代谢途径[6]。其中,肯尼迪途径中TAGs 的合成主要是在3 种酰基转移酶的作用下将底物acyl-CoA 顺序酰基化到甘油分子(Glycerol-3-phosphate)sn-1、sn-2、sn-3 位点的过程[7]。这3 种酰基转移酶依次为甘油三磷酸酰基转移酶(Glycerol-3-phosphateacyltransferase,GPAT)、溶血磷脂酸酰基转移酶(Lyso-phosphatidic acidacyltransferase,LPAT)和二酰甘油酰基转移酶(Diacylglycerol acyltransferase,DGAT)[8]。其中,DGAT 将酰基CoA 上的酰基部分转移到二酰基甘油(DAG)的sn-3 位置,从而完成TAGs 的最终合成,是TAGs 合成过程中的唯一关键限速酶[9-10]。目前,根据DGAT功能域、亚细胞定位等差异将其分为4 种类型:DGAT1、DGAT2、DGAT3 和WS/DGAT[11-12]。其中,DGAT1 和DGAT2 是结合在内质网上并编码内嵌于膜酯双分子层的微粒体酶,二者具有不同的拓扑结构;而DGAG3 和WS/DGAT 是最近刚发现的具有DGAT 活性的酶,DGAT3 是一种可溶性酶,WS/DGAT 除了具有DGAT 活性外还具有催化腊酯(Wax ester)合成的活性[12-13]。
雨生红球藻(Haematococcus pluvialis)是一种单细胞真核绿藻,在胁迫条件下(如高温、高光和N胁迫等),雨生红球藻细胞形态由快速生长的绿色营养细胞转变为红色厚壁孢子状态[14-15],此时虾青素在藻细胞内大量积累以抵御不良环境。天然虾青素具有极强的抗氧化性,抗氧化能力是维生素C 的65 倍,是β- 胡萝卜素的54 倍[16]。相较于其他产虾青素来源(红法夫酵母、南极磷虾等),雨生红球藻具有生长速度快、虾青素产量高(达细胞干质量的4%)等优点,是公认的天然虾青素的理想来源[17];此外,其还可以有效清除氧自由基[17],具有延缓衰老的功效,目前已被广泛应用于化妆品、食品等行业。
雨生红球藻中虾青素主要以虾青素单酯和虾青素双酯的形式存在,少量以游离形式存在。有研究表明,雨生红球藻受到外界胁迫时,油脂含量会随之增加[18]。超微结构显示,雨生红球藻产生的虾青素酯在虾青素积累期储存于富含TAGs 的油体中[18]。有研究表明[19],DGAT 可能是催化虾青素酯化的关键酶。此外,莱茵衣藻、三角褐指藻和小球藻来源的DGAT 都具有恢复DGAT 缺陷型酵母H1246菌株合成三酰甘油能力的活性[20-21]。然而,雨生红球藻中的DGAT 目前尚未见报道。
本研究以同源克隆的方法获得雨生红球藻中DGAT 基因,并利用生物信息学手段对其理化性质、高级结构和系统进化关系进行分析,以期为雨生红球藻虾青素酯化问题研究提供理论依据。
1 材料和方法
1.1 试验材料
1.1.1 试验藻种及培养条件 试验所用藻种为雨生红球藻Flotow 1844(购自Dunstaffnage Marine Laboratory),现保存于山西农业大学分子农业与生物能源研究所。所用藻种培养基为BG11 培养基,藻种培养于人工气候培养箱,培养条件设置为温度(23±1)℃、光/ 暗周期12 h/12 h、白天光照强度1 400 lx,并且每8 h 摇晃一次。
1.1.2 试验设备 研究所用仪器为:高压蒸汽灭菌锅Panasonic MLS-3781L(日本松下电器产业株氏会社);超净工作台Boxun BJ-CD(上海博迅实业有限公司);人工气候培养箱BIC-400(上海博讯实业有限公司);电泳仪JY-SPCT(北京君意东方电泳设备有限公司);凝胶成像仪、PCR 仪BIO-RAD(伯乐生命医学产品(上海)有限公司);分光光度计Nano Drop 2000(美国赛默飞世尔科技公司)。
1.2 试验方法
1.2.1 总RNA 的提取及cDNA 的获得 雨生红球藻总RNA 通过TRizol 法提取[22],并根据所提取的雨生红球藻RNA 的浓度利用反转录试剂盒(Prime-ScriptTMRT)获得cDNA 模板。为避免实验过程中样品污染,操作人员应佩戴口罩及头套,所用离心管均为提取RNA 专用离心管。另外,将所提取的RNA保存于-80 ℃冰箱。
1.2.2 同源克隆及RACE 扩增 对来自莱茵衣藻CC3491、小球藻ATCC 30412 以及三角褐指藻的DGAT2 进行序列比对,从而获得高度保守序列;在此基础上,利用CODEHOP 软件进行设计雨生红球藻DGAT2 的2 段同源克隆引物(F1、R1 和F2、R2)。以获得的雨生红球藻cDNA 为模板,按照TaKaRa LA TaqR扩增体系进行PCR 扩增,从而获得雨生红球藻HaeDGAT2-3 的同源克隆片段。RACE cDNA 模板的制备按照试剂盒(SMARTerTMRACE cDNA Amplication Kit)说明书进行。在同源克隆片段的基础上获得5′端和3′端引物(5′RACE R3、R4 和3′RACE F3、F4)。以RACE cDNA 为PCR扩增模板,以5′端和3′端引物为PCR 扩增引物获得目的基因两端(5′端和3′端)的片段。通过DNAStar SeqMan 软件对中间以及5′端和3′端RACE 片段进行拼接获得雨生红球藻HaeDGAT 基因cDNA 序列全长,并据此设计表达框引物(F5、R5)。所用引物信息如表1 所示,均由上海生工生物工程有限公司合成。

表1 引物信息
1.3 生物信息学分析
雨生红球藻HaeDGAT2-3 氨基酸序列是通过序列处理在线工具包(SMS)(http://www.bio-soft.net/sms/),将获得的HaeDGAT2-3 的核苷酸翻译而来,开放阅读框的查找是通过在线ORFfinder 软件(https://www.ncbi.nlm.nih.gov/orffinder/)对目的基因进行;通过在线软件ProtParam(http ://web.expasy.org/protparam/)对雨生红球藻HaeDGAT2-3 的理化性质进行分析;跨膜域及疏水区的预测是通过在线软件TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)及ProtScale(http ://web.expasy.org/protscale/)进行;雨生红球藻HaeDGAT2-3 及其他物种来源的DGAT2 蛋白保守功能域通过NCBI 的CCD 在线软件(https://www.ncbi.nlm.nih.gov/cdd/)进行预测;二级结构及三级结构的预测分别通过SOPMA(http://metadatabase.org/wiki/SOPMA) 和 SWISS-MODEL(http://swissmodel.expasy.org/)进行;多序列比对是通过本地软件BioEdit 进行分析;系统进化树是通过Clustal W 软件进行序列比对,再通过本地软件MEGA 7.0 进行绘制,最终通过在线软件EvolView(https://www.evolgenius.info/)进行修饰。
2 结果与分析
2.1 总RNA 的提取和基因克隆结果分析
由图1 可知,泳道1 从上往下依次是28SrRNA、18S rRNA 和5S rRNA 等3 个条带,其中,28S rRNA、18S rRNA 条带带型清晰,5S rRNA 条带较模糊,说明所提RNA 质量较好;NanoDorp 2000 测定总RNA浓度是850 ng/μL,A260/A280及A260/A230值分别为1.8~2.0 和1.9~2.0,说明所提RNA 质量较好,可进行后续试验;泳道2 和3 大小分别为482、773 bp;泳道4和5 大小分别为667、1 149 bp。
2.2 雨生红球藻HaeDGAT2-3 蛋白cDNA 序列分析
雨生红球藻2 型二酰甘油酰基转移酶DGAT2(HaeDGAT2-3)的cDNA 序列全长是在同源克隆的基础上利用RACE (rapid-amplification of cDNA ends)技术获得,NCBI 注册号为MN073496。如图2所示,HaeDGAT2-3 序列全长为1 446 bp,编码区从248~1 396 bp 共1 149 bp,编码383 个氨基酸,其中,5′端非编码区(5′-UTR)和3′端非编码区(3′-UTR) 长度分别为247、49 bp。利用NCBI 中的BLASTp 程序对HaeDGAT2-3 氨基酸序列进行比对分析,结果发现,HaeDGAT2-3 基因与衣藻来源的DGAT2 基因(NCBI 检索号为GAX79066.1)相似性最大(54.02%),与小球藻、月牙藻来源的DGAT2 基因(NCBI 检索号分别为QBG05555.1、GBF91693.1)相似性分别达45.08%和41.82%。
2.3 雨生红球藻HaeDGAT2-3 蛋白理化性质分析
雨生红球藻HaeDGAT2-3 蛋白基本理化性质通过在线软件ExPASY 中的ProtParam 工具进行分析,结果显示,该蛋白分子式为C1844H2824N486O489S17,分子质量为40.17 ku,理论等电点为9.36,属于碱性蛋白。编码HaeDGAT2-3 蛋白的20 种氨基酸中丙氨酸(Ala)含量最高(13.3%);其次为亮氨酸(Leu)和丝氨酸(Ser),含量分别为10.6%和7.6%;而色氨酸(Trp)和半胱氨酸(Cys)含量偏低,均为1.6%。说明该蛋白属于不稳定蛋白。在线分析软件ProtScale 分析结果显示,HaeDGAT2-3 蛋白存在至少3 个疏水区,属于疏水蛋白。
2.4 雨生红球藻HaeDGAT2-3 蛋白的高级结构分析
雨生红球藻HaeDGAT2-3 蛋白的二级结构通过在线软件SOPMA 进行分析,结果显示(图3-A),目的蛋白中无规则卷曲所占比例最高(44.87%),其次为α- 螺旋(32.88%),延伸链和β- 折叠分别占18.75%和5.71%。图3-B 为雨生红球藻DGAT2 的三级结构预测结果,图3-C 为三级结构所参照模型。
雨生红球藻HaeDGAT2-3 与其他不同来源DGAT2 蛋白的保守域通过NCBI 的在线工具进行预测,结果显示(图4-A),同其他来源的DGAT2 相同,HaeDGAT2-3 基因也有DGAT 基因的典型LPLAT 家族,暗示HaeDGAT2-3 具有与其他来源DGAT2 蛋白相似的功能;利用在线分析软件TMHMMServer 对雨生红球藻HaeDGAT2-3 与其他不同来源DGAT2 蛋白的跨膜域进行预测,结果显示(图4-B),HaeDGAT2-3 蛋白在40—90 位氨基酸区间内有2 个明显的跨膜域;对其他来源DGAT2 蛋白的跨膜域进行分析表明,DGAT2 蛋白有1~3 个跨膜域。此外,信号肽分析结果显示, 雨生红球藻HaeDGAT2-3 中无信号肽存在。图4 中DGAT2 序列相关信息如表2 所示。
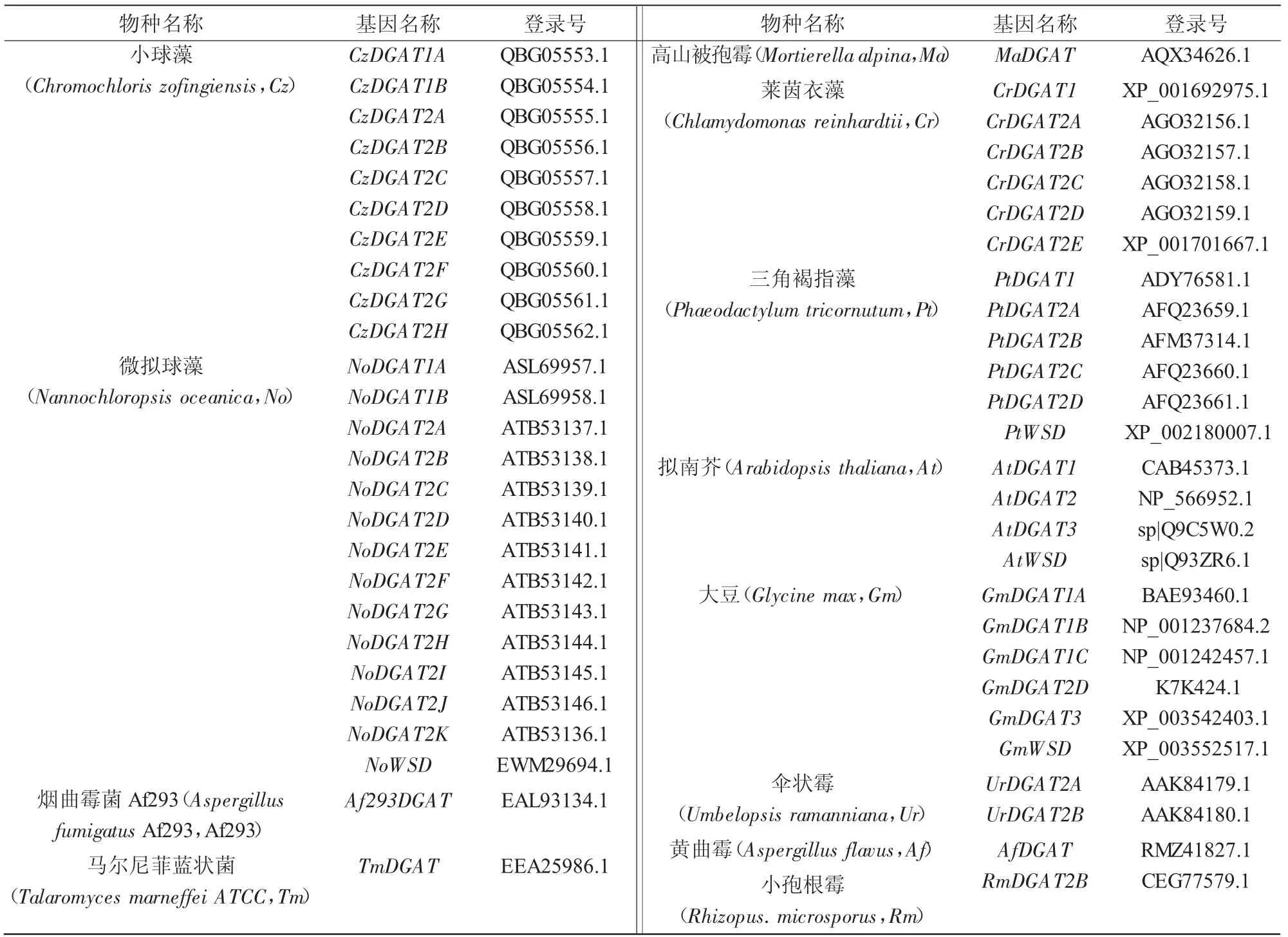
表2 DGAT 基因名称及其登录号
2.5 雨生红球藻HaeDGAT2-3 多序列比对分析
通过本地软件BioEdit 对HaeDGAT2-3 氨基酸序列和其他来源的DGAT2 进行比对分析,结果显示(图5),HaeDGAT2-3 氨基酸序列中共包含7 个保守的功能基序,分别为YFP、PH、PR、GGE、RGFA、VPFG 和G 基序。在YFP 基序中,所有序列前2 个氨基酸皆为YF,而只有HaeDGAT2-3、AtDGAT2、CzDGAT2D 等7 个序列在第3 个氨基酸位置上为苯丙氨酸(Phe);在PH 基序中,只有CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 等4 个序列第3 个氨基酸位置上的甘氨酸(Gly,G)被丝氨酸(Ser,S)所取代,暗示这4 个氨基酸序列在功能上相似;在PR 基序中,雨生红球藻来源的HaeDGAT2-3 氨基酸序列同高等植物来源的DGAT(AtDGAT2 和GmDGAT2)在第1 个位置上的氨基酸保持一致,第2 个位置上的氨基酸在所测试序列中散乱分布,只有氨基酸序列相对特殊的微拟球藻来源的NoDGAT2K 在第4 个位置上的氨基酸是甘氨酸(Gly,G),其余全都是精氨酸(Arg,R);在GGE 基序中,所有序列都含有完整的GGE 基序,但是在2 个位置上的氨基酸并不保守,而只有CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 这4 个氨基酸序列在这2 个位置上的氨基酸保持一致,再一次暗示其功能相似;在基序RGFA 中,雨生红球藻HaeDGAT2-3 异亮氨酸(Ile,I)替换了苯丙氨酸(Phe,F),这可能是造成功能差异的关键原因;VPFG 基序相比其他基序有很大的差异性,相对不保守;在基序G 中,所有氨基酸序列都有完整的G 基序,相对保守。从图5 的分析结果可以看出,HaeDGAT2- 3 与CrDGAT2A、CzDGAT2A和CzDGAT2B可能具有相同的功能。
2.6 雨生红球藻HaeDGAT2-3 的系统进化分析
为进一步研究雨生红球藻来源的HaeDGAT2-3与其他来源DGAT 的进化关系,从NCBI 数据库以及已发表的文献中搜集到53 条DGAT 序列,包括微藻、菌类和高等植物来源的DGAT1s、DGAT2s、DGAT3s 和WS/DGATs 等4 种不同类型的二酰甘油酰基转移酶,在序列比对的基础上对其进行系统进化分析,结果显示(图6),4 种不同类型的DGATs分别聚为一支,而且微藻、菌类和高等植物来源的DGAT 在各自的类群中明显分开;值得注意的是,HaeDGAT2-3 与CrDGAT2A、CzDGAT2A和CzDGAT 2B聚为一个小支,暗示它们具有相同的功能,进一步验证了多序列比对的预测结果。图6 进化树所有序列的相关信息列于表2。
3 结论与讨论
雨生红球藻是一种单细胞绿色微藻,被认为是天然虾青素的理想来源[5],并且其产生的虾青素以虾青素单酯和虾青素双酯的形式存储于富三酰甘油(TAGs)的油体中。二酰甘油酰基转移酶(DGAT)是依赖于乙酰辅酶A(acyl-CoA)合成TAG 的肯尼迪途径中催化TAG 生物合成最后一步的关键限速酶[9-10],目前,已有大量来源于不同物种(包括高等植物、微藻和真菌)的DGAT 被用来探究DGAT 的表达与脂肪酸含量及成分的变化关系,研究发现,高等植物中续随子来源的EiDGAT2 基因可以提高烟草叶片中的总脂含量[23],紫苏来源的PfDGAT1 被发现可以恢复TAG 缺陷型酵母H1246 重新合成油脂的能力[24];高山被包霉来源的MaDGAT 基因以及毛霉菌来源的McDGAT 基因被证实具有恢复H1246合成油脂的能力,但它们有着不同的底物偏好性[25-26];此外,莱茵衣藻、微拟球藻、三角褐指藻等微藻来源的DGAT 也相继被发现具有催化TAG 合成的能力[5,21,27]。然而,雨生红球藻来源的DGAT2 并未见报道。
本研究通过同源克隆方法获得了雨生红球藻DGAT2 的同源片段及同源克隆引物,并在此基础上通过RACE 扩增技术获得雨生红球藻HaeDGAT2-3的cDNA 序列全长1 446 bp,编码383 个氨基酸,其中,丙氨酸(Ala)含量最高,为13.3%。此外,雨生红球藻HaeDGAT2-3 蛋白分子质量为40.17 ku,理论等电点为9.36。二级结构分析结果显示,无规则卷曲所占比例最高,为44.87%。根据NCBI中的BLASTp分析结果显示,雨生红球藻来源的DGAT2 与莱茵衣藻、小球藻和月牙藻来源的DGAT2 相似性均在40%以上,分别达54.02%、45.08%和41.82%,说明其功能具有相似性。DGAT1s 和DGAT2s 虽然在分子结构上有很大的差异,属于2 个不同的基因家族,但在功能水平上却是一致的。DGAT1s 一般具有8~9 个跨膜域,而DGAT2s 中跨膜域相对较少,一般为1~3 个。比如,紫苏、小球藻来源的DGAT1 都有9 个跨膜螺旋区[27-28],续随子来源的DGAT2 有2 个跨膜区[24]。本研究发现,HaeDGAT2-3 蛋白具有2 个跨膜区,符合DGAT2 家族序列特点。保守域分析结果显示,雨生红球藻来源的DGAT2 具有同其他DGAT2来源相同的LPLAT 家族。
此外,本研究对HaeDGAT2-3 与微藻、高等植物和细菌来源的DGAT 进行多序列比对,结果表明,同其他物种来源的DGAT2 相同,HaeDGAT2-3也具有7 个功能域;在这7 个功能域中,YFP、RGFA和VPFG 基序的保守性较低。同时,CzDGAT2D、CrDGAT2D、AtDGAT2 和GmDGAT2 这4 个氨基酸序列具有很高的相似性。值得注意的是,在第5 个基序RGFA 中,HaeDGAT2-3 蛋白中异亮氨酸(Ile,I)替换了苯丙氨酸(Phe,F),暗示HaeDGAT2-3 基因可能具有不同的功能。系统进化分析结果表明,HaeDGAT2-3 基因确实属于DGAT2 家族,并且与CrDGAT2A、CzDGAT2A 和CzDGAT2B 聚为同一个小支,暗示其具有相同的功能。
本研究在同源克隆的基础上利用RACE 扩增技术首次获得了雨生红球藻中2 型二酰甘油酰基转移酶DGAT2 的cDNA 序列全长,并利用生物信息学方法对其进行分析,结果显示,HaeDGAT2-3 蛋白分子式为C1844H2824N486O489S17,分子质量为40.17 ku,编码区全长1 149 bp,共编码383 个氨基酸。功能域分析表明,其可能具有催化TAG 合成的功能。同其他来源的DGAT2 相同,HaeDGAT2-3 也具有7 个保守的功能域,而在RGFA 基序中,HaeDGAT2-3表现出一定的特异性,暗示其功能可能不尽相同。进化树分析结果表明,其与已经报道的具有TAG合成功能的DGAT2 相似性极高,暗示其可能亦具有合成TAG 的功能。本试验结果可为进一步了解雨生红球藻中虾青素酯化机制提供参考。
